核糖核酸開關(guān)用于微生物細(xì)胞工廠的智能與精細(xì)調(diào)控
2015-02-14 09:33:48趙雨佳張根林周曉宏李春
化工學(xué)報(bào) 2015年10期
趙雨佳,張根林,周曉宏,李春
(北京理工大學(xué)生命學(xué)院生物工程系,北京 100081)
核糖核酸開關(guān)用于微生物細(xì)胞工廠的智能與精細(xì)調(diào)控
趙雨佳,張根林,周曉宏,李春
(北京理工大學(xué)生命學(xué)院生物工程系,北京 100081)
利用代謝工程與合成生物技術(shù)對細(xì)胞內(nèi)復(fù)雜的代謝網(wǎng)絡(luò)和調(diào)控網(wǎng)絡(luò)進(jìn)行重構(gòu)和改造,以建立合成新化合物或提高目標(biāo)產(chǎn)物產(chǎn)量的微生物細(xì)胞工廠是當(dāng)今綠色化工技術(shù)發(fā)展的方向之一。微生物代謝途徑的調(diào)控受環(huán)境和遺傳的雙重影響,細(xì)胞通過全局轉(zhuǎn)錄因子、信使分子和反饋抑制等方式響應(yīng)環(huán)境變化來維持細(xì)胞的內(nèi)穩(wěn)態(tài);同時細(xì)胞還受自身遺傳基因線路的調(diào)控,在轉(zhuǎn)錄、翻譯以及翻譯后修飾過程中調(diào)控特定基因的表達(dá)。核糖核酸開關(guān)是一類調(diào)控基因線路表達(dá)的RNA元件,通過與金屬離子、糖類衍生物、氨基酸、核酸衍生物以及輔酶等特異性配體結(jié)合發(fā)生的構(gòu)象變化,從而啟動或阻斷mRNA的轉(zhuǎn)錄、翻譯、拼接等過程來調(diào)控基因的表達(dá)。核糖核酸開關(guān)作為天然的生物感受器和效應(yīng)器通過人工設(shè)計(jì)可成為微生物細(xì)胞工廠智能化和精細(xì)化調(diào)控的分子工具,并在化工、醫(yī)藥、環(huán)保、食品等領(lǐng)域得到廣泛應(yīng)用。
核糖核酸開關(guān);微生物細(xì)胞工廠;精細(xì)調(diào)控;生物轉(zhuǎn)化;優(yōu)化;代謝工程;合成生物學(xué)
Key words:riboswitch;microbial cell factory;fine regulation;biotransformation;optimization;metabolic engineering;synthetic biology
引 言
微生物由于其種屬多樣性和遺傳多樣性,在38億年的進(jìn)化歷程中,形成了復(fù)雜的代謝途徑,可以生產(chǎn)包括大宗化學(xué)品、醫(yī)藥、燃料、塑料在內(nèi)的多種重要的醫(yī)藥和化工產(chǎn)品,現(xiàn)已利用谷氨酸棒狀桿菌成功生產(chǎn)絲氨酸[1]、賴氨酸[2]、谷氨酸[3]以及丁二酸[4]等大宗化學(xué)品;利用釀酒酵母生產(chǎn)青蒿素[5-6]、紫杉醇[7]、丹參酮[8]等多種名貴藥物;通過藍(lán)細(xì)菌生產(chǎn)乙醇[9]、正丁醇[10]等生物燃料以及通過大腸桿菌生產(chǎn)聚羥基脂肪酸酯(polyhydroxyalkanoate,PHA)[11]、聚羥基丁酸酯(polyhydroxybutyrate, PHB)[12]等可降解的生物塑料。利用微生物生產(chǎn)化工產(chǎn)品為解決能源危機(jī)和環(huán)境污染提供了新思路。然而天然的微生物因?yàn)楫a(chǎn)量低或者底物譜窄等原因往往不能滿足工業(yè)化生產(chǎn)的需要,因此,需要人為地對其代謝途徑進(jìn)行改造,通過重組和優(yōu)化,使微生物細(xì)胞內(nèi)的代謝流和能量流重新分配,成為服務(wù)于工業(yè)需求的微生物細(xì)胞工廠。
利用微生物細(xì)胞工廠生產(chǎn)醫(yī)藥和化工產(chǎn)品具有清潔、高效、經(jīng)濟(jì)附加值高等優(yōu)點(diǎn),已成為發(fā)展綠色工業(yè)的核心部分。然而長期的研究發(fā)現(xiàn),微生物細(xì)胞暴露在環(huán)境中會受到來自外界和內(nèi)部的多種信號刺激,為了維持細(xì)胞的內(nèi)穩(wěn)態(tài),微生物進(jìn)化出了復(fù)雜的調(diào)控網(wǎng)絡(luò)來感受信號,進(jìn)而調(diào)控相應(yīng)的代謝途徑。因此單純的引入外源代謝途徑或者酶往往會導(dǎo)致代謝失衡,不能得到目標(biāo)產(chǎn)物或者產(chǎn)量難以滿足工業(yè)需求。為了得到能滿足工業(yè)需求的微生物細(xì)胞工廠不僅要對代謝途徑進(jìn)行改造還要對相應(yīng)的調(diào)控系統(tǒng)進(jìn)行修飾。但由于其調(diào)控的復(fù)雜性、人們?nèi)狈υ诜肿铀缴蠈?xì)胞調(diào)控機(jī)制的了解,難以開發(fā)智能的表達(dá)調(diào)控工具,嚴(yán)重制約了微生物細(xì)胞工廠的發(fā)展。
細(xì)胞的調(diào)控網(wǎng)絡(luò)受環(huán)境和遺傳的雙重影響。由于微生物細(xì)胞生活在不斷變化的環(huán)境中,溫度、pH、營養(yǎng)等環(huán)境因素都會對細(xì)胞的生理代謝產(chǎn)生影響,細(xì)胞通過各種全局轉(zhuǎn)錄因子以及信使分子感受環(huán)境因素的變化從而調(diào)控一系列生理過程并通過產(chǎn)物對代謝途徑的反饋抑制等方式使細(xì)胞適應(yīng)環(huán)境變化,維持內(nèi)穩(wěn)態(tài)。同時,細(xì)胞可以通過轉(zhuǎn)錄、翻譯以及翻譯后修飾等方式調(diào)控特定基因的表達(dá)。