養(yǎng)殖中華鱘的產(chǎn)后康復(fù)
2015-02-28 08:28:25張曉雁杜浩危起偉張艷珍王彥鵬蔡經(jīng)江劉志剛喬新美熊
水生生物學(xué)報 2015年4期
張曉雁杜 浩危起偉張艷珍王彥鵬蔡經(jīng)江劉志剛喬新美熊 偉
(1. 中國水產(chǎn)科學(xué)研究院長江水產(chǎn)研究所, 農(nóng)業(yè)部淡水生物多樣性保護(hù)重點實驗室, 武漢 430223; 2. 北京信沃達(dá)海洋科技有限公司(北京海洋館), 北京 100081)
養(yǎng)殖中華鱘的產(chǎn)后康復(fù)
張曉雁1, 2杜 浩1危起偉1張艷珍2王彥鵬2蔡經(jīng)江2劉志剛1喬新美1熊 偉1
(1. 中國水產(chǎn)科學(xué)研究院長江水產(chǎn)研究所, 農(nóng)業(yè)部淡水生物多樣性保護(hù)重點實驗室, 武漢 430223; 2. 北京信沃達(dá)海洋科技有限公司(北京海洋館), 北京 100081)
通過觀測初次繁殖養(yǎng)殖中華鱘(Acipenser sinensis)親本產(chǎn)后的攝食行為, 測量生長指標(biāo)和監(jiān)測血清中甲狀腺指標(biāo)、血液理化指標(biāo), 進(jìn)行產(chǎn)后親本11個月康復(fù)的效果分析。結(jié)果表明: 使用多春魚(Mallotus villosus)可誘導(dǎo)產(chǎn)后親本開口攝食及轉(zhuǎn)食營養(yǎng)豐富的混合鮮餌。雌性產(chǎn)后身體虛弱, 攝食時行為異常, 隨攝入營養(yǎng)水平逐漸提高, 雌性和雄性最大攝食量于產(chǎn)后7個月時分別達(dá)到體質(zhì)量的2.04%和1.60%, 7個月后恢復(fù)正常。隨混合鮮餌攝食量提高, 親本體質(zhì)量從下降轉(zhuǎn)而升高(9個月間雌雄親本增長率分別為44.16%和23.30%), 體長增長(增長率分別為5.00%和3.23%), 與同齡未產(chǎn)的養(yǎng)殖中華鱘(參照組)相比, 體質(zhì)量和體長的增長率均處于較高水平。產(chǎn)后親本血清總T3(TT3)、血清總T4(TT4)、血清游離T3(FT3)、血清游離T4(FT4)和多項血液理化指標(biāo)在恢復(fù)培養(yǎng)期明顯升高, 至培養(yǎng)期末, TT3 、FT3高于參照組, 其余指標(biāo)多接近參照組平均水平, 表明產(chǎn)后親本生理情況基本恢復(fù)。研究表明, 加強(qiáng)產(chǎn)后康復(fù)培養(yǎng), 可以提高初次繁殖親本, 尤其是雌性的健康恢復(fù)和生長效果, 對養(yǎng)殖中華鱘資源的重復(fù)利用和再次成熟后生殖力的提高至關(guān)重要。
中華鱘; 產(chǎn)后; 攝食行為; 生長; 血清中甲狀腺指標(biāo); 血液生理指標(biāo); 血清生化指標(biāo)
