鹽度對(duì)花鰻鱺和太平洋雙色鰻鱺幼鰻生長(zhǎng)性能及消化酶活力的影響
2015-02-28 08:28:18羅鳴鐘關(guān)瑞章靳
水生生物學(xué)報(bào) 2015年4期
羅鳴鐘關(guān)瑞章靳 恒
(1. 長(zhǎng)江大學(xué)動(dòng)物科學(xué)學(xué)院, 荊州 434025; 2. 集美大學(xué), 廈門(mén) 361021; 3. 鰻鱺現(xiàn)代產(chǎn)業(yè)技術(shù)教育部工程研究中心,廈門(mén) 361021)
鹽度對(duì)花鰻鱺和太平洋雙色鰻鱺幼鰻生長(zhǎng)性能及消化酶活力的影響
羅鳴鐘1關(guān)瑞章2, 3靳 恒1
(1. 長(zhǎng)江大學(xué)動(dòng)物科學(xué)學(xué)院, 荊州 434025; 2. 集美大學(xué), 廈門(mén) 361021; 3. 鰻鱺現(xiàn)代產(chǎn)業(yè)技術(shù)教育部工程研究中心,廈門(mén) 361021)
文章研究了不同鹽度對(duì)花鰻鱺(Anguilla marmorata)幼鰻和太平洋雙色鰻鱺(A. bicolor pacifica)幼鰻生長(zhǎng)性能及消化酶活力的影響。將花鰻鱺幼鰻[(9.76±0.36) g]和太平洋雙色鰻鱺幼鰻[(11.82±0.04) g]分別在淡水(鹽度0‰)與鹽度5‰、10‰、18‰水體中養(yǎng)殖30d, 測(cè)量每組實(shí)驗(yàn)魚(yú)總重后檢測(cè)胃、腸道和肝臟蛋白酶、淀粉酶和脂肪酶的活力。結(jié)果表明, 花鰻鱺和太平洋雙色鰻鱺在各鹽度處理中存活率均為100%, 未出現(xiàn)死亡。兩種鰻鱺在淡水中生長(zhǎng)良好, 特定生長(zhǎng)率最高, 而餌料系數(shù)最低。鹽度對(duì)花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻消化酶活力的影響存在差異, 其中花鰻鱺胃、腸道和肝臟蛋白酶活力在各鹽度處理中均無(wú)顯著變化(P>0.05),淀粉酶和脂肪酶活力均隨鹽度的增加而下降; 太平洋雙色鰻鱺胃蛋白酶活力在鹽度 10‰時(shí)最大, 肝蛋白酶活力在鹽度18‰時(shí)最大, 而淀粉酶和脂肪酶活力在各鹽度處理組無(wú)顯著變化(P>0.05)。這表明鹽度對(duì)花鰻鱺胃、腸道和肝臟的淀粉酶和脂肪酶活力具有抑制作用, 對(duì)太平洋雙色鰻鱺的蛋白酶活力有一定的激活作用。在相同鹽度條件下, 不同消化器官中同種消化酶活力存在差異, 各鹽度的兩種鰻鱺腸道中淀粉酶和脂肪酶的活力均顯著高于肝臟和胃(P<0.05), 胃中蛋白酶活力高于肝臟和腸道, 但不顯著(P>0.05)。研究發(fā)現(xiàn)兩種鰻鱺體內(nèi)脂肪酶活力相對(duì)較高, 表明其對(duì)脂肪具有較強(qiáng)的消化能力。建議在配制花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻飼料時(shí), 適當(dāng)提高粗脂肪比例, 有助于促進(jìn)對(duì)營(yíng)養(yǎng)物質(zhì)的消化吸收, 提高養(yǎng)殖效益。
鹽度; 花鰻鱺; 太平洋雙色鰻鱺; 生長(zhǎng)性能; 消化酶
鹽度是水環(huán)境中重要的生態(tài)因子, 對(duì)魚(yú)類生長(zhǎng)代謝等各種生理活動(dòng)產(chǎn)生影響。同時(shí), 鹽度通過(guò)影響魚(yú)類消化器官中消化酶活力大小來(lái)影響食物的消化吸收, 最終影響魚(yú)類的生長(zhǎng)發(fā)育[1, 2]。鰻鱺屬(Anguilla)魚(yú)類為繁殖洄游性生物, 具有較強(qiáng)的鹽度耐受性, 廣泛分布于淡水、咸水以及半咸水地區(qū)[3]。不同種類的鰻鱺有相似的生活史即受精卵、柳葉鰻、玻璃鰻、幼鰻、黃鰻及銀鰻共6個(gè)階段。一般認(rèn)為鰻鱺在受精卵和柳葉鰻時(shí)期, 生活在海水環(huán)境; 玻璃鰻生活在半咸水的河口地區(qū); 幼鰻和黃鰻溯河進(jìn)入江河湖泊, 進(jìn)行淡水生活; 銀鰻向產(chǎn)卵場(chǎng)作降海洄游, 環(huán)境鹽度不斷上升, 直至進(jìn)入海洋。但Tzeng等[4]利用電子微分析探針檢測(cè)黃鰻期以及銀鰻期日本鰻鱺(A. japonica)耳石中鍶和鈣的比例, 發(fā)現(xiàn)部分日本鰻鱺從幼鰻期到銀鰻期一直生活在海水或半咸水中, 不經(jīng)歷淡水的洄游過(guò)程, 且分布于海水和河口地區(qū)的群體中雌性個(gè)體全長(zhǎng)顯著大于淡水地區(qū)。目前, 鹽度對(duì)鰻鱺屬魚(yú)類生長(zhǎng)的影響已有部分報(bào)道,如日本鰻鱺[5]、歐洲鰻鱺(A. anguilla)[6]、澳洲鰻鱺(A. australis)和大鰻鱺(A. dieffenbachii)[7]。研究表明不同種鰻鱺間對(duì)鹽度的適應(yīng)性存在差異, 而鹽度對(duì)花鰻鱺(A. marmorata)和太平洋雙色鰻鱺(A. bicolor pacifica)生長(zhǎng)性能及消化酶活力的影響尚未見(jiàn)報(bào)道。
花鰻鱺隸屬鰻鱺目、鰻鱺科、鰻鱺屬, 廣泛分布于西太平洋和印度洋的熱帶和亞熱帶地區(qū), 在中國(guó)的東南省份和臺(tái)灣地區(qū)、日本南部、非洲東部、太平洋中部諸島以及印度洋東部及西南群島均有發(fā)現(xiàn)[8]。我國(guó)福建、廣東及浙江等省份有食用花鰻鱺的傳統(tǒng)習(xí)俗, 花鰻鱺肉味鮮美, 成年個(gè)體可長(zhǎng)至40—50 kg, 其市場(chǎng)價(jià)格是其他食用鰻鱺的3—5倍,具有較高的營(yíng)養(yǎng)價(jià)值和經(jīng)濟(jì)價(jià)值。太平洋雙色鰻鱺屬熱帶鰻鱺, 主要分布于西太平洋的赤道地區(qū), 包括婆羅洲東部、新幾內(nèi)亞北部以及菲律賓的呂宋島,與花鰻鱺分布范圍部分重疊, 在河口地區(qū)其鰻苗與前者往往被一同捕撈[9]。近年來(lái), 日本鰻鱺和歐洲鰻鱺苗種資源枯竭, 已嚴(yán)重限制了鰻鱺養(yǎng)殖業(yè)及其加工業(yè)的發(fā)展。東南亞地區(qū)花鰻鱺以及太平洋雙色鰻鱺苗種資源豐富, 價(jià)格便宜, 我國(guó)部分鰻場(chǎng)已開(kāi)始了對(duì)該兩種鰻苗的引進(jìn)試養(yǎng)。本研究對(duì)不同鹽度下花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻生長(zhǎng)性能及消化酶活力變化進(jìn)行比較分析, 旨在了解該兩種鰻鱺對(duì)環(huán)境鹽度的適應(yīng)性, 以期為其洄游生態(tài)生理和人工養(yǎng)殖提供研究資料。