核糖核酸開關(guān)是細(xì)胞內(nèi)天然存在的表達(dá)調(diào)控工具,能夠響應(yīng)單磷酸腺苷環(huán)二聚體(c-di-AMP)、單磷酸鳥苷環(huán)二聚體(c-di-GMP)等第二信使進(jìn)而調(diào)控微生物細(xì)胞產(chǎn)芽孢、維持滲透壓等多種生理過程,還能感受細(xì)胞內(nèi)氨基酸的濃度,在濃度較高時通過使轉(zhuǎn)錄提前終止或抑制翻譯起始的方式對相應(yīng)的代謝途徑進(jìn)行反饋抑制[13-15],自2002年首次發(fā)現(xiàn)就引起了轟動。作為調(diào)控基因表達(dá)的RNA元件,核糖核酸開關(guān)由兩部分組成,適體區(qū)(aptamer domain,AD)和基因表達(dá)區(qū)(expression domain,ED),其中AD能識別特異性的小分子代謝物或金屬離子,與之結(jié)合后使ED變構(gòu),從而能夠在轉(zhuǎn)錄水平或翻譯水平上調(diào)控相關(guān)基因的表達(dá)(圖1)[16]。由于核糖核酸開關(guān)不僅能感受信號還能使細(xì)胞針對信號變化做出相應(yīng)的反應(yīng),并且不需要蛋白質(zhì)的參與,具有結(jié)構(gòu)簡單、反應(yīng)迅速等優(yōu)點(diǎn)。自發(fā)現(xiàn)其原理以來,人們通過模擬天然核糖核酸開關(guān)的調(diào)控機(jī)理設(shè)計(jì)出一系列人工核糖核酸開關(guān),并將這些核糖核酸開關(guān)成功應(yīng)用于調(diào)控細(xì)菌的趨化性、構(gòu)建電子基因線路、開發(fā)生物傳感器等方面,近年來也致力于將其作為微生物細(xì)胞工廠的智能化精細(xì)調(diào)控工具而開發(fā)應(yīng)用。因此本文將對核糖核酸開關(guān)的作用機(jī)制及其應(yīng)用前景進(jìn)行綜述。

圖1 核糖核酸開關(guān)的結(jié)構(gòu)Fig.1 Architecture of riboswitch
1 核糖核酸開關(guān)的種類及作用機(jī)制
隨著對核糖核酸開關(guān)研究的逐步深入,現(xiàn)已發(fā)現(xiàn)至少有20種天然的核糖核酸開關(guān)(表1)。這些天然的核糖核酸開關(guān)可以按照配體的不同分為5類,分別是響應(yīng)離子、糖衍生物、氨基酸、核酸衍生物以及響應(yīng)輔酶的核糖核酸開關(guān),下面將對這些天然的核糖核酸開關(guān)的種類及作用機(jī)制進(jìn)行簡要綜述。
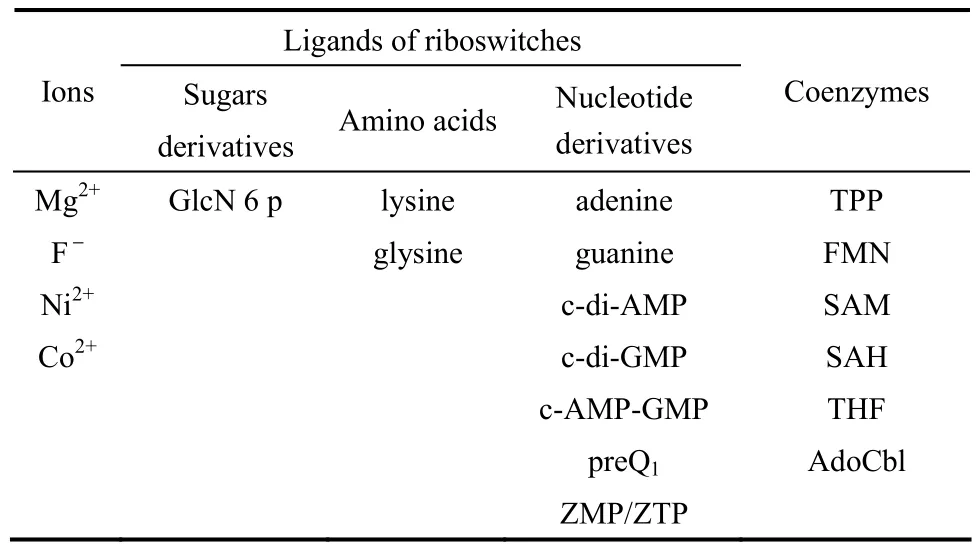
表1 核糖核酸開關(guān)的種類Table 1 Class of riboswitches
1.1 離子配體的核糖核酸開關(guān)
微生物生活在包含各種離子的環(huán)境中,這些離子參與生物體內(nèi)很多重要的催化反應(yīng),如Mg2+是DNA聚合酶的輔酶,如果缺少M(fèi)g2+,DNA聚合酶的效率會顯著降低;但是離子過多則會對細(xì)胞造成毒害,使細(xì)胞死亡。因此維持細(xì)胞內(nèi)的離子內(nèi)穩(wěn)態(tài)至關(guān)重要。早先的研究表明微生物主要通過蛋白質(zhì)來調(diào)控離子轉(zhuǎn)運(yùn)蛋白的表達(dá)從而維持細(xì)胞內(nèi)的離子內(nèi)穩(wěn)態(tài)。近年來,許多研究表明一些位于離子轉(zhuǎn)運(yùn)蛋白5′-UTR的核糖核酸開關(guān)也能感受離子濃度并通過變構(gòu)效應(yīng)調(diào)控離子轉(zhuǎn)運(yùn)蛋白的表達(dá)。
迄今為止,發(fā)現(xiàn)的能識別離子的核糖核酸開關(guān)有Mg2+核糖核酸開關(guān)[17-18]、F?核糖核酸開關(guān)[19]、Ni2+、Co2+核糖核酸開關(guān)[20]。這些識別離子的核糖核酸開關(guān)能夠感受細(xì)胞內(nèi)離子的濃度,高濃度狀態(tài)下核糖核酸開關(guān)打開,使下游編碼的轉(zhuǎn)運(yùn)蛋白的基因表達(dá);低濃度狀態(tài)下核糖核酸開關(guān)關(guān)閉,抑制下游轉(zhuǎn)運(yùn)蛋白的表達(dá)[圖2(a)],從而維持細(xì)胞內(nèi)離子的內(nèi)穩(wěn)態(tài),維持細(xì)胞正常生理過程。
1.2 糖衍生物配體的核糖核酸開關(guān)
目前只發(fā)現(xiàn)一種能識別葡糖衍生物——葡糖胺-6-磷酸(glucosamine-6-phosphate,GlcN6p)的核糖核酸開關(guān),這種核糖核酸開關(guān)被命名為glmS。glmS主要存在于各種革蘭陽性菌中,其中很多都是人類的病原菌。
glmS是現(xiàn)已發(fā)現(xiàn)的天然核糖核酸開關(guān)中一種非常獨(dú)特的核糖核酸開關(guān),其作用機(jī)制不同于一般的調(diào)控轉(zhuǎn)錄的提前終止或者是抑制核糖體與核糖體結(jié)合位點(diǎn)(ribosome binding site,RBS)的結(jié)合。glmS是一種自剪切核酶,其位于基因glmS(編碼谷氨酰胺-果糖-6-磷酸酰胺轉(zhuǎn)移酶,可利用果糖-6-磷酸和谷氨肽合成GlcN6p參與糖代謝和細(xì)胞壁的生物合成)的5′-UTR,通過變構(gòu)效應(yīng)對GlcN6p的合成起反饋抑制作用[21]。當(dāng)細(xì)胞內(nèi)GlcN6p濃度較高時,GlcN6p與glmS結(jié)合,glmS的酶活增強(qiáng),降解谷氨酰胺-果糖-6-磷酸酰胺轉(zhuǎn)移酶的前導(dǎo)mRNA,減少GlcN6p的合成量[圖2(b)]。GlcN6p是細(xì)胞壁成分N-乙酰葡糖胺的前體物質(zhì),GlcN6p量的減少會導(dǎo)致細(xì)胞壁合成障礙,因此可以利用glmS作為開發(fā)新藥的靶位點(diǎn),用來抵抗細(xì)菌的多重耐藥性機(jī)制,研制新型抗生素[22]。

