不同鹽脅迫對柳枝稷生物量、品質和光合生理的影響
2015-01-19 07:06:37趙春橋李繼偉范希峰侯新村武菊英胡躍高劉吉利
生態學報 2015年19期
趙春橋, 李繼偉, 范希峰, 侯新村, 武菊英, 胡躍高, 劉吉利
1 中國農業大學農學與生物技術學院, 北京 100193 2 北京市農林科學院草業與環境研究發展中心, 北京 100097 3 寧夏大學新技術應用研究開發中心, 銀川 750021
不同鹽脅迫對柳枝稷生物量、品質和光合生理的影響
趙春橋1,2, 李繼偉2, 范希峰2, 侯新村2, 武菊英2, 胡躍高1,*, 劉吉利3
1 中國農業大學農學與生物技術學院, 北京 100193 2 北京市農林科學院草業與環境研究發展中心, 北京 100097 3 寧夏大學新技術應用研究開發中心, 銀川 750021
為明確不同鹽脅迫對柳枝稷生物量、品質及光合生理的影響,以無鹽脅迫作為對照(CK),選取0.40% NaCl、0.80% Na2SO4和0.80% NaHCO3進行了土柱試驗。結果表明:(1)與CK相比,NaCl、Na2SO4、NaHCO3脅迫下柳枝稷地上生物量、地下生物量、總生物量、籽粒產量及根冠比均顯著降低(P< 0.05),總生物量分別降低49.39%、60.52%、76.45%,NaHCO3對柳枝稷的生長抑制作用最強,NaCl最弱;(2)NaCl脅迫下柳枝稷地上生物質灰分含量顯著增高14.89%,Na2SO4脅迫下硫(S)含量顯著增高262.32%,纖維素含量顯著降低13.71%,NaHCO3脅迫下鉀(K)含量顯著增高54.95%,半纖維素含量顯著增高10.87%,灰分和S含量的增高不利于柳枝稷地上生物質的燃燒利用,纖維素含量的降低和半纖維素含量的增高不利于其轉化利用;(3)NaCl、Na2SO4、NaHCO3脅迫下柳枝稷葉片凈光合速率(Pn)分別顯著降低21.89%、29.54% 和24.59%,氣孔限制因素可能是其光合作用受到抑制、生物量下降的關鍵因素。
柳枝稷; 鹽脅迫; 生物量; 品質; 光合生理; 邊際土地
利用邊際土地種植能源植物,是我國獲取生物質原材料、發展生物質能產業的一條重要途徑[1]。鹽漬化土地是我國典型的邊際土地類型,總面積約為1.30×107hm2,且仍不斷增加[2]。在鹽漬邊際土地種植能源植物,既可有效緩解生物質原材料供應不足問題,又可改善鹽漬地區生態環境[3]。