1 材料與方法
1.1 實(shí)驗(yàn)材料與飼養(yǎng)條件
實(shí)驗(yàn)材料為自菲律賓引進(jìn)的花鰻鱺和太平洋雙色鰻鱺玻璃鰻, 后經(jīng)集美大學(xué)福建省海水試驗(yàn)場(chǎng)人工培育1年的幼鰻。挑選外觀正常, 體表無(wú)傷的個(gè)體用于實(shí)驗(yàn)。實(shí)驗(yàn)前, 幼鰻于水族箱中暫養(yǎng)超過(guò)1周,實(shí)驗(yàn)用水為曝氣的自來(lái)水。溫度為(24.5—26.5) , ℃pH保持7.2±0.4, 鹽度<0.3‰, 溶氧≥6.0 mg/L, 氨氮<0.5 mg/L, 亞硝酸鹽<0.2 mg/L, 用日光燈作為光源,光照周期12D : 12L。連續(xù)充氣, 每天08: 00和18: 00 分2次投喂鰻魚(yú)粉狀配合飼料(福建天馬飼料有限公司, 營(yíng)養(yǎng)成分為粗蛋白47%、粗脂肪4%、粗纖維3%、灰分17%、水分10%), 每次投喂前稱量餌料并記錄,采取飽食投喂并根據(jù)殘餌剩余情況適時(shí)調(diào)整投喂量。攝食結(jié)束2h后虹吸排污, 換水率為10%/d。每天觀察實(shí)驗(yàn)魚(yú)活動(dòng)與攝食狀況。
1.2 實(shí)驗(yàn)設(shè)計(jì)
實(shí)驗(yàn)鹽度共設(shè)4個(gè)鹽度梯度, 即淡水對(duì)照組(鹽度0‰)與鹽度5‰、10‰、18‰組, 每梯度設(shè)2個(gè)平行組, 每組20尾幼鰻。花鰻鱺初始體重為(9.76±0.36) g,太平洋雙色鰻鱺初始體重為(11.82±0.04) g。鹽度調(diào)節(jié)用曝氣的自來(lái)水和過(guò)濾的海水配置, 用RHS-28ATC手持型鹽度計(jì)測(cè)定。幼鰻暫養(yǎng)15d后, 開(kāi)始以3‰/24h的速率將鹽度調(diào)整到各實(shí)驗(yàn)設(shè)計(jì)的鹽度梯度并馴養(yǎng)1周。整個(gè)實(shí)驗(yàn)持續(xù)30d, 第15和第30天測(cè)量每組實(shí)驗(yàn)魚(yú)總重。實(shí)驗(yàn)結(jié)束后隨機(jī)選取3尾, 經(jīng)丁香油麻醉后, 在冰盤(pán)上解剖實(shí)驗(yàn)魚(yú), 取出胃、腸道和肝臟。用生理鹽水快速洗凈、濾紙吸干稱重后置于5 mL離心管中, 再加入9倍體積的生理鹽水, 剪碎后用高速勻漿器在冰水浴中以20000 r/min 轉(zhuǎn)速勻漿,勻漿液在Thermo型高速冷凍離心機(jī)(4 ℃, 4000 r/min)中離心10min, 取上清液待測(cè)。
1.3 測(cè)定方法
蛋白含量測(cè)定采用考馬斯亮藍(lán)法測(cè)定, 單位: mg/mL。蛋白酶采用福林-酚試劑法測(cè)定, 酶活力單位定義為: 在37℃條件下, 每毫克組織蛋白每分鐘分解蛋白生成1 μg氨基酸相當(dāng)于1個(gè)酶活力單位(U)。淀粉酶采用碘-淀粉比色法測(cè)定, 酶活力單位定義為: 在37℃條件下, 每毫克組織蛋白與底物作用30min水解10 mg淀粉相當(dāng)于1個(gè)酶活力單位。脂肪酶采用比濁法測(cè)定, 酶活力單位定義為: 在37℃條件下, 每毫克組織在本反應(yīng)體系中與底物反應(yīng)1min消耗1 μmol底物相當(dāng)于1個(gè)酶活力單位。消化酶活力采用比活力表示, 單位: U/(mg·min)。
1.4 數(shù)據(jù)處理
在本研究中花鰻鱺與太平洋雙色鰻鱺生長(zhǎng)性能采用日增重、特定生長(zhǎng)率和餌料系數(shù)表示, 計(jì)算公式如下:
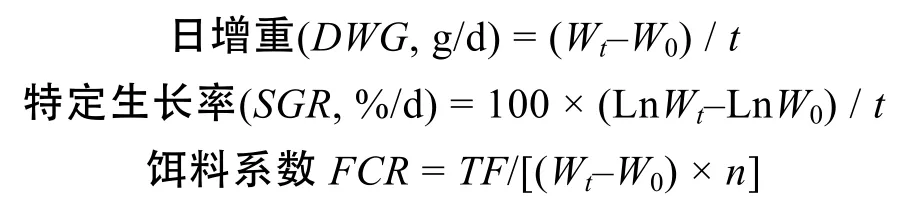
上式中, W0和Wt分別為實(shí)驗(yàn)初始和實(shí)驗(yàn)?zāi)┯做犉骄w重, t為實(shí)驗(yàn)天數(shù), TF為實(shí)驗(yàn)階段總攝食量, n為實(shí)驗(yàn)初幼鰻尾數(shù)。
所有數(shù)據(jù)采用 SPSS 13.0軟件進(jìn)行處理, 平均數(shù)采用均值±標(biāo)準(zhǔn)誤(Mean±SE)表示, 各鹽度梯度組間生長(zhǎng)性能指標(biāo)和酶活力的差異采用單因素方差分析(One-way ANOVA)和 Duncan檢驗(yàn)法比較, 當(dāng)P<0.05時(shí)認(rèn)為差異顯著, P<0.01 時(shí)差異極顯著。
2 結(jié)果
2.1 鹽度對(duì)生長(zhǎng)性能的影響
在不同鹽度下花鰻鱺幼鰻的體重變化、存活率和日增重(DWG)情況見(jiàn)表1, 太平洋雙色鰻鱺見(jiàn)表2。花鰻鱺和太平洋雙色鰻鱺在各鹽度處理中存活率均為100%, 未出現(xiàn)死亡。在各鹽度處理中, 兩種鰻鱺的生長(zhǎng)存在差異。實(shí)驗(yàn)?zāi)? 鹽度0‰和鹽度5‰處理組花鰻鱺體重顯著大于鹽度10‰和18‰處理組(P<0.05), 鹽度0‰處理組太平洋雙色鰻鱺體重顯著大于其他組(P<0.05)。鹽度0‰處理組花鰻鱺的DWG顯著大于鹽度5‰處理組(P<0.05), 而鹽度10‰和18‰處理組體重下降, 出現(xiàn)負(fù)生長(zhǎng)。鹽度0‰處理組太平洋雙色鰻鱺的DWG顯著大于其他組(P<0.05),各組均無(wú)負(fù)生長(zhǎng)。