中華鱘(Acipenser sinensis)是國家一級保護(hù)動物。由于人類活動的干擾, 自然繁殖群體數(shù)量急劇減少, 自然種群滅絕的危險提高[1]。開展全人工繁殖是實現(xiàn)人工保種的有效途徑[2]。由于中華鱘體型大, 性成熟晚, 性周期時間長, 養(yǎng)殖過程風(fēng)險大、成本高,養(yǎng)殖達(dá)到初次性成熟的子代個體不僅數(shù)量有限, 相比于長江野生雌性最小性成熟個體, 其體質(zhì)量和懷卵量亦明顯減小[2, 3]。另外, 就野生產(chǎn)后個體而言, 經(jīng)歷長距離繁殖洄游期間不攝食, 繁殖后體質(zhì)虛弱, 易引起死亡, 通過人工干預(yù)法可克服恢復(fù)攝食的障礙,獲得康復(fù)及性腺再次發(fā)育[4, 5]。對于養(yǎng)殖中華鱘親本,與野生相比, 沒有產(chǎn)卵洄游前的育肥階段, 性腺發(fā)育期間多停食, 產(chǎn)后恢復(fù)困難[2, 6]。因此, 進(jìn)行養(yǎng)殖親本產(chǎn)后康復(fù)和生長研究, 是促進(jìn)產(chǎn)后親本成活率和再次成熟后生殖力提高的關(guān)鍵, 為開展養(yǎng)殖中華鱘資源的重復(fù)利用提供支撐。同時, 通過產(chǎn)后親魚培育效果分析, 為產(chǎn)前親魚培育技術(shù)提供借鑒。
本研究通過對養(yǎng)殖產(chǎn)后親本的攝食行為進(jìn)行觀察、測量生長指標(biāo)及監(jiān)測血清中甲狀腺指標(biāo)和血液理化指標(biāo), 通過與同齡性腺未成熟的養(yǎng)殖個體(參照組)比較, 以確定養(yǎng)殖親本恢復(fù)效果, 為建立養(yǎng)殖親本產(chǎn)后恢復(fù)培養(yǎng)技術(shù)提供指導(dǎo)。本研究對中華鱘物種保護(hù)具有現(xiàn)實意義。
1 材料與方法
1.1 養(yǎng)殖設(shè)施和養(yǎng)殖時間
本文產(chǎn)后恢復(fù)培養(yǎng)從2012年11月至次年9月止, 共計11個月。在臨時養(yǎng)殖池(養(yǎng)殖池的池體規(guī)格32.0 m×12.0 m×3.8 m, 水溫19.0—19.5 , ℃ 溶氧6.0—8.4 mg/L, 流速 0.2—0.5 m/s)培養(yǎng)前 2個月, 于12 月 28日轉(zhuǎn)入馴養(yǎng)池(馴養(yǎng)池的池體規(guī)格 29.0 m× 11.0 m×4.4 m, 展示觀察窗規(guī)格20.0 m×3.0 m, 水溫21.0—22.5 , ℃ 溶氧7.0—7.8 mg/L, 循環(huán)水過濾系統(tǒng)的循環(huán)率為50 min/次, 養(yǎng)殖密度2.2 kg/m3)繼續(xù)產(chǎn)后恢復(fù)培養(yǎng)9個月。
1.2 養(yǎng)殖親本和飼養(yǎng)管理
雌、雄產(chǎn)后親本[2](編號F51、F38)分別為長江水產(chǎn)研究所2001年和1998年秋季通過野生中華鱘人工繁殖的養(yǎng)殖個體, 于2012年10月28—30日實施人工催產(chǎn), 繁殖前、后即刻測得的體長分別為180、180 cm和186、186 cm; 體質(zhì)量分別為57、51 kg和64、58 kg。
在同一養(yǎng)殖條件下, 接近性成熟年齡的未產(chǎn)個體(12齡養(yǎng)殖個體), 血清Mg2+、TP、GLB、ALP和Ht值(本文檢測的血液生理、生化指標(biāo))略低于處于性腺發(fā)育間隔期的野生成熟個體[7]。本文為分析產(chǎn)后親本的恢復(fù)情況, 在馴養(yǎng)池內(nèi)隨機(jī)選取健康的雌雄魚各4尾(同為長江水產(chǎn)研究所人工繁殖個體, 分別與雌雄親本同齡, 處于性腺慢速發(fā)育階段)作為參照組, 12月體長分別為(206±6)和(189±4) cm, 體質(zhì)量為(89±9)和(72±4) kg。