柳枝稷(PanicumvirgatumL.)原產于北美地區,禾本科黍屬,系多年生高大叢生C4草本植物,具有抗旱、抗寒、耐鹽堿、貧瘠等多種逆境脅迫的能力,適應范圍廣,生物質產量高,易于管理、收獲[4]。20世紀80年代柳枝稷被引種到中國,適應性強,逐漸受到人們關注[5]。柳枝稷苗期耐鹽性良好,具有較好的研發潛力[6]。水培條件下柳枝稷幼苗生物量下降 50% 的鹽濃度為10.44 g/L[7]。以生物量為衡量指標,柳枝稷在不同土壤鹽脅迫下的適宜濃度范圍分別為:NaHCO3≤ 0.40%,Na2SO4≤ 0.60%,NaCl ≤ 0.20%[8]。不同土壤鹽脅迫顯著影響了柳枝稷植株生長與根系垂直分布[9]。鹽脅迫與pH在對柳枝稷生長的影響上存在著協同或拮抗作用[10]。對柳枝稷鹽脅迫下抗氧化指標的模型構建方便了人們對其耐鹽性的研究[11]。由此可見,人們對鹽脅迫下柳枝稷幼苗生長、發育、根系分布及耐鹽評價模型等進行了研究,而不同鹽脅迫對全生育期柳枝稷生物量、品質及光合生理的影響尚不清楚。
本文根據前期研究基礎得出保證柳枝稷幼苗成活最大土壤NaCl、Na2SO4和NaHCO3濃度(以下簡稱臨界致死濃度)分別為0.40%、0.80% 和0.80%[12]。基于此,為明確不同鹽脅迫對全生育期柳枝稷生物量、品質及光合生理的影響,本文選取0.40% NaCl、0.80% Na2SO4、0.80% NaHCO3進行了土柱試驗。
1 材料與方法
1.1 供試材料
試驗于2009年在北京草業與環境研究發展中心人工防雨棚內進行,供試柳枝稷品種為Alamo,其種子于2008年11月采集于北京草業與環境研究發展中心能源草種植基地(N39°34′,E116°28′)。土壤基質為潮褐土,有機質含量為1.72%,速效N、速效P、速效K含量分別為84.00 mg/kg,46.35 mg/kg,127.00 mg/kg,土壤pH為7.42。
1.2 試驗設計
本文采用配對設計[13],土柱法[14]開展試驗。土壤鹽分類型及質量百分比分別為:0.40% NaCl、0.80% Na2SO4和0.80% NaHCO3,無鹽脅迫作為對照(CK),按質量百分比與過篩后的壤土充分混勻,試驗設3重復。土柱直徑40 cm、高120 cm,管壁下部設有10個小孔(孔徑大小1 cm),置于120 cm深的坑內,保持土柱內土面與地面齊平。每個土柱配有直徑50 cm的桶(桶內保證有10 cm深的水),以防止鹽分流失。2009年6月13日將3葉期柳枝稷幼苗移栽后首次灌足安家水12.70 L,之后維持桶內水深10 cm。鹽脅迫處理1個月后測定各生理指標,至2009年11月4日試驗結束,取樣,測定柳枝稷生物量、品質指標。
1.3 測定項目和方法
(1)生物量 取樣后將柳枝稷地上與地下部分開、洗凈后裝入紙袋并于105℃殺青30 min,80℃烘干至恒重,稱重。柳枝稷籽粒采用人工收獲,去除稃和穎后80℃下烘干至恒重,稱重。