表1 不同鹽度下花鰻鱺幼鰻的體重、存活率和日增重Tab. 1 The weight, the survival rate and the daily weight gain of A. marmorata elver at different salinities

表2 不同鹽度下太平洋雙色鰻鱺幼鰻的體重、存活率和日增重Tab. 2 The weight, the survival rate and the daily weight gain of A. bicolor pacifica elver at different salinities
由圖1可知, 4個(gè)鹽度水平下太平洋雙色鰻鱺的特定生長(zhǎng)率(SGR)均顯著大于花鰻鱺(P<0.05)。鹽度0‰處理組花鰻鱺的SGR大于鹽度5‰處理組, 但差異不顯著性(P>0.05); 鹽度0‰處理組太平洋雙色鰻鱺的SGR最大, 并顯著性大于其他組(P<0.05)。
由圖2可知, 鹽度對(duì)花鰻鱺和太平洋雙色鰻鱺餌料系數(shù)(FCR)的影響與SGR不同。在鹽度0‰和5‰時(shí),花鰻鱺的FCR顯著大于太平洋雙色鰻鱺(P<0.05)。花鰻鱺在鹽度0‰時(shí)FCR最小, 低于其他組。太平洋雙色鰻鱺的FCR隨鹽度升高而增大, 鹽度0‰時(shí)FCR最小, 顯著低于其他組(P<0.05), 鹽度18‰時(shí)最大, 鹽度5‰和10‰處理組間差異不顯著(P>0.05)。
2.2 鹽度對(duì)花鰻鱺幼鰻消化酶活力的影響
在4個(gè)鹽度處理中, 鹽度相同時(shí)花鰻鱺3個(gè)消化器官的蛋白酶活力均為肝臟最低為0.23—1.71 U/(mg·min),腸道其次為 1.12—3.29 U/(mg·min), 胃最高為5.81—8.34 U/(mg·min) (圖3)。鹽度對(duì)肝臟、胃和腸道中蛋白酶活力的影響存在差異, 其中鹽度 18‰處理組中肝臟蛋白酶活力顯著高于其他鹽度組(P<0.05),
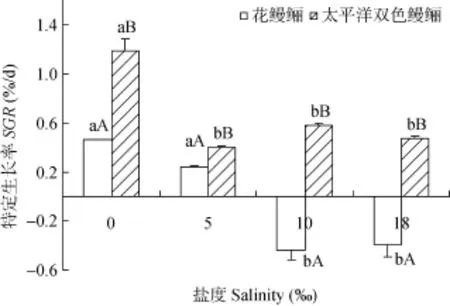
圖 1 在不同鹽度下花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻的特定生長(zhǎng)率比較Fig. 1 Comparison of SGR for A. marmorata elver and A. bicolor pacifica elver at different salinities
圖標(biāo)上方不同小寫(xiě)字母, 表示同種鰻鱺不同鹽度間存在顯著性差異(P<0.05); 不同大寫(xiě)字母, 表示同種鹽度下兩種鰻鱺間存在顯著性差異(P<0.05); 下同
Different small letters on the columns indicate significant difference between different salinities for the same specie (P<0.05), and different capital letter superscripts indicated significant differences between two species at the same salinity (P<0.05). The same applies below胃和腸道中蛋白酶活力在鹽度0‰時(shí)均顯著最高(P<0.05), 其他鹽度處理中均無(wú)顯著性差異(P>0.05)。

圖 2 在不同鹽度下花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻的餌料系數(shù)比較Fig. 2 Comparison of FCR for A. marmorata elver and A. bicolor pacifica elver at different salinities

圖3 在不同鹽度下花鰻鱺幼鰻肝臟、胃和腸道蛋白酶活力Fig. 3 The activity of protease in the liver, the stomach and the intestine of A. marmorata elver at different salinities
圖4顯示, 各鹽度下花鰻鱺腸道淀粉酶活力為0.58—1.58 U/(mg·min), 鹽度相同時(shí), 均顯著高于肝臟和胃(P<0.05)。比較同種消化器官不同鹽度處理中的淀粉酶活力可知, 肝臟淀粉酶活力在各鹽度處理中均無(wú)顯著性差異(P>0.05); 胃淀粉酶活力與鹽度梯度呈負(fù)相關(guān), 鹽度0‰時(shí)最高, 鹽度18‰時(shí)最低;在鹽度0‰時(shí), 腸道淀粉酶活力最高, 顯著高于其他鹽度組(P<0.05), 而其他3個(gè)鹽度組之間均無(wú)顯著性差異(P>0.05)。
圖5顯示, 在4個(gè)鹽度下花鰻鱺腸道脂肪酶活力為26.38—192.53 U/(mg·min), 在鹽度相同時(shí), 除鹽度 18‰處理組, 均顯著高于肝臟和胃(P<0.05),
肝臟和胃脂肪酶活力在各鹽度處理組間均無(wú)顯著性差異(P>0.05)。比較同種消化器官不同鹽度處理中的脂肪酶活力可知, 肝臟脂肪酶活力隨鹽度增加而下降, 鹽度 0‰和5‰時(shí)顯著高于鹽度18‰ (P<0.05);
在鹽度 0‰時(shí), 胃脂肪酶活力最高, 顯著高于鹽度
5‰和10‰處理組(P<0.05), 而其他3個(gè)鹽度組之間均無(wú)顯著性差異(P>0.05); 腸道脂肪酶活力與鹽度呈負(fù)相關(guān), 鹽度 0‰處理組最高且顯著高于其他組
(P<0.05), 鹽度18‰處理組最低。
2.3 鹽度對(duì)太平洋雙色鰻鱺幼鰻消化酶活力的影響如圖6所示, 在4個(gè)鹽度下太平洋雙色鰻鱺胃和腸道蛋白酶活力分別為2.86—7.01和2.24—
4.76 U/(mg·min)。在鹽度梯度相同時(shí), 除鹽度18‰
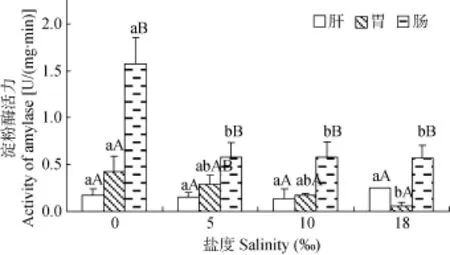