圖2 天然核糖核酸開關(guān)調(diào)控機(jī)制Fig.2 Regulatory mechanism of natural riboswitches
1.3 氨基酸配體的核糖核酸開關(guān)
現(xiàn)已發(fā)現(xiàn)兩種受核糖核酸開關(guān)調(diào)控表達(dá)的氨基酸,分別是甘氨酸和賴氨酸。
甘氨酸核糖核酸開關(guān)的結(jié)構(gòu)非常特別,它是發(fā)現(xiàn)的天然核糖核酸開關(guān)中唯一一個串聯(lián)了兩個識別同一配體的AD和一個ED構(gòu)成的核糖核酸開關(guān)。并且兩個AD表現(xiàn)出與甘氨酸結(jié)合的協(xié)同性,推測這種機(jī)制能夠使核糖核酸開關(guān)對配體的親和性變強(qiáng),提高調(diào)控系統(tǒng)的靈活性[23]。當(dāng)細(xì)胞內(nèi)甘氨酸濃度較高時,甘氨酸與AD結(jié)合,使gcvT基因的5′-UTR區(qū)原本存在的前終止子結(jié)構(gòu)被破壞,轉(zhuǎn)錄出完整的mRNA,使gcvT基因表達(dá)[圖2(c)]。因此,甘氨酸核糖核酸開關(guān)還是一種罕見的與配體結(jié)合后使下游基因表達(dá)的“ON”開關(guān)。
賴氨酸核糖核酸開關(guān)通常位于基因lysC的5′-UTR區(qū),由能識別賴氨酸的AD和包含有RNAaseE識別位點(diǎn)的ED組成。當(dāng)細(xì)胞內(nèi)賴氨酸濃度高時,賴氨酸與AD結(jié)合促使ED變構(gòu)暴露出RNAaseE識別位點(diǎn),從而降解新生的mRNA[圖2(d)],實(shí)現(xiàn)對賴氨酸的反饋抑制[24-25]。
1.4 核酸衍生物配體的核糖核酸開關(guān)
c-di-GMP、c-di-AMP和7-氨甲基-7-去氮雜鳥嘌呤(7-aminomethyl-7-deazaguanine,preQ1)等核酸衍生物均有相應(yīng)的核糖核酸開關(guān),通過與核糖核酸開關(guān)結(jié)合后的變構(gòu)效應(yīng)來調(diào)控基因的表達(dá)。以c-di-GMP核糖核酸開關(guān)為例介紹其作用機(jī)制。
c-di-GMP是細(xì)胞內(nèi)最主要的第二信使分子,能調(diào)控多種重要的生理過程,如調(diào)控病原菌的毒力、鞭毛的運(yùn)動以及生物膜的形成等[26]。2008年耶魯大學(xué)的Breaker課題組首次通過比較基因組學(xué)發(fā)現(xiàn)多種微生物基因組中普遍存在保守的GEMM motif,GEMM motif能識別c-di-GMP并能影響相關(guān)下游基因的表達(dá)。經(jīng)過實(shí)驗(yàn)驗(yàn)證GEMM是一類能識別c-di-GMP的核糖核酸開關(guān)[27]。研究發(fā)現(xiàn),病原菌梭狀芽胞桿菌(Clostridium difficile)中調(diào)控其毒力基因的5′-UTR區(qū)存在一類c-di-GMP核糖核酸開關(guān),這類核糖核酸開關(guān)是一種自剪切核酶,被Breaker命名為I型自剪切核酶(Group I self-splicing ribozyme)。這類核酶通過與c-di-GMP的結(jié)合暴露或者覆蓋住其自身的切割位點(diǎn),從而阻止或者開始下游基因的表達(dá)[28]。
1.5 輔酶配體的核糖核酸開關(guān)
硫胺素焦磷酸(thiamine pyrophosphate,TPP)、黃素單核苷酸(flavin monoucleotide , FMN)、腺苷蛋氨酸(S-adenosyl methionine,SAM)等都是重要的輔酶,分別參與細(xì)胞內(nèi)的脫羧反應(yīng)、氧化還原反應(yīng)以及甲基轉(zhuǎn)移過程,對于維持細(xì)胞的內(nèi)穩(wěn)態(tài)至關(guān)重要。通過比較基因組學(xué)分析發(fā)現(xiàn),多種細(xì)菌基因組中普遍存在保守的響應(yīng)這些輔酶的核糖核酸開關(guān),與相應(yīng)的輔酶結(jié)合后發(fā)生變構(gòu)從轉(zhuǎn)錄水平或翻譯水平調(diào)控相關(guān)基因的表達(dá)。基于這些輔酶的重要生理作用和獨(dú)特的調(diào)控機(jī)制,可以認(rèn)為這些核糖核酸開關(guān)是RNA世界的遺跡。
2 人工核糖核酸開關(guān)的設(shè)計(jì)
天然的核糖核酸開關(guān)可以同時作為生物感受器和效應(yīng)器——感受細(xì)胞內(nèi)小分子代謝物或金屬離子的濃度并根據(jù)相應(yīng)的濃度調(diào)控相關(guān)基因的表達(dá)從而使細(xì)胞維持內(nèi)穩(wěn)態(tài)以適應(yīng)周圍的環(huán)境。核糖核酸開關(guān)的這種特性,可用于開發(fā)智能化的精細(xì)調(diào)控工具,優(yōu)化細(xì)胞工廠的調(diào)控網(wǎng)絡(luò)。然而天然核糖核酸開關(guān)的配體多為細(xì)胞的初級代謝產(chǎn)物,存在調(diào)控的背景干擾過強(qiáng)的問題,因此設(shè)計(jì)識別細(xì)胞內(nèi)原本不存在的配體的人工核糖核酸開關(guān)將更有利于實(shí)現(xiàn)細(xì)胞工廠的智能化調(diào)控。
2.1 基于控制轉(zhuǎn)錄提前終止原理設(shè)計(jì)核糖核酸開關(guān)
