綠色熒光蛋白基因在木葡糖酸醋桿菌中的表達
2015-01-08 09:52:40韓培培譚之磊賈士儒
天津科技大學學報 2015年4期
關鍵詞:綠色
鐘 成,劉 淼,李 晶,韓培培,譚之磊,賈士儒
(工業發酵微生物教育部重點實驗室,天津科技大學生物工程學院,天津 300457)
木葡糖酸醋桿菌(Gluconacetobacter xylinus)的靜置培養過程中,在氣液界面處生成細菌纖維素膜,這一現象被認為是木葡糖酸醋桿菌的趨氧性造成的[1].此外,李晶等[2]研究表明,木葡糖酸醋桿菌對氨基酸、碳源、酸和重金屬等都有一定程度的趨化性.Sano 等[3]通過在水中外加電場,利用電解的作用產生氧氣,誘導好氧性的木醋桿菌有序運動,從而實現了細菌纖維素的有序纖維結構.然而,目前研究者對微生物的趨化過程和鞭毛驅動蛋白的作用機理尚不明確.
綠色熒光蛋白(green fluorescent protein,GFP)是由238 個氨基酸組成的單體蛋白,其相對分子質量約為2,688.GFP 最初是由Shimomure[4]從多管水母屬(Aequorea victoria)中分離出來的,以其良好的物理特性和熒光特性,而成為良好的報告基因和熒光標記分子,并在探索生命現象過程中得到了非常廣泛的應用[5],可用于活細胞中直接觀察細胞運動.本文通過構建重組質粒pMV24-gfp+,并在木葡糖酸醋桿菌中進行表達,證明其可作為選擇標記用于今后的研究.開展GFP 相關的信號傳導研究,可為研究木葡糖酸醋桿菌生物合成細菌纖維素與趨化性之間的關系提供重要的依據.
1 材料與方法
1.1 材料
1.1.1 菌種、質粒和培養基
木葡糖酸醋桿菌(Gluconacetobacter xylinus)CGMCC 2955 由工業發酵微生物教育部重點實驗室篩選,中國普通微生物菌種保藏管理中心(China general microbiological culture collection center,CGMCC)保藏.大腸桿菌(E.,coli)DH5α 由工業發酵微生物教育部重點實驗室保藏.
pMV24 穿梭載體由日本Mitsukan 集團有限公司贈予.
木葡糖酸醋桿菌培養基(g/L):葡萄糖25,酵母粉7.5,蛋白胨10,Na2HPO410,初始pH 6.0.固體培養基加2%瓊脂.121,℃滅菌15,min.
LB 培養基(g/L):蛋白胨10,酵母粉5,氯化鈉10,pH 7.0.固體培養基加 2%瓊脂.121,℃滅菌15,min.
1.1.2 試劑與儀器
蛋白酶K,北京鼎國生物技術有限責任公司;溶菌酶、引物、Taq DNA 聚合酶、dNTPs,上海生工生物工程有限公司;DNA Marker、pCR2.1,T-Vector、DNA柱回收試劑盒、DNA 凝膠回收試劑盒,大連寶生物科技(北京)有限公司;RNase A,美國Genview 公司;限制性內切酶,Fermentas 公司.
電熱恒溫水浴鍋,天津市中環實驗電爐有限公司;臺式高速離心機,上海醫用分析儀器廠;穩壓穩流電泳儀,北京六一儀器廠;基因擴增儀、凝膠分析儀、Micro pulser 型電轉化儀,美國Bio-Rad 公司;熒光顯微鏡、BX51 型熒光顯微鏡,日本Olympus 公司.
1.2 方法
1.2.1 引物設計
設計引物分別在上游和下游加上 EcoRⅠ和XbaⅠ的酶切位點及相應保護堿基.F:5′-CGGAATT CAAGAAGATATACATATGGCT-3′;R:5′-CTCTAGA CTCGAATTCATTATTTGTAG-3′.下劃線為酶切位點.
1.2.2 轉基因木葡糖酸醋桿菌CGMCC,2955/pMV24-gfp+的構建
以質粒pMUTIN-gfp+為模板擴增gfp 基因.PCR體系為25,μL 反應程序:95,℃模板預變性5,min;95,℃模板變性30,s,50,℃引物退火30,s,72,℃引物延伸1,min,共33 個循環;最后72,℃延伸10,min.PCR 產物利用1%瓊脂糖凝膠電泳進行分析.
取出后純化PCR 產物.PCR 產物片段與表達載體pMV24 用EcoRⅠ和XbaⅠ雙酶切.酶切后的gfp基因與pMV24 按3∶1 和5∶1 體系連接,得到重組質粒pMV24-gfp+.質粒構建流程如圖1 所示.將重組質粒化轉入E.,coli DH5α (方法見1.2.3),陽性克隆進行質粒提取并測序.將同源性最高(100%)的重組質粒命名為pMV24-gfp+.