圖4 在不同鹽度下花鰻鱺幼鰻肝臟、胃和腸道淀粉酶活力Fig. 4 The activity of amylase in the liver, the stomach and the intestine of A. marmorata elver at different salinities
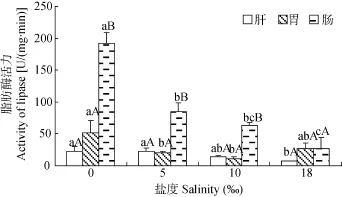
圖5 在不同鹽度下花鰻鱺幼鰻肝臟、胃和腸道脂肪酶活力Fig. 5 The activity of lipase in the liver, the stomach and the intestine of A. marmorata elver at different salinities
處理組, 均顯著高于肝臟(P<0.05), 胃和腸道淀粉酶活力在各鹽度下均無(wú)顯著性差異(P>0.05)。不同鹽度處理對(duì)同種消化器官蛋白酶活力的影響存在差異, 在鹽度18‰時(shí), 肝臟蛋白酶活力最高, 顯著高于其他鹽度組(P<0.05), 而其他3個(gè)鹽度組之間均無(wú)顯著性差異(P>0.05); 在鹽度10‰時(shí), 胃蛋白酶活力最高, 顯著高于鹽度5‰和18‰處理組(P<0.05),與鹽度0‰處理組無(wú)顯著性差異(P>0.05); 腸道蛋白酶活力在各鹽度處理中均無(wú)顯著性差異(P>0.05)。
如圖7所示, 在4個(gè)鹽度處理中太平洋雙色鰻鱺腸道淀粉酶活力為0.52—0.80 U/(mg·min), 在鹽度相同時(shí), 均顯著高于肝臟和胃(P<0.05), 肝臟和胃淀粉酶活力在各鹽度處理組之間均無(wú)顯著性差異(P>0.05)。比較同種消化器官不同鹽度處理中的淀粉酶活力可知, 肝臟和腸道淀粉酶活力在各鹽度處理中均無(wú)顯著性差異(P>0.05); 在鹽度18‰時(shí), 胃淀粉酶活力最高, 顯著高于鹽度10‰處理組(P<0.05),而與鹽度0和5‰處理組之間無(wú)顯著性差異(P>0.05)。

圖6 在不同鹽度下太平洋雙色鰻鱺幼鰻肝臟、胃和腸道蛋白酶活力Fig. 6 The activity of protease in the liver, the stomach and the intestine of A. bicolor pacifica elver at different salinities

圖7 在不同鹽度下太平洋雙色鰻鱺幼鰻肝臟、胃和腸道淀粉酶活力Fig. 7 The activity of amylase in the liver, the stomach and the intestine of A. bicolor pacifica elver at different salinities
如圖 8所示, 在各鹽度下太平洋雙色鰻鱺腸道脂肪酶活力為 67.50—92.08 U/(mg·min), 在鹽度相同時(shí), 除鹽度 10處理組, 均顯著高于肝臟和胃(P<0.05), 肝臟和胃脂肪酶活力在各鹽度下均無(wú)顯著性差異(P>0.05)。比較同種消化器官不同鹽度處理中的脂肪酶活力可知, 肝臟、胃和腸道脂肪酶活力在各鹽度處理中均無(wú)顯著性差異(P>0.05)。

圖8 在不同鹽度下太平洋雙色鰻鱺幼鰻肝臟、胃和腸道脂肪酶活力Fig. 8 The activity of lipase in the liver, the stomach and the intestine of A. bicolor pacifica elver at different salinities
3 討論
3.1 鹽度對(duì)花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻生
長(zhǎng)性能的影響
鹽度是影響魚(yú)類生長(zhǎng)代謝等各種生理活動(dòng)的重要環(huán)境因素, 其變化迫使魚(yú)類自身通過(guò)一系列生理變化來(lái)調(diào)整體內(nèi)外滲透壓的動(dòng)態(tài)平衡, 從而對(duì)魚(yú)體與環(huán)境間物質(zhì)交換與能量流動(dòng)產(chǎn)生影響[10]。魚(yú)類種類繁多, 生物學(xué)特征及生活環(huán)境存在差異, 因此其滲透調(diào)節(jié)方式與機(jī)理及能量消耗的關(guān)系也存在較大差異。環(huán)境鹽度對(duì)魚(yú)類生長(zhǎng)的影響存在以下兩種現(xiàn)象: (1)魚(yú)類基本處于等滲環(huán)境下(內(nèi)環(huán)境滲透壓一般維持在300—400 mOsm/kg, 即鹽度為10‰—15‰的水體), 生長(zhǎng)率最高, 餌料系數(shù)最低, 如弓斑東方鲀(Fugu obscurus)[11]、小鋸蓋魚(yú)(Centropomus parallelus)[12]、 暗紋東方 鲀(Takifugu obscures)[13]等;(2)魚(yú)類在低滲或者高滲環(huán)境下, 生長(zhǎng)率最高, 餌料系數(shù)最低, 如歐洲鱸(Dicentrarchus labrax)[14]、波紋短須石首魚(yú)(Umbrina cirrosa)[15]、高體革(Scortum barcoo)[16]等。本實(shí)驗(yàn)結(jié)果與后者相符, 花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻均在淡水中生長(zhǎng)良好, 特定生長(zhǎng)率高于各鹽度組, 而餌料系數(shù)低于各鹽度組。王云峰等[10]認(rèn)為魚(yú)類滲透壓調(diào)節(jié)的能量消耗極小, 可能并不足以影響生長(zhǎng)。而鹽度變化不僅作用于魚(yú)類的滲透壓調(diào)節(jié), 對(duì)其攝食行為、食欲、新陳代謝的刺激或抑制以及激素的傳導(dǎo)途徑都產(chǎn)生影響[17]。