某些基因的5′-UTR存在一些不依賴于Rho的終止子結(jié)構(gòu),天然的核糖核酸開關(guān)AD與配體結(jié)合后會使ED變構(gòu)導(dǎo)致ED的部分序列與終止子序列形成W-C配對,從而破壞5′-UTR天然的終止子結(jié)構(gòu),使基因能夠轉(zhuǎn)錄。基于這一原理,M?rl課題組設(shè)計(jì)了一系列在轉(zhuǎn)錄水平調(diào)控基因表達(dá)的人工核糖核酸開關(guān)[29]。他們的設(shè)計(jì)策略是在茶堿適體的3′端和終止子結(jié)構(gòu)的聚尿嘧啶尾巴之間隨機(jī)插入不同長度的核苷酸,利用軟件RNA fold模擬插入序列能否與聚尿嘧啶尾巴形成類似終止子的莖環(huán)結(jié)構(gòu),再通過報(bào)告基因的顏色變化進(jìn)行篩選,最后得到一個人工設(shè)計(jì)的能阻止轉(zhuǎn)錄提前終止的核糖核酸開關(guān)。當(dāng)沒有茶堿時,ED形成終止子結(jié)構(gòu)轉(zhuǎn)錄不能進(jìn)行,添加茶堿后,茶堿與AD結(jié)合,終止子結(jié)構(gòu)被破壞,轉(zhuǎn)錄開始,目標(biāo)基因表達(dá)[圖3(a)]。
2.2 基于控制RBS與核糖體結(jié)合原理設(shè)計(jì)核糖核酸開關(guān)
通過控制RBS與核糖體的結(jié)合來調(diào)控翻譯的起始,是天然核糖核酸開關(guān)最常用的兩種調(diào)控策略之一,天然的核糖核酸開關(guān)AD與配體結(jié)合后會導(dǎo)致RBS被遮蓋或者暴露,從而抑制或起始目標(biāo)基因的翻譯過程。利用這一機(jī)制,Gallivan等[30]設(shè)計(jì)了一系列人工核糖核酸開關(guān)。其設(shè)計(jì)思想是在茶堿適體和RBS之間隨機(jī)插入不同長度的核苷酸,通過報(bào)告基因cat對四環(huán)素的抗性篩選,分別得到響應(yīng)茶堿后表達(dá)cat基因的“ON”核糖核酸開關(guān)和響應(yīng)茶堿后抑制cat基因表達(dá)的“OFF”核糖核酸開關(guān)[圖3(b)]。
2.3 基于控制mRNA提前降解原理設(shè)計(jì)核糖核酸開關(guān)
有些天然核糖核酸開關(guān)本身是自剪切核酶,如GlcN6p核糖核酸開關(guān),與配體結(jié)合后暴露出酶切位點(diǎn),降解目標(biāo)基因的mRNA。基于這一原理,耶魯大學(xué)的Breaker課題組將一個ATP適體與錘頭狀核酶通過一段隨機(jī)序列的交流模塊連接起來,通過外加ATP,觀察錘頭狀核酶能否自剪切,最終得到兩種具有自剪切核酶功能的人工核糖核酸開關(guān),一種加入ATP后使核酶自剪切,一種加入ATP后能夠抑制核酶的自剪切[31][圖3(c)]。
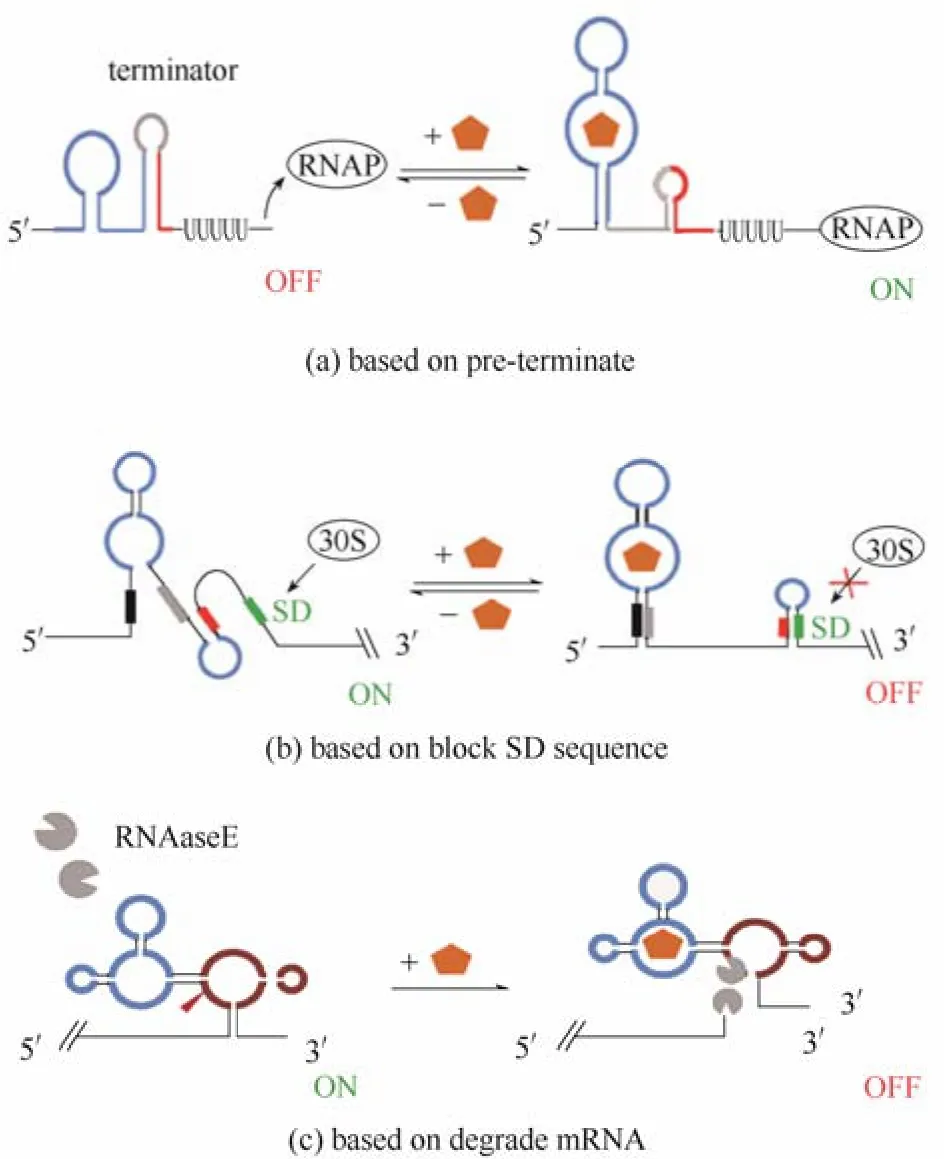
圖3 人工核糖核酸開關(guān)設(shè)計(jì)原理Fig.3 Principle of design artificial riboswitch
3 核糖核酸開關(guān)在細(xì)胞工廠中的應(yīng)用
3.1 應(yīng)用于工業(yè)菌株的篩選和途徑的精細(xì)調(diào)控
性狀優(yōu)良的菌株是發(fā)酵工業(yè)的靈魂,傳統(tǒng)方法往往是對出發(fā)菌株進(jìn)行物理或化學(xué)誘變,再通過對突變文庫進(jìn)行人工篩選來檢驗(yàn)有沒有理想的高產(chǎn)菌株。但這種方法對環(huán)境和實(shí)驗(yàn)人員的傷害較大,而且誘變不能針對某一特定性狀,基因的隨機(jī)突變導(dǎo)致突變文庫較大,很難篩選到理想的工業(yè)菌株。天然核糖核酸開關(guān)的效應(yīng)物大多是一些初級代謝產(chǎn)物,這些核糖核酸開關(guān)可以特異性地針對某種代謝產(chǎn)物在細(xì)胞內(nèi)行使反饋調(diào)節(jié)的作用。因此可以利用核糖核酸開關(guān)對代謝途徑進(jìn)行精細(xì)調(diào)控,提高目標(biāo)產(chǎn)物產(chǎn)量。例如Zeng課題組在生產(chǎn)賴氨酸的工業(yè)菌株谷氨酸棒狀桿菌LP917(Corynebacterium glutamicumLP917)基因gltA(編碼檸檬酸合酶)的5′-UTR區(qū)插入響應(yīng)賴氨酸的核糖核酸開關(guān),通過外加賴氨酸抑制檸檬酸合酶的轉(zhuǎn)錄,使菌體積累草酰乙酸。草酰乙酸作為合成賴氨酸的前體物質(zhì),它的積累能提高賴氨酸的產(chǎn)量,進(jìn)一步抑制檸檬酸合酶的產(chǎn)生,形成一個正循環(huán),提高工業(yè)菌株中賴氨酸的產(chǎn)量[32]。利用這種方法,他們成功使賴氨酸的產(chǎn)量提高了63%。
Jung課題組[33]采用另一種策略構(gòu)建了包含賴氨酸核糖核酸開關(guān)和tetA基因[34](編碼四環(huán)素/H+轉(zhuǎn)運(yùn)蛋白,少量表達(dá)能夠抗四環(huán)素,大量表達(dá)會使細(xì)菌對Ni2+類金屬離子敏感)的篩選系統(tǒng)。在培養(yǎng)基中添加NiCl2作為篩選壓力,菌體內(nèi)賴氨酸濃度高,抑制tetA基因的表達(dá),使菌體對NiCl2不敏感,能夠存活;而當(dāng)菌體內(nèi)賴氨酸濃度低,tetA基因表達(dá),菌體對NiCl2敏感,死亡。因此經(jīng)過幾輪的篩選和富集,最終得到的就是高產(chǎn)賴氨酸的菌株。Jung課題組構(gòu)建的這種基于核糖核酸開關(guān)來篩選高產(chǎn)菌株的平臺,只要能找到響應(yīng)效應(yīng)物的核糖核酸開關(guān)就能定向地篩選到高產(chǎn)菌株,具有針對性強(qiáng)、適用范圍廣、操作簡單等優(yōu)點(diǎn)。
此外,工業(yè)發(fā)酵過程會產(chǎn)生大量的發(fā)酵熱,不僅影響微生物的生長和生產(chǎn),還因冷卻控溫增加了生產(chǎn)成本,因此發(fā)酵工業(yè)迫切需要耐熱的底盤宿主。基于溫度變構(gòu)原理,人工設(shè)計(jì)響應(yīng)不同溫度的核糖核酸開關(guān)[35],使熱激蛋白(heat shock protein,HSP)、超氧化物歧化酶(superoxide dismutase, SOD)等能提高微生物耐熱性的耐熱元件在不同溫度下依次表達(dá),不僅能提高微生物的耐熱性還能減少能耗,保護(hù)環(huán)境。