(2)光合參數 采用便攜式光合系統測定儀LI-6400 (LI-COR Lincoln,USA)于晴朗的9:00—11:00選取柳枝稷第3片成熟葉片進行連體測定,所測葉片完整無損并使其保持自然取向,每葉片重復記錄3組數據,結果取其平均值。光合有效輻射(PAR)為1100—1200 μmol m-2s-1。光響應曲線的測定由紅藍光源(Li-6400-02B)提供不同的PAR: 2000、1800、1500、1000、800、500、200、100、50、20、0 μmol m-2s-1。CO2濃度設定為 400 μmol/mol,流速為400 μmol/s、葉室(2 cm × 3 cm)溫度設定為(30 ± 1)℃。采用直角雙曲線模型進行響應曲線模擬,并計算光補償點LCP與光飽和點LSP[15]。
(3)水分利用效率(WUE) 采用LI-6400 (LI-COR Lincoln,USA)測定得到Pn和Ci后計算得到。計算公式為:WUE =Pn/Ci。
(4)生物質品質 將烘干至恒重的柳枝稷材料粉碎并過40目篩,存放于干燥器中待測。采用直接灰化法測定灰分,馬弗爐測定揮發分,N、P、K采用H2SO4-H2O2消煮-比色法測定,S含量采用H2NO4-HClO4消煮-流動注射分析儀法測定,Cl含量采用莫爾法測定,Si含量采用H2SO4-H2O2消煮-重量法測定[16],熱值采用XRY-1C型氧彈式熱量計測定,洗滌法測定纖維素,半纖維素和木質素含量[17]。
1.4 統計分析
采用PAIRED T-TEST(SAS 8.2)進行差異顯著性檢驗,Excel 2003和Origin7.0統計軟件進行數據分析和曲線擬合。
2 結果與分析
2.1 不同鹽脅迫對柳枝稷生物量的影響
NaCl、Na2SO4和NaHCO3脅迫下,柳枝稷生長受到顯著抑制(表1),就生物量而言,與CK相比,柳枝稷地上生物量分別降低56.14%、61.73%、76.90%,地下生物量分別降低36.12%、58.67%、77.57%,總生物量分別降低49.39%、60.52%、76.45%,根冠比分別降低25.00%、31.25%、32.50%,籽粒產量分別降低70.95%、52.88%、33.81%。3種鹽脅迫之間比較,柳枝稷生長受抑制程度顯著不同,以NaHCO3抑制作用最強,而NaCl抑制作用最弱。就籽粒產量而言,NaCl脅迫下籽粒產量最低,質量百分含量為0.92%,NaHCO3脅迫下籽粒產量最高,質量百分含量為4.50%;就根冠比而言,3種鹽脅迫間差異并不顯著。
2.2 不同鹽脅迫對柳枝稷地上生物質品質的影響
2.2.1 不同鹽脅迫對柳枝稷地上生物質燃燒特性的影響
不同鹽脅迫下,柳枝稷地上生物質燃燒特性的變化較小(表2)。NaCl脅迫下,柳枝稷地上生物質灰分含量顯著增高14.89%。其余兩種鹽脅迫對柳枝稷地上生物質燃燒特性的影響均不顯著。3種鹽脅迫之間比較,NaCl脅迫下,柳枝稷地上生物質灰分含量顯著高于其他兩種鹽脅迫,其他指標則無顯著差異。