提取pMV24-gfp+質粒,電轉入木葡糖酸醋桿菌CGMCC 2955 感受態細胞(方法見1.2.4).隨機挑取轉化子,提取質粒,酶切驗證并測序.驗證正確的轉化子命名為CGMCC 2955/pMV24-gfp+.
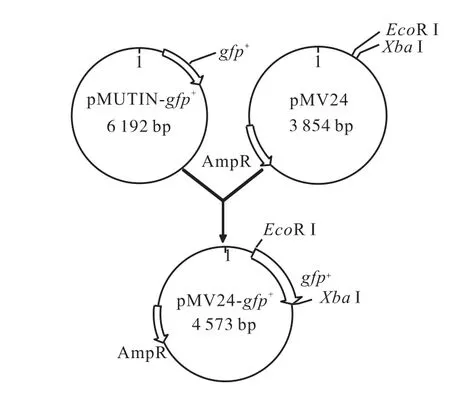
圖1 質粒構建流程圖Fig.1 Construction of pMV24-gfp+ plasmid
1.2.3 化學轉化
于感受態細菌中加入5.0,μL 連接產物,輕輕攪拌混勻,冰浴30,min;42,℃水浴熱激90,s,冰浴2,min;每管加LB 液體培養液900,μL,37,℃水浴60,min,180,r/min 振蕩培養1,h;將上述菌液按原倍和稀釋10倍分別涂布于含100,μg/mL 氨芐青霉素100,μL 和40,μg/mL 的X-Gal 的LB 平板,置于37,℃培養箱,37,℃正放至涂布液體基本被吸收,再倒置培養18~24,h.從平板上挑取單個分離良好的白色菌落.
1.2.4 木葡糖酸醋桿菌電轉化
取甘油管中保存的木葡糖酸醋桿菌的菌液接入裝有100,mL 液體培養基的500,mL 搖瓶中,30,℃、160,r/min 振蕩培養24,h,向液體搖瓶中加入1,mL 纖維素酶(10,000,U/mL),30,℃酶解2,h.酶解后離心洗滌后的細胞,經10%甘油-氯化鎂溶液洗滌、離心各2次,再以10%甘油懸浮、離心各2 次,最后用10%甘油溶液懸浮制成感受態細胞.電擊條件為:電壓2.40,kV、電容25,μF、電阻200,Ω.在含氨芐青霉素100,μg/mL 的抗性固體平板上篩選轉化子.
1.2.5 轉基因芽的GFP 熒光觀察
在BX51 型熒光顯微鏡下觀察轉基因的細胞中GFP 熒光的表達情況,采用自動成像系統拍照.
2 結果與分析
2.1 目的基因的擴增結果與克隆載體的構建
以質粒pMUTIN-gfp+為模板擴增gfp 基因,擴增的片段與表達載體pMV24 用EcoRⅠ和XbaⅠ雙酶切,結果如圖2 所示.從電泳結果看,在PCR 擴增的gfp 基因與預期大小相符.經切膠回收,連接pCR2.1 T 載體,轉化到大腸桿菌DH5α.陽性克隆測序結果分析顯示gfp 基因片段與pMUTIN-gfp+的目的片段序列大小(717,bp)一致,二者的堿基序列相同.
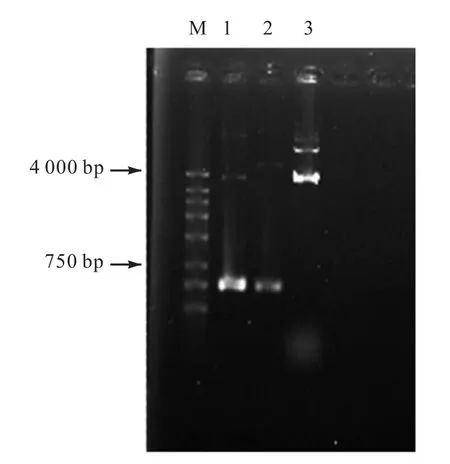
圖2 目的基因和穿梭質粒的雙酶切電泳圖Fig.2 Electrophoretogram of target gene and shuttle plasmid with restriction enzyme digestion
2.2 重組質粒的構建和鑒定
將EcoRⅠ和XbaⅠ雙酶切處理后的pCR2.1,T/gfp+質粒,通過瓊脂糖凝膠電泳分離T 載體片段和gfp 目的片段,將gfp 基因回收后,與pMV24 按3∶1和5∶1 體系分別過夜連接,并轉化至大腸桿菌感受態細胞中.挑取綠色菌落,提取質粒并采用EcoRⅠ和XbaⅠ雙酶切驗證,結果如圖3 所示.由圖3(b)可以看出,在4,000,bp 和750,bp 的DNA Marker 條帶附近有與預期大小抑制的條帶出現.