綜上所述,利用核糖核酸開關(guān)來構(gòu)建的工業(yè)菌株篩選方法具有特異性強(qiáng)、靈活度高、對環(huán)境和實(shí)驗(yàn)人員危害小等優(yōu)點(diǎn),是一種簡便高效的篩選工業(yè)菌株的方法,在發(fā)酵工業(yè)上有巨大的應(yīng)用價值。
3.2 應(yīng)用于新藥開發(fā)
隨著抗生素類藥物的廣泛使用,細(xì)菌的多耐藥現(xiàn)象引起了人們的關(guān)注,近30年來研發(fā)的新型抗生素藥物屈指可數(shù),甚至出現(xiàn)了“抗生素悖論”效應(yīng)。傳統(tǒng)抗生素的作用機(jī)制僅限于阻礙核酸的合成、干擾蛋白質(zhì)的合成、影響細(xì)胞膜和細(xì)胞壁的功能,因此亟需找到新型藥物作用靶點(diǎn),利用特異性的調(diào)控作用來抑制致病基因的表達(dá)。核糖核酸開關(guān)在細(xì)菌中普遍存在并參與細(xì)菌基本生命代謝的重要過程,將核糖核酸開關(guān)作為研發(fā)新藥的作用靶點(diǎn)[36],大大拓展了傳統(tǒng)抗生素的作用機(jī)制,為解決多重耐藥性問題提供了嶄新的思路。
氟化物在ATP或GTP結(jié)合二價金屬離子的過程中能模擬ATP或GTP的γ磷酸基團(tuán)[37],從而阻斷很多酶的活性[38],對各種有機(jī)體都有毒性。病原菌中存在天然的響應(yīng)氟離子的核糖核酸開關(guān),當(dāng)細(xì)胞內(nèi)氟離子過多時,開關(guān)打開,表達(dá)氟離子轉(zhuǎn)運(yùn)蛋白,外排出過多的氟化物從而維持細(xì)胞內(nèi)氟離子的內(nèi)穩(wěn)態(tài)。開發(fā)新藥使氟離子更多地進(jìn)入細(xì)胞,打破病原菌內(nèi)的氟離子內(nèi)穩(wěn)態(tài),則能有效殺死病原菌。基于這一思想,Breaker課題組在病原菌中構(gòu)建了一個高效篩選系統(tǒng),這個系統(tǒng)包含響應(yīng)氟離子的核糖核酸開關(guān)和報(bào)告基因lacZ。將含有高效篩選系統(tǒng)的病原菌涂在含有X-gal和氟化物以及不同藥物的培養(yǎng)基上,通過顏色的變化判斷哪種新藥能使更多的氟離子進(jìn)入細(xì)胞,通過一系列實(shí)驗(yàn)發(fā)現(xiàn)短桿菌肽作用于細(xì)胞膜上的氟離子通道,使更多的氟離子進(jìn)入細(xì)胞,從而殺死病原菌[39]。通過短桿菌肽輔助氟離子殺菌的方式能有效抑制口腔病原菌的生長,對治療齲齒有良好的效果。
glmS核糖核酸開關(guān)與配體GlcN6p結(jié)合后抑制谷氨酰胺-果糖-6-磷酸酰胺轉(zhuǎn)移酶的表達(dá),導(dǎo)致產(chǎn)物GlcN6p的量減少,而GlcN6p是細(xì)菌細(xì)胞壁N-乙酰葡糖胺的前體,減少GlcN6p將抑制細(xì)菌細(xì)胞壁的合成。利用這一思路,Soukup課題組設(shè)計(jì)了一系列GlcN6p的結(jié)構(gòu)類似物,其中有兩個能有效與glmS核糖核酸開關(guān)結(jié)合發(fā)揮抑菌作用[40]。這兩種GlcN6p的結(jié)構(gòu)類似物可以作為新藥作用于含有g(shù)lmS核糖核酸開關(guān)的多種病原菌。
Gallivan課題組在大腸桿菌中構(gòu)建了一個調(diào)控大腸桿菌趨化性[41]的系統(tǒng),這個系統(tǒng)由茶堿核糖核酸開關(guān)和調(diào)控鞭毛旋轉(zhuǎn)的cheZ基因組成。通過外源添加茶堿,使CheZ表達(dá),控制細(xì)菌向茶堿方向移動[42]。利用這種改變細(xì)菌趨化性的策略,可以使攜帶藥物工程菌向病灶聚集增強(qiáng)治病能力。
3.3 作為酶分子定向進(jìn)化的高通量篩選方法
細(xì)胞工廠通過酶的催化作用生產(chǎn)各種滿足人類需要的化合物,可以說酶的催化作用是工業(yè)發(fā)酵的核心。然而,許多天然酶具有與底物識別的專一性差、催化效率低以及熱穩(wěn)定性差等問題,需要對其進(jìn)行改性才能滿足工業(yè)生產(chǎn)的需求。
定向進(jìn)化是一種有效的酶改性策略,可以通過易錯PCR等方法對編碼基因進(jìn)行隨機(jī)突變或重組,使其原有活性提高或者產(chǎn)生新的功能。由于隨機(jī)突變后產(chǎn)生的文庫較大,篩選難度高,定向進(jìn)化成功的關(guān)鍵在于能否針對目標(biāo)特性開發(fā)出高效的篩選方法。核糖核酸開關(guān)在微生物中普遍存在并且能響應(yīng)特定的效應(yīng)物調(diào)節(jié)目標(biāo)基因的表達(dá),因此可作為一個有效的篩選模塊,為開發(fā)高效篩選方法提供嶄新的思路。Smolke課題組將響應(yīng)茶堿的適體與自剪切核酶融合后構(gòu)建了能響應(yīng)茶堿的核酶開關(guān),當(dāng)沒有茶堿時該核糖核酸開關(guān)結(jié)構(gòu)不穩(wěn)定會降解目標(biāo)基因的mRNA,抑制目標(biāo)基因的表達(dá);只有當(dāng)茶堿與適體結(jié)合后核酶結(jié)構(gòu)穩(wěn)定,才能表達(dá)目標(biāo)基因。將這個人工設(shè)計(jì)的核糖核酸開關(guān)構(gòu)建到咖啡因脫甲基酶與綠色熒光蛋白(green fluorescent protein,GFP)融合的蛋白質(zhì)的3′-UTR區(qū),可以用于咖啡因脫甲基酶的定向進(jìn)化。因?yàn)榭Х纫蛎摷谆改軌驅(qū)⒖Х纫蜣D(zhuǎn)化成茶堿,而茶堿是該核糖核酸開關(guān)的配體,通過熒光強(qiáng)度的不同利用流式細(xì)胞儀(fluorescence activated cell sorter,F(xiàn)ACS)篩選到酶活提高了32倍的咖啡因脫甲基酶[43]。