表1 不同鹽脅迫下柳枝稷生物量
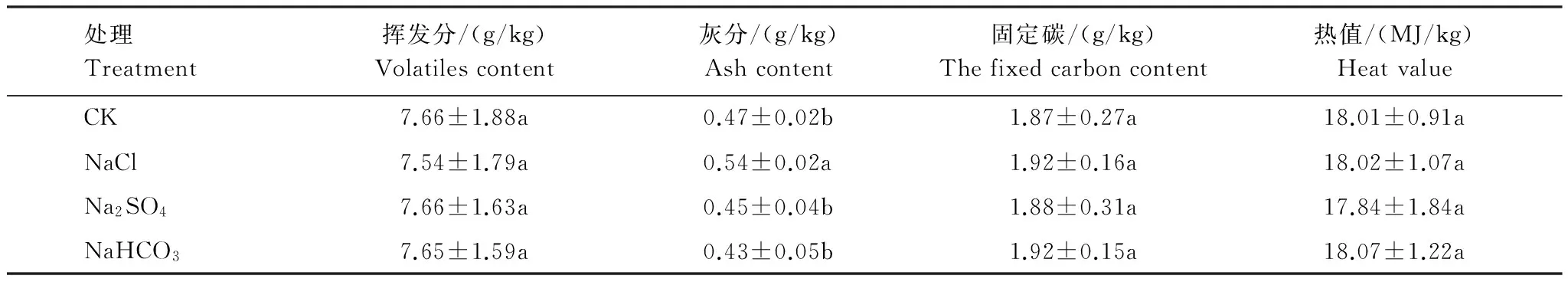
表2 不同鹽脅迫下柳枝稷地上生物質燃燒特性
2.2.2 不同鹽脅迫對柳枝稷地上生物質礦質元素含量的影響
不同鹽脅迫下柳枝稷地上生物質礦質元素含量存在差異(表3)。與CK相比,3種鹽脅迫下柳枝稷地上生物質N、P、Cl、Si含量無顯著差異。Na2SO4脅迫下,S含量顯著增高262.32%;NaHCO3脅迫下,K含量顯著降低54.95%。3種鹽脅迫間比較,Na2SO4脅迫下柳枝稷地上生物質S含量顯著高于其他兩種鹽脅迫,NaHCO3脅迫下K含量顯著低于其余兩種鹽脅迫。
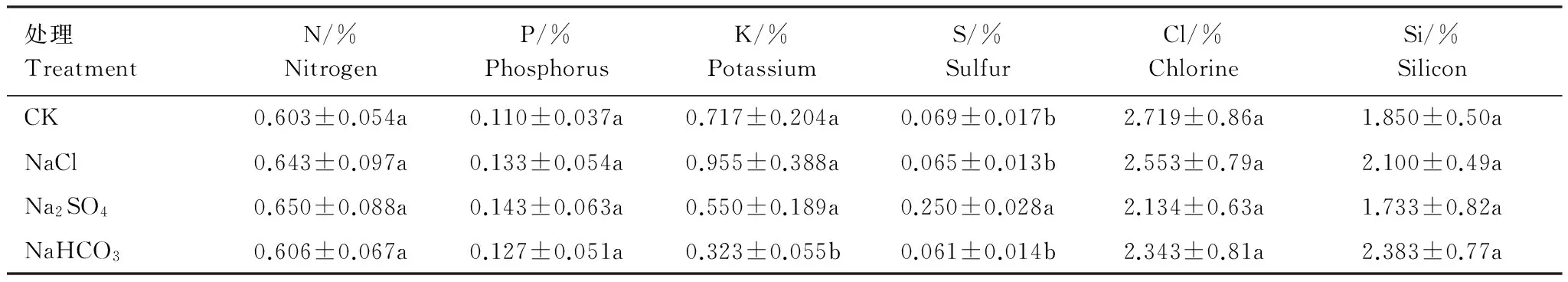
表3 不同鹽脅迫下柳枝稷地上生物質礦質元素含量(干物質)
2.2.3 不同鹽脅迫對柳枝稷地上生物質細胞壁組分含量的影響
與CK相比,不同鹽脅迫下柳枝稷地上生物質纖維素和木質素含量都呈現出降低的趨勢,半纖維素含量呈現出增高的趨勢(表 4),但只有Na2SO4脅迫下纖維素含量顯著降低13.71% NaHCO3脅迫下,半纖維素含量顯著增高10.87%。3種鹽脅迫間比較,Na2SO4脅迫下,纖維素含量顯著高于其他兩種鹽脅迫,NaHCO3脅迫下,半纖維素含量顯著高于其余兩種鹽脅迫。

表4 不同鹽脅迫下柳枝稷地上生物質細胞壁組分含量
2.3 不同鹽脅迫對柳枝稷葉片光合生理特征的影響
2.3.1 不同鹽脅迫對柳枝稷光合參數的影響
不同鹽脅迫對柳枝稷光合參數的影響不同(表 5),與CK相比,NaCl、Na2SO4、NaHCO3脅迫下柳枝稷Pn、Gs、Ci、Tr、Ls和SUE均顯著降低,WUE變化并不顯著。NaCl和Na2SO4脅迫下CUE 顯著增高。3種鹽脅迫間比較,Na2SO4脅迫下WUE顯著高于NaCl脅迫,其余指標則無顯著差異。
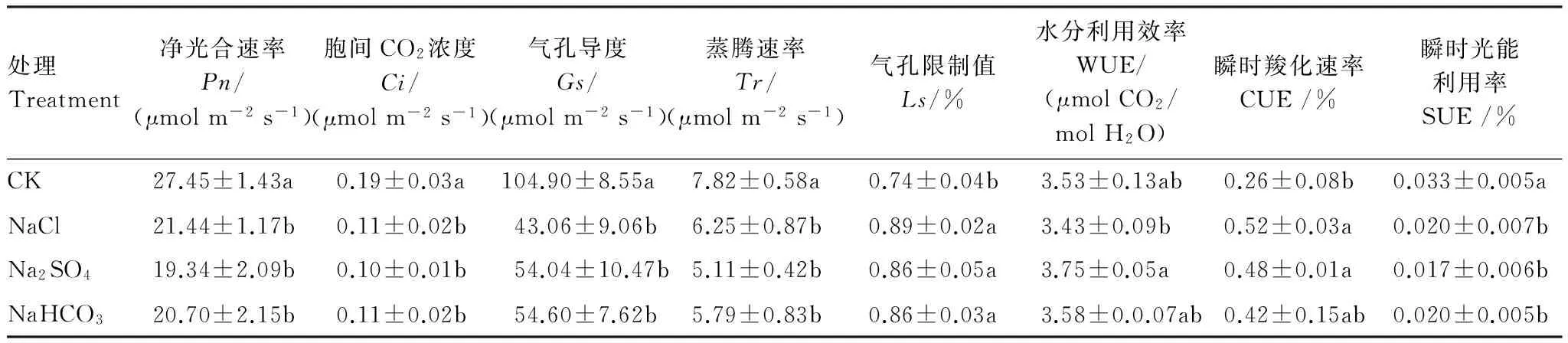
表5 不同鹽脅迫下柳枝稷光合參數
Pn: net photosynthetic rate,Ci: intercellular carbon dioxide concentration,Gs: stomatal conductance,Tr: transpiration rate,Ls: limiting value of stomata, WUE: water use efficiency, CUE: Instantaneous carboxylation rate, SUE: instantaneous light energy utilization efficiency