Kearney等[7]的研究結(jié)果顯示, 水溫 17.5℃時(shí), 養(yǎng)殖于鹽度 0‰、17.5‰和 35‰水體中的澳洲鰻鱺玻璃鰻特定生長(zhǎng)率無(wú)顯著性差異(P>0.05), 水溫26.5℃時(shí), 養(yǎng)殖于鹽度0‰和 17.5‰水體中的澳洲鰻鱺玻璃鰻特定生長(zhǎng)率顯著高于鹽度 35‰組; 在水溫 17.5℃和 26.5℃時(shí),大鰻鱺玻璃鰻均在鹽度 0‰組中獲得最大特定生長(zhǎng)率。這表明鹽度對(duì)鰻鱺屬魚(yú)類生長(zhǎng)的影響同時(shí)受到溫度的作用, 該作用具有一定的種類特異性。另外,鹽度對(duì)不同生活史階段的同種鰻鱺屬魚(yú)類生長(zhǎng)的影響也存在差異, 如 Okamura等[5]比較了日本鰻鱺柳葉鰻在不同鹽度梯度下的生長(zhǎng)性能, 發(fā)現(xiàn)其在海水濃度為30%和50%的水體(即鹽度10‰和17.5‰)養(yǎng)殖 7d后, 體寬顯著增加(P<0.05), 而養(yǎng)殖于海水濃度0、10%、70%和100%的個(gè)體體寬無(wú)顯著性變化(P>0.05); Tzeng等[4]發(fā)現(xiàn)分布于海洋和河口地區(qū)(即鹽度 17.5‰—35‰)的日本鰻鱺(A. japonica)幼成鰻雌性個(gè)體生長(zhǎng)速率顯著高于淡水個(gè)體。
3.2 鹽度對(duì)花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻消化酶活力的影響
魚(yú)類對(duì)營(yíng)養(yǎng)物質(zhì)的消化、吸收需要消化酶的參與, 酶活力的高低在一定程度上反映了魚(yú)類消化、吸收能力的大小, 而鹽度可通過(guò)影響消化酶活力大小來(lái)影響魚(yú)類對(duì)食物的消化吸收。在一定范圍內(nèi)鹽度的改變會(huì)導(dǎo)致魚(yú)體消化酶活力的變化, 主要分為3種情況: 激活作用、無(wú)影響、抑制作用。在本實(shí)驗(yàn)中, 鹽度對(duì)花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻消化酶活力的影響存在差異, 其中各鹽度處理組的花鰻鱺蛋白酶活力無(wú)顯著變化, 淀粉酶和脂肪酶活力均隨鹽度的增加而下降; 太平洋雙色鰻鱺胃蛋白酶在鹽度10‰時(shí)最大, 肝蛋白酶在鹽度18‰時(shí)最大, 而淀粉酶和脂肪酶活力在各鹽度處理組無(wú)顯著變化。這表明鹽度的增加對(duì)花鰻鱺的淀粉酶和脂肪酶活力具有抑制作用, 對(duì)太平洋雙色鰻鱺的蛋白酶活力有一定的激活作用, 這與鹽度對(duì)兩種鰻鱺生長(zhǎng)性能的影響基本相符。許多研究表明, 海水中部分無(wú)機(jī)離子對(duì)酶具有激活或抑制作用, 無(wú)機(jī)離子直接作用于消化酶是鹽度影響魚(yú)體內(nèi)消化酶活力的主要原因[18]。如較低濃度的氯離子對(duì)虱目魚(yú)(Chanos chanos)腸道、胰腺、幽門(mén)盲囊和肝臟的α-淀粉酶有激活作用, 而較高濃度的氯離子抑制α-淀粉酶[19]。也有學(xué)者認(rèn)為海水的攝入, 改變了消化器官內(nèi)環(huán)境的pH, 從而引起消化酶活力的變化。而當(dāng)消化酶活力隨著鹽度的改變會(huì)發(fā)生變化時(shí), 魚(yú)類可以通過(guò)產(chǎn)生同工酶保持正常的消化水平[20]。
魚(yú)類對(duì)各種消化酶活力的比值, 可看成是對(duì)餌料中蛋白質(zhì)、糖類和脂類物質(zhì)消化能力的比較。一般認(rèn)為雜食性魚(yú)類和草食性魚(yú)類的蛋白酶活力與淀粉酶活力比值低于肉食性魚(yú)類[21]。在本實(shí)驗(yàn)中花鰻鱺和太平洋雙色鰻鱺各消化器官中蛋白酶活力均遠(yuǎn)大于淀粉酶活力, 表現(xiàn)出肉食性的特點(diǎn)。另外, 兩種鰻鱺的脂肪酶活力在各鹽度組中均較高, 與其肌肉中粗脂肪含量較高相符合, 其中花鰻鱺粗脂肪含量為鮮重的8.62%, 太平洋雙色鰻鱺為17.37%(另文發(fā)表)。因此, 適當(dāng)提高兩種鰻魚(yú)幼鰻飼料中粗脂肪比例, 有助于促進(jìn)消化吸收, 加快生長(zhǎng), 提高養(yǎng)殖效益。
3.3 不同消化器官間的消化酶活力差異
在本實(shí)驗(yàn)結(jié)果中, 養(yǎng)殖在各鹽度水體的花鰻鱺幼鰻和太平洋雙色鰻鱺幼鰻腸道的淀粉酶和脂肪酶活力均顯著高于胃和肝臟, 胃蛋白酶活力高于肝臟和腸道。這表明腸道是花鰻鱺和太平洋雙色鰻鱺消化淀粉和脂肪的主要場(chǎng)所, 而蛋白質(zhì)的消化吸收主要在胃中進(jìn)行。這與田田等[6]對(duì)歐洲鰻鱺幼鰻蛋白酶研究的結(jié)果基本一致, 而與黃永春等[22]對(duì) 4種不同規(guī)格歐洲鰻鱺淀粉酶和蛋白酶檢測(cè)結(jié)果不同。其原因可能與實(shí)驗(yàn)魚(yú)規(guī)格大小有關(guān), 在本研究中兩種鰻鱺規(guī)格[(9.76±0.36) g、(11.82±0.04) g]均與前者實(shí)驗(yàn)魚(yú)規(guī)格[(7.2±1.4) g]相似, 而遠(yuǎn)小于后者所用實(shí)驗(yàn)魚(yú)規(guī)格(51.5—233 g)。
[1] Guo Q D, Wang Y J, Lü W Q. Combined effects of temperature and salinity on the physiological osmotic induction and antioxidant responses in the juvenile Japanese flounder (Paralichthys olivaceus) [J]. Acta Hydrobiologica Sinica, 2014, 38(1): 58—67 [郭勤單, 王有基, 呂為群. 溫度和鹽度對(duì)褐牙鲆幼魚(yú)滲透生理及抗氧化水平的影響.水生生物學(xué)報(bào), 2014, 38(1): 58—67]