3.4 構(gòu)建新型生物傳感器
生物傳感器是將生物識別元件和信號轉(zhuǎn)換元件緊密結(jié)合,從而檢測目標(biāo)化合物的分析裝置。將核糖核酸開關(guān)與報(bào)告基因(beta-半乳糖苷酶、熒光蛋白、熒光素酶)串聯(lián)在一起,通過報(bào)告基因的表達(dá)強(qiáng)度來判斷相應(yīng)的配體濃度,開發(fā)出各種新型生物傳感器。這些新型生物傳感器與傳統(tǒng)的生物傳感器相比,不需要借助細(xì)胞外的各種光學(xué)元件或者壓電裝置,可以直接將細(xì)胞內(nèi)部某一代謝物的濃度信號轉(zhuǎn)化為顏色變化等輸出信號。具有結(jié)構(gòu)簡單、反應(yīng)靈敏、價格低廉、生物安全性高等優(yōu)點(diǎn),可以廣泛應(yīng)用于食品藥品的檢測等方面。如Gu等[44]將天然的響應(yīng)腺苷鈷胺素(adenosyl-cobalamin, AdoCbl)的核糖核酸開關(guān)構(gòu)建到GFP的5′-UTR區(qū),通過熒光強(qiáng)度的變化來檢測B12的濃度,利用這種方法對AdoCbl的檢測限達(dá)到10 ng·ml?1,相當(dāng)于高效液相色譜(high performance liquid chromatography,HPLC)的水平。
融合適體與核酶構(gòu)建成的人工適體酶(aptazyme)能將與靶分子結(jié)合的信號穩(wěn)定轉(zhuǎn)換成更清晰的酶學(xué)信號,如自切割活性,通過在適體酶上標(biāo)記放射性核素或熒光基團(tuán),能夠檢測到微量濃度的配體[45]。
2011年Jaffrey等[46]利用RNA成功模擬了綠色熒光蛋白,設(shè)計(jì)出一種名為Spinach的核糖核酸開關(guān),結(jié)合底物DMHBI之后會發(fā)出綠色熒光。通過將Spinach與相應(yīng)不同的適體融合,開發(fā)出能檢測c-AMP- GMP[47]、腺苷酸、ADP、SAM、谷氨酰胺和GFP[48]的核糖核酸開關(guān)。綜上所述基于核糖核酸開關(guān)開發(fā)的生物傳感器是一種檢測發(fā)酵工業(yè)中某種特定產(chǎn)物的有效手段。
4 結(jié)論與展望
自2002年Breaker首次提出核糖核酸開關(guān)這個術(shù)語[49],對于核糖核酸開關(guān)的研究已經(jīng)整整13年,工程化的核糖核酸開關(guān)已經(jīng)不再僅僅是一個構(gòu)想,而被用于在各種工業(yè)微生物中構(gòu)建具有魯棒性和正交性的調(diào)控系統(tǒng),利用不同的調(diào)控機(jī)制對細(xì)胞工廠進(jìn)行智能調(diào)控。目前,工程化的核糖核酸開關(guān)已經(jīng)成功應(yīng)用于篩選工業(yè)菌株、研發(fā)新藥、構(gòu)建酶分子的定向進(jìn)化的篩選方法以及開發(fā)新型生物傳感器等方面。
與基因表達(dá)的蛋白調(diào)控系統(tǒng)比較,核糖核酸開關(guān)調(diào)控基因的表達(dá)不需要蛋白質(zhì)的參與,具有免疫原性小、結(jié)構(gòu)簡單、反應(yīng)迅速等優(yōu)勢。然而天然核糖核酸開關(guān)響應(yīng)的效應(yīng)物大多是細(xì)胞內(nèi)的初級代謝產(chǎn)物,利用天然核糖核酸開關(guān)開發(fā)的細(xì)胞內(nèi)調(diào)控系統(tǒng)存在背景干擾過強(qiáng)的問題。針對這一現(xiàn)象,可以以細(xì)胞內(nèi)不存在的物質(zhì)作為配體設(shè)計(jì)人工核糖核酸開關(guān),能有效區(qū)分核糖核酸開關(guān)調(diào)控的基因是受細(xì)胞內(nèi)源配體影響還是外加配體影響,解決細(xì)胞內(nèi)調(diào)控背景干擾過強(qiáng)的問題。除此之外,核糖核酸開關(guān)的工業(yè)化應(yīng)用還存在一個問題——難以篩選到合適的響應(yīng)非細(xì)胞內(nèi)天然產(chǎn)物的適體。雖然理論上任何一種物質(zhì)都能在體外篩選到相應(yīng)的適體,然而大量實(shí)驗(yàn)證明體外篩選到的適體大多不能在體內(nèi)起作用,這可能是因?yàn)轶w內(nèi)的環(huán)境與體外不同所導(dǎo)致的。而這一問題大大限制了核糖核酸開關(guān)的應(yīng)用,因此,如何開發(fā)出更多的能在體內(nèi)響應(yīng)不同配體的適體是核糖核酸開關(guān)在工業(yè)化應(yīng)用進(jìn)程中的一大挑戰(zhàn)。
符 號 說 明
AD——核糖核酸開關(guān)的適體區(qū)
AdoCbl——腺苷鈷胺素
c-di-AMP——單磷酸腺苷環(huán)二聚體
c-di-GMP——單磷酸鳥苷環(huán)二聚體
FACS——流式細(xì)胞儀
FMN——黃素單核苷酸
GFP——綠色熒光蛋白
GlcN6p——葡糖胺-6-磷酸
HPLC——高效液相色譜
HSP——熱激蛋白
ORF——開放閱讀框
PHA——聚羥基脂肪酸酯
PHB——聚羥基丁酸酯
preQ1——7-氨甲基-7-去氮雜鳥嘌呤
RBS——核糖體結(jié)合位點(diǎn)
SAM——腺苷蛋氨酸
SOD——超氧化物歧化酶
TPP——硫胺素焦磷酸
UTR——非翻譯區(qū)
[1] Lai Shujuan, Zhang Yun, Liu Shuwen, Liang Yong, Shang Xiuling, Chai Xin, Wen Tingyi. Metabolic engineering and flux analysis ofCorynebacterium glutamicumfor L-serine production [J].Science China Life Sciences, 2012, 55(4): 283-290.