2.3.2 不同鹽脅迫對柳枝稷光合-光強響應曲線特征參數的影響
不同鹽脅迫下,柳枝稷的Pn、Ci、Tr和WUE隨PAR的增高呈現出較為一致的變化趨勢。均表現為較低PAR時各項指標值迅速增高然后增幅逐漸平緩。本文采用直角雙曲線模型對光合-光強響應曲線進行模擬,結果顯示各方程的決定系數都在0.99以上,表明該模型能較好地反應葉片光合對光強的響應過程。
與對照相比,NaCl脅迫下,柳枝稷葉片Pmax、Rd、LSP均分別顯著降低14.52%、10.52%、12.59%(表 6);Na2SO4脅迫下,柳枝稷葉片AQY顯著增高22.73%,Pmax、LCP和LSP分別顯著降低10.00%、17.41%、30.43%;NaHCO3脅迫下柳枝稷葉片Pmax、Rd、LSP分別顯著降低4.10%、6.38%、20.92%。其余指標變化不顯著。3種鹽脅迫之間比較,Na2SO4脅迫下AQY、Rd顯著高于其余兩種鹽脅迫,LCP顯著低于其余兩種鹽脅迫。
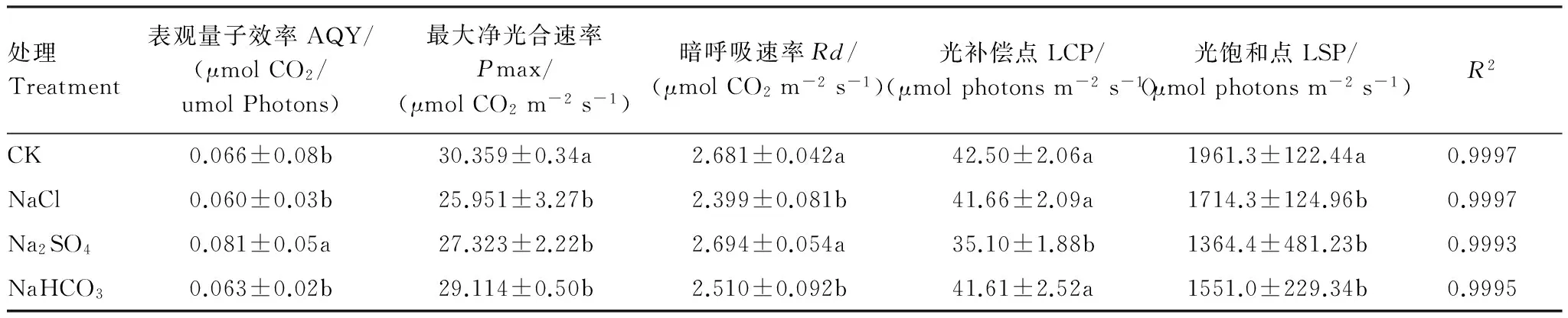
表6 直角雙曲線修正模型擬合的不同鹽脅迫處理柳枝稷光強響應參數
3 討論
鹽脅迫嚴重影響了植物的生長發育[18]。本研究表明,不同鹽脅迫顯著降低了柳枝稷地上生物量、地下生物量、總生物量、籽粒產量和根冠比,這與前人的研究結果相似[19]。不同鹽脅迫對柳枝稷生長的抑制作用顯著不同,以NaHCO3脅迫抑制作用最強。有研究表明鹽分與pH在對柳枝稷的影響方面存在著協同與拮抗關系[10]。本研究表明,NaHCO3較其他兩種鹽對柳枝稷生長的抑制作用最強,這可能是由于NaHCO3與pH共同抑制了柳枝稷的生長。有報道表明,有些植物可通過生物量分配模式的調整來適應鹽脅迫環境[20-21]。不同鹽脅迫下,柳枝稷植株營養器官與生殖器官生物量的分配體現出顯著差異,這可能是柳枝稷適應鹽脅迫環境的一種策略。
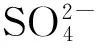
鹽脅迫下,引起植物葉片光合效率降低的因素主要有兩種,一種是氣孔限制因素,表現為Ls顯著增高,Gs下降,CO2進入葉片受阻,Ci降低,導致Pn降低[27]。另一種是非氣孔限制因素,鹽脅迫導致光合機構受損,電子傳遞速率下降,Pn、Tr、Gs顯著降低,Ci升高,從而影響同化力的形成[28]。本研究表明,不同鹽脅迫下,柳枝稷葉片蒸騰速率顯著降低,氣孔限制值顯著增高,CO2進入葉片受阻,SUE顯著下降,從而導致Pn和Pmax顯著降低。由此可見,鹽脅迫造成的氣孔限制可能是導致柳枝稷光合速率降低、生物量下降的關鍵因素。不同鹽脅迫間比較,此種抑制作用并未表現出顯著性差異。