[2] Yan M J, Li Z J, Xiong B X. Food intake, growth and feed utilization of puffer fish (Takifugu fasciatus) after different salinity pretreatments [J]. Acta Hydrobiologica Sinica, 2005, 29(2): 142—145 [嚴(yán)美姣, 李鐘杰, 熊邦喜. 不同鹽度預(yù)處理后Ⅰ 鲀齡暗紋東方 的攝食、生長(zhǎng)和飼料利用. 水生生物學(xué)報(bào), 2005, 29(2): 142—145]
[3] Eric E, Pierre E. Is salinity choice related to growth in juvenile eel Anguilla Anguilla [J]? Cybium, 2004, 28(suppl): 77—82
[4] Tzeng W N, Lizuka Y, Shiao J C, et al. Identification and growth rate comparison of divergent migratory contingents of Japanese eel (Anguilla japonica) [J]. Aquaculture, 2003, 216(1-4): 77—86
[5] Okamura A, Yamada Y, Mikawa N, et al. Growth and survival of eel leptocephali (Anguilla japonica) in lowsalinity water [J]. Aquaculture, 2009, 296(3—4): 367—372
[6] Tian T, Yuan C G, Ruan C X. The study on variation of growth of Anguilla anguilla elver at different salinities [J]. Journal of Fujian Fisheries, 2006, 111(4): 63—66 [田田, 袁重桂, 阮成旭. 歐鰻黑仔苗在不同鹽度條件下生長(zhǎng)效果的比較. 福建水產(chǎn), 2006, 111(4): 63—66]
[7] Kearney M, Jeffs A, Lee P. Effects of salinity and temperature on the growth and survival of New Zealand shortfin, Anguilla australis, and longfin, A. dieffenbachii, glass eels [J]. Aquaculture Research, 2008, 39(16): 1769—1777
[8] Jamandre B W D, Shen K N, Yambot A V, et al. Molecular phylogeny of Philippine freshwater eels Anguilla spp. (Actinopterygi: Anguilliformes: Anguillidae) inferred from mitochondrial DNA [J]. The Raffles Bulletin of Zoology, 2007, (suppl): 51—59
[9] Han Y S, Chang C W, He J T, et al. Validation of the occurrence of short-finned eel Anguilla bicolor pacifica in natural waters of Taiwan [J]. Acta Zoologica Taiwanica, 2001, 12(1): 9—19
[10] Luo M Z, Guan R Z, Jin H. Effects of salinity on Na+/K+-ATPase activity in gill and kidney of Anguilla marmorata and A. bicolor pacifica elver [J]. Oceanologia et Limnologia Sinica, 2013, 44(3): 807—813 [羅鳴鐘, 關(guān)瑞章, 靳恒. 鹽度對(duì)花鰻鱺(Anguilla marmorata) 和太平洋雙色鰻鱺(A. bicolor pacifica) 幼鰻鰓絲及腎臟Na+/K+-ATP酶活力的影響. 海洋與湖沼, 2013, 44(3): 807—813]
[11] Yan M, Li Z, Xiong B, et al. Effects of salinity on food intake, growth, and survival of pufferfish (Fugu obscurus) [J]. Journal of Applied Ichthyology, 2004, 20(2): 146—149
[12] Tsuzukia M Y, Sugaib J K, Maciela J C, et al. Survival, growth and digestive enzyme activity of juveniles of the fat snook (Centropomus parallelus) reared at different salinities [J]. Aquaculture, 2007, 271(1—4): 319—325
[13] Bian P J, Qiu C G, Xu SL, et al. Effects of salinity on growth, activity of non-specific immune and antioxidant enzymes in obscure puffer Takifugu obscures [J]. Acta Hydrobiologica Sinica, 2014, 38(1): 108—114 [邊平江, 邱成功, 徐善良, 等. 鲀鹽度對(duì)暗紋東方 生長(zhǎng)、非特異性免疫和抗氧化酶活力的影響. 水生生物學(xué)報(bào), 2014, 38(1): 108—114]