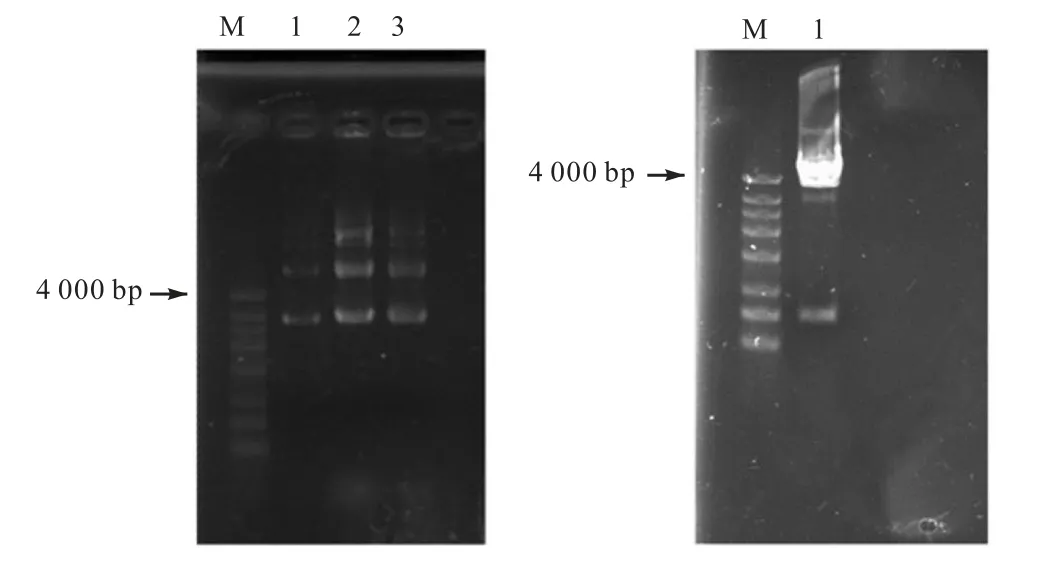
圖3 gfp+轉化子的鑒定Fig.3 Identification of gfp+ transformation
2.3 GFP在G.xylinus CGMCC 2955中的表達
將構建好的質粒pMV24-gfp+電轉化至木葡糖酸醋桿菌中.將轉化菌株置于熒光顯微鏡下進行觀察,結果如圖4 所示.轉化菌株在熒光顯微鏡的藍色激發光下可發出綠色熒光,拍照可觀察到菌體發出綠色的熒光,說明重組質粒pMV24-gfp+在木葡糖酸醋桿菌中表達,證明其可作為選擇標記用于今后木葡糖酸醋桿菌的趨化性研究.
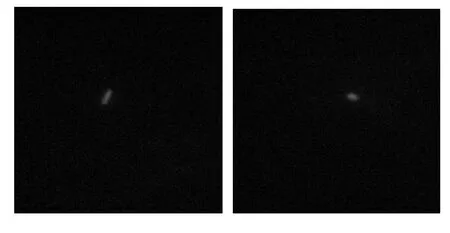
圖4 轉化子熒光顯微照片Fig.4 Fluorescence micrograph of transformant
3 討論
作為一種報告基因,GFP 比lacZ、CAT 等報告基因有許多無可比擬的優越性:GFP 不具有種屬依賴性,在多種原核和真核生物細胞中都表達;不需要反應底物與其他輔助因子,在藍光的激發下可產生綠色熒光;通過替換特性氨基酸,可產生不同顏色的光,滿足不同的研究需求等[6].因而,GFP 可廣泛應用于報告基因、基因的表達與調控、蛋白質的定位、信號傳導、轉染與轉化以及細胞的分離與純化等領域[7],可在更接近自然真實狀態的條件下進行活細胞實時定位觀察.如在活細胞中直接觀察蛋白質向細胞核、內質網運動的狀態,還可實時觀察到外界信號刺激下目的蛋白的變化過程.
然而,蛋白在原核表達系統中表達時,常常受到目的基因本身結構、轉錄水平調控和蛋白質折疊等諸多因素的影響.目的基因在原核表達系統的有效表達成為基因工程技術的難點之一,制約了蛋白質的功能研究和應用.本文所采用的穿梭質粒pMV24 是由日本Mizknan Holdings 中央研究院構建而成,目前已廣泛用作醋酸菌屬的穿梭質粒和表達質粒[8].Fukay等[8]使用pMV24 穿梭質粒研究Acetobacter polyoxogenes 中乙醛脫氫酶對醋酸產量的影響.本文也成功使用該質粒使GFP 蛋白在木葡糖酸醋桿菌中表達.此外,pMV24 質粒中的氨芐青霉素抗性篩選基因也正是木葡糖酸醋桿菌所缺失的.因而,陽性克隆細胞的篩選簡便、成功率高.