4 結論
(1) 不同鹽脅迫顯著抑制了柳枝稷的生長,使其生物量顯著降低,生物量分配顯著變化。
(2) 不同鹽脅迫對柳枝稷地上生物質品質影響有限,NaCl脅迫下柳枝稷地上生物質灰分含量的顯著增高和Na2SO4脅迫下S含量的顯著增高對其燃燒利用不利,Na2SO4脅迫下纖維素含量的顯著降低和NaHCO3脅迫下半纖維素含量的顯著增高對其轉化利用不利。
(3) 不同鹽脅迫導致的氣孔限制因素可能是導致柳枝稷生物量降低的關鍵因素。
[1] Tang Y, Xie J S, Geng S. Marginal land-based biomass energy production in China. Journal of Integrative Plant Biology, 2010, 52(1): 112-121.
[2] 王繼和, 楊自輝, 胡明貴, 劉虎俊, 紀永福. 干旱區鹽漬化土地綜合治理技術研究. 中國生態農業學報, 2001, 9(1): 64-66.
[3] 侯新村, 范希峰, 武菊英, 左海濤. 纖維素類能源草在京郊地區的經濟效益與生態價值評價. 草業學報, 2011, 20(6): 12-17.
[4] Sanderson M, Reed R L, McLaughlin S B, Wullschleger S D, Conger B V, Parrish D J, Wolf D D, Taliaferro C, Hopkins A A, Ocumpaugh W R, Hussey M A, Read J C, Tischler C R. Switchgrass as a sustainable bioenergy crop. Bioresource Technology, 1996, 56(1): 83-93.
[5] 范希峰, 候新村, 左海濤, 武菊英, 段留生. 邊際土地類型及移栽方式對柳枝稷苗期生長的影響. 草業科學, 2010, 27(1): 97-102.
[6] 范希峰, 侯新村, 朱毅, 武菊英. 鹽脅迫對柳枝稷苗期生長和生理特性的影響. 應用生態學報, 2012, 23(6): 1476-1480.
[7] 于曉丹, 杜菲, 張蘊薇. 鹽脅迫對柳枝稷種子萌發和幼苗生長的影響. 草地學報, 2012, 18(6): 810-815.
[8] 左海濤, 李繼偉, 郭斌, 宋尚有, 高旺盛. 鹽分和土壤含水量對營養生長期柳枝稷的影響. 草地學報, 2009, 17(6): 760-766.
[9] 李繼偉, 左海濤, 李青豐, 范希峰, 侯新村. 柳枝稷根系垂直分布及植株生長對土壤鹽分類型的響應. 草地學報, 2011, 19(4): 644-651.