[14] Conides A J, Glamuzina B. Laboratory simulation of the effects of environmental salinity on acclimation, feeding and growth of wild-caught juveniles of European sea bass Dicentrarchus labrax and gilthead sea bream, Sparus aurata [J]. Aquaculture, 2006, 256(1—4): 235—245
[15] Mylonas C C, Pavlidis M, Papandroulakis N, et al. Growth performance and osmoregulation in the shi drum (Umbrina cirrosa) adapted to different environmental salinities [J]. Aquaculture, 2009, 287(1—2): 203—210
[16] Liang Y J, Sun X J, Shi D J, et al. Effects of salinity on growth and flesh quality of Scortum barcoo [J]. Acta Hydrobiologica Sinica, 2010, 34(4): 880—884 [梁擁軍, 孫向軍, 史東杰, 等. 鯻鹽度對(duì)高體革 生長(zhǎng)和肉質(zhì)的影響.水生生物學(xué)報(bào), 2010, 34(4): 880—884]
[17] Boeuf G, Payan P. How should salinity influence fish growth [J]? Comparative Biochemistry and Physiology part C, 2001, 130(4): 411—423
[18] Tian X L, Ren X W, Dong S L, et al. Studies on the specific activities of digestive enzymes of Cynoglossus semilaevis Gunther at different salinities and temperatures [J]. Periodical of Ocean University of China, 2008, 38(6): 895—901 [田相利, 任曉偉, 董雙林, 等. 溫度和鹽度對(duì)半滑舌鰨幼魚(yú)消化酶活性的影響. 中國(guó)海洋大學(xué)學(xué)報(bào), 2008, 38(6): 895—901]
[19] Chiu Y N, Benitez L V. Studies on the carbohydrates in the digestive tract of the milkfish Chanos chanos [J]. Marine Biology, 1981, 61(2—3): 247—254
[20] Tang B G, Chen G, Zhang J D, et al. Effects of pH, amylum concentration and temporary-keeping salinity on the activity of amylase in the digestive tract of Lautianus erythopterus [J]. Chinese Journal of Zoology, 2004, 39(2): 70—73 [湯保貴, 陳剛, 張健東, 等. pH、底物濃度及暫養(yǎng)鹽度對(duì)紅鰭笛鯛消化道淀粉酶活力的影響. 動(dòng)物學(xué)雜志, 2004, 39(2): 70—73]
[21] Hofer R, Schiemer F. Proteolytic activity in the digestive tract of several species of fish with different feeding habits [J]. Oecologia, 1981, 48(3): 342—345
[22] Huang Y C, Liu D. Preliminary study on the effect of temperature on proteases and amylases activities from different sizes European eel (Anguillia anguillia) [J]. Journal of Oceanography in Taiwan Strait, 2004, 23(2): 138—143 [黃永春, 劉登. 溫度對(duì)不同規(guī)格歐鰻消化器官蛋白酶和淀粉酶活性影響的初步研究. 臺(tái)灣海峽, 2004, 23(2): 138—143]
EFFECTS OF THE SALINITY ON THE GROWTH PERFORMANCE AND DIGESTIVE ENZYME ACTIVITIES OF ANGUILLA MARMORATA ELVER AND A. BICOLOR PACIFICA ELVER
LUO Ming-Zhong1, GUAN Rui-Zhang2, 3and JIN Heng1
(1. College of Animal Science, Yangtze University, Jingzhou 434025, China; 2. Engineering Research Centre of Eel Modern Technical Industry, Ministry of Education, Xiamen 361021, China; 3. Fisheries College, Jimei University, Xiamen 361021, China)