[2] Wu Y, Li P, Zheng P, Zhou W, Chen N, Sun J. Complete genome sequence ofCorynebacterium glutamicumB253, a Chinese lysine-producing strain [J].J. Biotechnol., 2015, 4(207): 10-11.
[3] Wang N, Ni Y, Shi F. Deletion of odhA or pyc improves production of γ-aminobutyric acid and its precursor L-glutamate in recombinantCorynebacterium glutamicum[J].Biotechnol. Lett., 2015, 37(7): 1473-1481.
[4] Shi X, Chen Y, Ren H, Liu D, Zhao T, Zhao N, Ying H. Economically enhanced succinic acid fermentation from cassava bagasse hydrolysate usingCorynebacterium glutamicumimmobilized in porous polyurethane filler [J].Bioresour. Technol., 2014, 174: 190-197.
[5] Dae-Kyun Ro, Eric M Paradise, Jay D Keasling,et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast [J].Nature, 2005, 440(7086): 940-943.
[6] Patrick J Westfall, Douglas J Pitera, Jay D Keasling,et al. Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin [J].Proc. Natl. Acad. Sci. USA, 2012, 109(3): E111-E118.
[7] Jennewein S, Park H, DeJong J M, Long R M, Bollon A P, Croteau R B. Coexpression in yeast of Taxus cytochrome P450 reductase with cytochrome P450 oxygenases involved in Taxol biosynthesis [J].Biotechnol. Bioeng., 2005, 89(5): 588-598.
[8] Guo J, Zhou Y J, Hillwig M L, Huang L,et al. CYP76AH1 catalyzes turnover of miltiradiene in tanshinones biosynthesis and enables heterologous production of ferruginol in yeasts [J].Proc. Natl. Acad.Sci.USA, 2013, 110(29): 12108-12113.
[9] Gao Z X, Zhao H, Li Z M, Tan X M, Lu X F. Photosynthetic production of ethanol from carbon dioxide in genetically engineered cyanobacteria [J].Energy & Environmental Science,2012, 5(12): 9857-9865.
[10] Lan E I, Liao J C. Metabolic engineering of cyanobacteria for 1-butanol production from carbon dioxide [J].Metabolic Engineering, 2011, 13(4): 353-363.
[11] Meng D C, Wang Y, Wu L P, Shen R, Chen J C, Wu Q, Chen G Q. Production of poly (3-hydroxypropionate) and poly (3-hydroxybutyrate-co-3-hydroxypropionate) from glucose by engineeringEscherichia coli[J].Metab. Eng., 2015, 29: 189-195.
[12] Jari M, Khatami S R, Galehdari H, Shafiei M. Cloning and expression of poly 3-hydroxybutyrate operon intoEscherichia coli[J].Jundishapur Journal of Microbiology, 2015, 8(2): e16318.
[13] Nahvi A, Sudarsan N, Ebert M S, Zou X, Brown K L, Breaker R R. Genetic control by a metabolite binding mRNA [J].ChemBiol., 2002, 9(9): 1043.
[14] Winkler W C, Cohen-Chalamish S, Breaker R R. An mRNA structure that controls gene expression by binding FMN [J].Proc. Natl. Acad. Sci. USA, 2002, 99: 15908-15913.
[15] Mironov A S, Gusarov I, Rafikov R, Lopez L E, Shatalin K, Kreneva R A, Perumov D A, Nudler E. Sensing small molecules by nascent RNA: a mechanism to control transcription in bacteria [J].Cell, 2002, 111: 747-756.
[16] Breaker R R. Prospects for riboswitch discovery and analysis [J].Molecular Cell, 2011, 43(6): 867-879.
[17] Dann C E 3rd, Wakeman C A, Sieling C L, Baker S C, Irnov I, Winkler W C. Structure and mechanism of a metal-sensing regulatory RNA [J].Cell, 2007, 130(5): 878-892.
[18] Ramesh A, Winkler W C. Magnesium-sensing riboswitches in bacteria [J].Journal of RNA Biol., 2010, 7: 77-83.
[19] Chawla M, Credendino R, Poater A, Oliva R, Cavallo L. Structural stability, acidity, and halide selectivity of the fluoride riboswitch recognition site [J].J. Am. Chem. Soc.,2015, 137(1): 299-306.
[20] Furukawa K, Ramesh A, Zhou Z Y, Weinberg Z, Vallery T, Wade W C, Breaker R R. Bacterial riboswitches cooperatively bind Ni2+or Co2+ions and control expression of heavy metal transporters [J].Molecular Cell, 2015, 57(6): 1088-1098.
[21] McCown P J, Winkler W C, Breaker R R. Mechanism and distribution of glmS ribozymes [J].Methods. Mol. Biol., 2012, 848: 113-129.
[22] Soukup J K. The structural and functional uniqueness of the glmS ribozyme [J].Catalytic Rna., 2013, 120: 173-193.
[23] Mandal M, Lee M, Barrick J E, Weinberg Z, Emilsson G M, Ruzzo W L, Breaker R R. A glycine-dependent riboswitch that uses cooperative binding to control gene expression. [J].Science, 2004, 306(5694): 275-279.
[24] Mellin J R, Pascale Cossart. Unexpected versatility in bacterial riboswitches [J].Trends in Genetics, 2015, 31(3): 150-156.
[25] Wilson-Mitchell S N, Grundy F J, Henkin T M. Analysis of lysine recognition and specificity of theBacillus subtilisL box riboswitch [J].Nucleic Acids Res., 2012, 40(12): 5706-5717.