[10] Liu Y, Wang Q Z, Zhang Y W, Cui J, Chen G, Xie B, Wu C H, Liu H T. Synergistic and antagonistic effects of salinity and pH on germination in switchgrass (PanicumvirgatumL.). PloS ONE, 2014, 9(1): e85282.
[11] Wang Q Z, Wu C H, Xie B, Liu Y, Cui J, Chen G, Zhang Y W. Model analysing the antioxidant responses of leaves and roots of switchgrass to NaCl-salinity stress. Plant Physiology and Biochemistry, 2012, 58: 288-296.
[12] 李繼偉. 栽培管理措施和環境脅迫對柳枝稷生長特性和生物質品質的影響[D]. 內蒙古: 內蒙古農業大學, 2011.
[13] Mass E V, Poss J A. Salt sensitivity of wheat at various growth stages. Irrigation Science, 1989, 10(1): 29-40.
[14] 黃燕, 吳平. SAS統計分析及應用. 北京: 機械工業出版社, 2007: 67-98.
[15] Farquhar G D, Von C S, Berry J A. Models of photosynthesis. Plant Physiology, 2001, 125(1): 42-45.
[16] 鮑士旦. 土壤農化分析. 北京: 中國農業出版社, 2008: 257-282.
[17] Van Soest P J, Robertson J B, Lewis B A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal of Dairy Science, 1991, 74(10): 3583-3597.
[18] Zhu J K. Plant salt tolerance. Trends in Plant Science, 2001, 6(2): 66-71.
[19] Azevedo Neto A D, Prisco J T, Enéas-Filho J, Lacerda C F D, Silva J V, Costa P H A D, Gomes-Filho E. Effects of salt stress on plant growth, stomatal response and solute accumulation of different maize genotypes. Brazilian Journal of Plant Physiology, 2004, 16(1): 31-38.
[20] Mauchamp A, Mésleard F. Salt tolerance inPhragmitesaustralispopulations from coastal Mediterranean marshes. Aquatic Botany, 2001, 70(1): 39-52.
[21] Van Zandt P A, Tobler M A, Mouton E, Hasenstein K H, Mopper S. Positive and negative consequences of salinity stress for the growth and reproduction of the clonal plant,Irishexagona. Journal of Ecology, 2003, 91(5): 837-846.
[22] Hoson T, Soga K, Mori R, Saiki M, Nakamura Y, Wakabayashi K, Kamisaka S. Stimulation of elongation growth and cell wall loosening in rice coleoptiles under microgravity conditions in space. Plant and Cell Physiology, 2002, 43(9): 1067-1071.
[23] Piro G, Leucci M R, Waldron K, Dalessandro G. Exposure to water stress causes changes in the biosynthesis of cell wall polysaccharides in roots of wheat cultivars varying in drought tolerance. Plant science, 2003, 165(3): 559-569.
[24] 裴惠娟, 張滿效, 安黎哲. 非生物脅迫下植物細胞壁組分變化. 生態學雜志, 2011, 30(6): 1279-1286.
[25] Cheeseman J M. Mechanisms of salinity tolerance in plants. Plant Physiology, 1988, 87(3): 547-550.
[26] 林舜華, 黃銀曉, 蔣高明, 韓榮莊, 高雷明. 海河流域植物硫素含量特征的研究. 生態學報, 1994, 14(3): 235-242.
[27] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33: 317-345.
[28] 許大全. 光合作用效率. 上海: 上海科學技術出版社, 2002: 32-34.