In this study, we investigated the effects of salinity on the growth and the activities of the digestive enzymes of Anguilla marmorata elver [body weight (9.76 ± 0.36) g] and A. bicolor pacifica elver [body weight (11.82 ± 0.04) g]. Subjects of the two species were cultured at four salinities (0‰, 5‰, 10‰ and 18‰) for 30 days. We first weighed all the eels in each tank, and then analyzed three types of digestive enzymes (protease, amylase and lipase) in three digestive organs (liver, stomach and intestine) of both species. The results showed that the survival rates of the two species were both 100% in all four salinities with no death case. In fresh water both species exhibited excellent growth performance, the highest specific growth rates (SGR), and the lowest food conversion ratios (FCR). The effects of salinity on the digestive enzyme activities were different between the two species. In the three digestive organs, the salinity did not significantly affect the activity of protease in A. marmorata elver (P>0.05), but the increase in salinity caused reduced activities of amylase and lipase. For A. bicolor pacifica elver, the highest activity of protease in the stomach and the liver appeared at the salinities of 10‰ and 18‰ respectively; whereas the salinity did not affect the activities of amylase and lipase in the three digestive organs (P>0.05). We deducted that saline water could inhibit the activities of amylase and lipase in the liver, the stomach and the intestine of A. marmorata elver, but might increase the protease activity of A. bicolor pacifica elver. At the same salinity the same digestive enzymes in different digestive organs showed different activities. For both species the activities of amylase and lipase in the intestine were significantly higher than in the liver and the stomach (P<0.05); the activity of protease in the stomach was insignificantly higher than in the liver and the intestine (P>0.05). Our study suggested that the high activity of lipase enabled A.marmorata elver and A. bicolor pacifica elver to digest fat efficiently. Therefore it may be suitable to increase the portion of the crude fat in the compounding feed, which could boost the digestion and absorption of nutrients and thus improve the quality of aquaculture.
Salinity; Anguilla marmorata; A. bicolor pacifica; Growth performance; Digestive enzyme
S965.223
A
1000-3207(2015)04-0653-08
10.7541/2015.87
2014-07-21;
2014-12-12
國(guó)家農(nóng)業(yè)部公益性行業(yè)(農(nóng)業(yè))科研專項(xiàng)(nyhyzx07-043-03)資助
羅鳴鐘(1984—), 男, 湖北黃石人; 博士, 講師; 主要從事水產(chǎn)養(yǎng)殖學(xué)研究。E-mail: kklmz413@hotmail.com
關(guān)瑞章, E-mail: rzguan@jmu.edu.cn