[26] Furukawa K, Gu H, Sudarsan N, Hayakawa Y, Hyodo M, Breaker R R. Identification of ligand analogues that control c-di-GMP riboswitches [J].ACS Chem. Biol., 2012, 7(8): 1436-1443.
[27] Sudarsan N, Lee E R, Weinberg Z, Moy R H, Kim J N, Link K H, Breaker R R. Riboswitches in eubacteria sense the second messenger cyclic di-GMP [J].Science, 2008, 321: 411-413.
[28] Lee E R, Baker J L, Weinberg Z, Sudarsan N, Breaker R. An allosteric self-splicing ribozyme triggered by a bacterial second messenger [J].Science, 2010, 329(5993): 845-848.
[29] Wachsmuth M, Findeiss S, Weissheimer N, Stadler P F, M?rl M. Denovodesign of a synthetic riboswitch that regulates transcription termination [J].Nucleic Acids Res., 2013, 41(4): 2541-2551.
[30] Desai S K, Gallivan J P. Genetic screens and selections for small molecules based on a synthetic riboswitch that activates protein translation [J].Journal of the American Chemical Society, 2004, 126(41): 13247-13254.
[31] Tang J, Breaker R R. Rational design of allosteric ribozymes [J].Chemistry & Biology, 1997, 4(6): 453-459.
[32] Zhou L B, Zeng A P. Exploring lysine riboswitch for metabolic flux control and improvement of L-lysine synthesis inCorynebacterium glutamicum[J].ACS Synth. Biol., 2015, 4(6): 729-734.
[33] Yang J, Seo S W, Jang S, Shin S I, Lim C H, Roh T Y, Jung G Y. Synthetic RNA devices to expedite the evolution of metabolite-producing microbes [J].Nat. Commun., 2013, 4: 1413.
[34] Muranaka N, Sharma V, Nomura Y, Yokobayashi Y. An efficient platform for genetic selection and screening of gene switches inEscherichia coli[J].Nucleic Acids Res., 2009, 37(5): e39.
[35] Cimdins A, Klinkert B, Aschke-Sonnenborn U, Kaiser F M, Kortmann J, Narberhaus F. Translational control of small heat shock genes in mesophilic andThermophilic cyanobacteriaby RNA thermometers [J].RNA Biol., 2014, 11(5): 594-608.
[36] Deigan K E, Ferré-D'Amaré A R. Riboswitches: discovery of drugs that target bacterial gene-regulatory RNAs [J].Accounts of Chemical Research, 2011, 44(12): 1329-1338.
[37] Li L. The biochemistry and physiology of metallic fluoride: action, mechanism, and implications [J].Critical Reviews in Oral Biology & Medicine, 2003, 14(2): 100-114.
[38] Barbier O, Arreola-Mendoza L, Del Razo L M. Molecular mechanisms of fluoride toxicity [J].Chem. Biol. Interact., 2010, 188(2): 319-333.
[39] Nelson J W, Zhou Z, Breaker R R. Gramicidin D enhances the antibacterial activity of fluoride [J].Bioorg. Med. Chem. Lett., 2014, 24(13): 2969-2971.
[40] Fei X, Holmes T, Diddle J, Hintz L, Delaney D, Stock A, Renner D, McDevitt M, Berkowitz D B, Soukup J K. Phosphatase-inert glucosamine 6-phosphate mimics serve as actuators of the glmS riboswitch [J].ACS Chem. Biol., 2014, 9(12): 2875-2882.
[41] Bren A, Eisenbach M. How signals are heard during bacterial chemo taxis: protein-protein interactions in sensory signal propagation [J].J. Bacteriol.,2000, 182(24): 6865-6873.
[42] Topp S, Gallivan J P. Guiding bacteria with small molecules and RNA [J].J. Am. Chem. Soc., 2007, 129(21): 6807-6811.
[43] Joshua K Michener, Christina D Smolke. High-throughput enzyme evolution inSaccharomyces cerevisiaeusing a synthetic RNA switch [J].Metabolic Engineering,2012, 14(4): 306-316.
[44] Zhu X, Wang X, Zhang C, Wang X, Gu Q. A riboswitch sensor to determine vitamin B12in fermented foods [J].Journal of Food Chem.,2015, 15(175): 523-528.
[45] Knudsen S M, Lee J, Ellington A D, Savran C A. Ribozyme-mediated signal augmentation on a aass-sensitive biosenser [J].J. Am. Chem.Soc., 2006, 128(50): 15936-15937.
[46] Paige J S, Wu K Y, Jaffrey S R. RNA mimics of green fluorescent protein [J].Science,2011, 333(6042): 642-646.
[47] Kellenberger C A, Wilson S C, Sales-Lee J, Hammond M C. RNA-based fluorescent biosensors for live cell imaging of second messengers cyclic di-GMP and cyclic AMP-GMP [J].J. Am. Chem. Soc.,2013, 135(13): 4906-4909.
[48] Strack R L, Jaffrey S R. New approaches for sensing metabolites and proteins in live cells using RNA [J].Curr. Opin. Chem. Biol., 2013, 17(4): 651-655.
[49] Nahvi A, Sudarsan N, Ebert M S, Zou X, Brown K L, Breaker R R. Genetic control by a metabolite binding mRNA [J].Chem. Biol., 2002, 9(9): 1043-1049.
Intelligent and fine regulation of microbial cell factory based on riboswitches
ZHAO Yujia,ZHANG Genlin,ZHOU Xiaohong, LI Chun
(Department of Bioengineering,College of Life Science,Beijing Institute of Technology,Beijing100081,China)
Construction of the microbial cell factory is one of the developmental directions of current green chemical industry. The microbial cell factory is a kind of recombined microorganism and its metabolic and regulatory pathways have been reconstructed by metabolic engineering and synthetic biology to synthetic new compounds or to improve the yield of target production. The microbial metabolic pathway is regulated by two points: environment and genetic information. The cell maintains its homeostasis by global transcription factors, messenger molecules and feedback inhibition when the circumstance is changed. Meanwhile, the cell is affected by its own genetic circulate through transcription, translation and post-translational modification to regulate the expression of target gene. The riboswitches are RNA elements which change their conformation when bind to specific ligands such as ions, sugar derivatives, amino acids, nucleic acid derivatives and coenzymes to regulate the process of transcription, translation and splicing of mRNA. The riboswitches are natural biosensors and bioeffectors which can be designed as the intelligent molecular tools to fine regulate microbial cell factories. Using riboswitches in the microbial cell factory can extend the application in the field of chemical, pharmaceutical, environmental protection and food production.
Prof. LI Chun, lichun@bit.edu.cn
10.11949/j.issn.0438-1157.20150935
TQ 936.2
:A
:0438—1157(2015)10—3811—09
2015-06-16收到初稿,2015-07-16收到修改稿。
聯(lián)系人:李春。
:趙雨佳(1989—),女,博士研究生。
國家杰出青年科學(xué)基金項(xiàng)目(21425624);國家自然科學(xué)基金面上項(xiàng)目(21376028,21476026)。
Received date: 2015-06-16.
Foundation item: supported by the National Science Fund for Distinguished Young Scholars(21425624)and the National Natural Science Foundation of China(21376028, 21476026).