Effects of salt stress on biomass, quality, and photosynthetic physiology in switchgrass
ZHAO Chunqiao1,2, LI Jiwei2, FAN Xifeng2, HOU Xincun2, WU Juying2, HU Yuegao1,*, LIU Jili3
1CollegeofAgricultureandBiotechnology,ChinaAgriculturalUniversity,Beijing100193,China2Research&DevelopmentCenterforGrassandEnvironment,BeijingAcademyofAgriculturalandForestrySciences,Beijing100097,China3ResearchandDevelopmentCenterofNewTechnologyApplication,NingxiaUniversity,Yinchuan750021,China
In order to study the effects of salt stress on biomass, quality, and photosynthetic characteristics in switchgrass, a soil column test was performed with different salt stresses: 0.40% NaCl, 0.80% Na2SO4, and 0.80% NaHCO3(critical lethal concentration). Switchgrass with no salt stress served as control (CK). The results obtained are as follows: (1) The aboveground biomass, underground biomass, total biomass, seed yield, and root cap ratio of switchgrass decreased significantly (P< 0.05) under salt stresses of 0.40% NaCl, 0.80% Na2SO4, and 0.80% NaHCO3compared to the CK. Aboveground biomass of switchgrass was 56.14%, 61.73%, and 76.90% lower than the CK, respectively; underground biomass was 36.12%, 58.67%, and 77.57% lower than the CK; total biomass was 49.39%, 60.52%, and 76.45% lower than the CK; seed yields were 70.95%, 52.88%, and 33.81% lower than the CK; and the root cap ratios were 25.00%, 31.25%, and 32.50% lower than the CK. The relative seed mass of switchgrass was 0.92%, 1.91%, and 4.50% of the control under 0.40% NaCl, 0.80% Na2SO4, and 0.80% NaHCO3, respectively. Large differences were observed among salt stresses in the degree to which growth was inhibited, although all were under critical lethal concentrations. Salt stress with 0.80% NaHCO3caused the most serious negative impact on switchgrass biomass, and the weakest was with 0.40% NaCl. Biomass distribution between vegetative parts and sexual organs varied greatly with salt stress. (2) Grown with 0.40% NaCl, the ash content of aboveground biomass increased 14.89% compared to the CK. Grown with 0.80% Na2SO4, the sulfur (S) content of aboveground biomass increased 262.32% and cellulose content decreased 13.71%. With 0.80% NaHCO3, potassium (K) content increased 54.95% and hemicellulose content increased 10.87%. All five of these differences were statistically significant. Other quality indexes showed no significant differences, either among salt stresses or in comparison to CK values. The increased ash content and S content observed under 0.40% NaCl and 0.80% Na2SO4negatively affected the biomass combustion quality. The reduction in cellulose content and increase in hemicellulose content rendered the transformation and utilization of switchgrass biomass more difficult. (3) Grown with 0.40% NaCl, 0.80% Na2SO4, and 0.80% NaHCO3, the net photosynthetic rate (Pn) and the maximum photosynthetic rate (Pmax) of switchgrass leaves was inhibited significantly compared to the CK.Pndecreased 21.89%, 29.54%, and 24.59%, respectively, andPmaxdecreased 14.52%, 10.00%, and 4.1%, respectively. Stomatal conductance (Gs), intercellular CO2concentration (Ci), transpiration rate (Tr), light saturation point (LSP), and instantaneous light energy utilization efficiency (SUE) all decreased significantly compared to the CK. The limiting value of stomata (Ls) increased 20.27%, 16.22%, and 16.22%, respectively. The apparent quanta efficiency (AQY), respiration rate (Rd), instantaneous carboxylation efficiency (CUE), light compensation point (LCP), and water use efficiency (WUE) showed different magnitudes of change under different salt stresses. Stoma limitation may be the key factor inhibiting the photosynthesis and growth of switchgrass. This study lays a solid foundation for the large-scale planting of switchgrass in saline marginal land in northern and northwestern China.
switchgrass; salt stress; biomass; quality; photosynthetic physiology; marginal land
國家自然科學基金(31101103); 北京市農林科學院科技創新能力建設專項(KJCX201201001); 北京市農林科學院青年科研基金(QNJJ201219)
2014-02-17; < class="emphasis_bold">網絡出版日期:
日期:2014-12-04
10.5846/stxb201402170269
*通訊作者Corresponding author.E-mail: huyuegao@cau.edu.cn
趙春橋, 李繼偉, 范希峰, 侯新村, 武菊英, 胡躍高, 劉吉利.不同鹽脅迫對柳枝稷生物量、品質和光合生理的影響.生態學報,2015,35(19):6489-6495.
Zhao C Q, Li J W, Fan X F, Hou X C, Wu J Y, Hu Y G, Liu J L.Effects of salt stress on biomass, quality, and photosynthetic physiology in switchgrass.Acta Ecologica Sinica,2015,35(19):6489-6495.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