目前,由于木葡糖酸醋桿菌的定向運動可直接引起細菌纖維素的規則性排列,使得木葡糖酸醋桿菌的趨化性廣泛受到研究者的關注.孫臻[9]利用熒光染料coriphosphine O 對木醋桿菌(Acetobacter xylinus)進行染色,以觀察微生物細胞在單根條紋管道和網格狀微流控芯片中的運動.然而,其染色過程通常伴隨著對菌體的離心和洗滌等多個處理程序,往往造成微生物細胞的損傷.而GFP 無毒性且是菌體在不受任何外界環境影響的前提下自發表達的,可在最自然的狀態下追蹤細胞的運動.或者,將GFP 與其他蛋白融合追蹤蛋白的變化過程.Englert 等[10]就曾用表達綠色熒光蛋白和紅色熒光蛋白的大腸桿菌對趨化性進行了定量分析.Sourjik 等[11-12]用熒光共振能量轉移技術(fluorescence resonance energy transfer,FRET)研究了趨化信號傳導系統中各種蛋白的相互作用.
綜上所述,將GFP 基因在木葡糖酸醋桿菌中成功表達是追蹤菌體細胞運動和胞內蛋白遷移的最佳選擇,還可用于檢測基因表達的時序性,例如觀察使木葡糖酸醋桿菌具有趨化反應的鞭毛及其動力蛋白等分子發動機的形成.因此,本實驗采用PCR 技術擴增出綠色熒光蛋白,以pMV24 穿梭質粒為載體,實現了GFP 綠色熒光蛋白基因在木葡糖酸醋桿菌中的表達;在熒光顯微鏡的藍色激發光下可觀察到綠色熒光;此轉化子的成功構建為木葡糖酸醋桿菌趨化性的深入研究奠定了堅實的研究基礎.
[1]Morgan J L W,McNamara J T,Zimmer J.Mechanism of activation of bacterial cellulose synthase by cyclic di-GMP[J].Nature Structural &Molecular Biology,2014,21(5):489-496.
[2]李晶,賈士儒,楊洪江,等.木葡糖酸醋桿菌趨化性的初步研究[J].天津科技大學學報,2012,27(1):1-4.
[3]Sano M B,Rojas A D,Gatenholm P,et al.Electromagnetically controlled biological assembly of aligned bacterial cellulose nanofibers[J].Annals of Biomedical Engineering,2010,38(8):2475-2484.
[4]Shimonura O.Structure of the chromophore of Aequorea green fluorescent protein[J].FEBS Letters,1979,104(2):220-222.
[5]Prasher D C,Eckenrode V K,Ward W,et al.Primary structure of the Aequorea victoria green fluorescent protein[J].Gene,1992,111(2):229-233.
[6]Cubitt A B,Heim R,A dams S R,et al.Understanding,improving and using green fluorescent protein[J].Ttrends in Biochemical Sciences,1995,20(11):448-455.
[7]Kain S R,Adams M,Kondepudi A,et al.Green fluorescent protein as a reporter of gene expression and protein localization[J].BioTechniques,1995,19(4):650-655.
[8]Fukaya M,Tayama K,Tamaki T,et al.Cloning of the membrane-bound aldehyde dehydrogenase gene of Acetobacter polyoxogenes and improvement of acetic acid production by use of the cloned gene[J].Applied and Environmental Microbiology,1989,55(1):171-176.
[9]孫臻.圖案化細菌纖維素的可控生物制造[D].武漢:華中科技大學,2013.
[10]Englert D L,Manson M D,Jayaraman A.Flow-based microfluidic device for quantifying bacterial chemotaxis in stable[J].Competing Gradients Applied and Environmental Microbiology,2009,75(13):4557-4564.
[11]Sourjik V,Berg H C.Receptor sensitivity in bacterial chemotaxis[J].Proceedings of the National Academy of Sciences of the United States of America,2002,99(1):123-127.
[12]Sourjik V,Berg H C.Functional interactions between receptors in bacterial chemotaxis[J].Nature,2004,428(6981):437-441.
猜你喜歡
品牌研究(2022年29期)2022-10-15 08:01:00
品牌研究(2022年26期)2022-09-19 05:54:46
品牌研究(2021年31期)2022-01-25 06:41:46
品牌研究(2021年36期)2022-01-21 09:29:28
品牌研究(2021年34期)2022-01-18 10:54:04
品牌研究(2021年35期)2022-01-18 08:45:18
品牌研究(2021年20期)2022-01-11 04:07:56
品牌研究(2021年27期)2022-01-04 06:41:14
品牌研究(2021年17期)2021-12-09 05:06:54
海峽姐妹(2019年3期)2019-06-18 10:37:10