產(chǎn)后親本采用直接投灑冰鮮多春魚(Mallotus villosus)的方式誘導(dǎo)開口。在轉(zhuǎn)入馴養(yǎng)池后, 由潛水員水下采用多春魚攜帶方式馴化投喂混合餌料, 逐漸增加混合餌料量, 直至完全攝食。每次投喂量以自主攝食量為主, 隔日喂食。同期, 參照組采用混合餌料由潛水員水下投喂, 平均投餌率為體質(zhì)量的0.91%。將一定比例的冰鮮多春魚、藍(lán)點馬鮫(Scomberomorus niphonius)、中國槍烏賊(Loligo chinensis)、南美白對蝦(Penaeus vanname)絞碎, 添加適量維生素和螺旋藻, 攪拌均勻后黏合成混合鮮餌, 多春魚和混合餌料的營養(yǎng)成分見表1。
1.3 攝食行為與生長和生理觀察
攝食行為觀察項目包括: 潛水員潛入水底喂食前后產(chǎn)后親本的游泳區(qū)域、游泳水層、游泳平穩(wěn)性,攝食積極性, 吞咽餌料行為過程, 攝食量等。各項目的計算方法如下:
游泳區(qū)域: 親本在觀察時段內(nèi)經(jīng)常游動的區(qū)域。區(qū)域劃分方法為, 將馴養(yǎng)池按照長度平均分成6個區(qū)域, 約5 m/區(qū), 順序記為1—6, 3—4為中華鱘的日常水下投食區(qū); 從池底向池水表面垂直分為 4個水層, 1.1 m/層, 從水體底部向表面順序記為底層、下層、中層、上層(接近水體表面記為表層)。在池內(nèi)外設(shè)置區(qū)域和水層劃分記號。
游泳平穩(wěn)性: 通過觀察親本游動時軀體左右搖擺程度及上浮和下潛的平穩(wěn)性確定。平穩(wěn)性好—游動時身體平穩(wěn), 可自如下潛或上浮; 平穩(wěn)性較好—游動時身體左右擺動, 但不明顯, 可自如下潛或上浮; 平穩(wěn)性較差—游動時身體左右擺動較明顯, 下潛或上浮時不僅尾部擺動幅度明顯加大, 身體左右擺動較明顯; 平穩(wěn)性差—游動時身體左右擺動明顯,下潛或上浮時尾部擺動幅度大, 且下潛或上浮困難。
攝食積極性: 通過觀察潛水員投遞餌料時親本口的伸展程度表示。強(qiáng)—口完全伸展, 伸展長度≥10 cm; 較強(qiáng)—口伸展長度約為5—10 cm; 較弱—口伸展長度≤5 cm; 弱—口不主動伸展。
吞咽餌料過程: 觀察描述行為表現(xiàn)和每次喂食時親本吞吐和吞咽餌料的平均用時。
攝食量: 記錄每次所攝食種類和攝食量。
為降低捕撈對產(chǎn)后親本的影響, 從轉(zhuǎn)入馴養(yǎng)池開始, 每 3個月捕撈測定生長參數(shù)和血液參數(shù)。尾靜脈采血 5 mL, 血細(xì)胞參數(shù)的測定用肝素鈉抗凝,其余參數(shù)測定用 3000 r/min離心 20min 后取血清, –4℃保存?zhèn)溆谩8黜椦簠?shù)的測定方法如下:
紅細(xì)胞計數(shù)(Red blood sell, RBC): 以濃度為0.5% NaCl溶液為稀釋液將血樣稀釋 200倍后, 用Neubarner氏計數(shù)板在顯微鏡下計數(shù)。
血紅蛋白量(Hemoglobin, Hb): 采用比色法測定。
紅細(xì)胞比積(Hematocritvalue, Ht): 采用溫氏法測定。
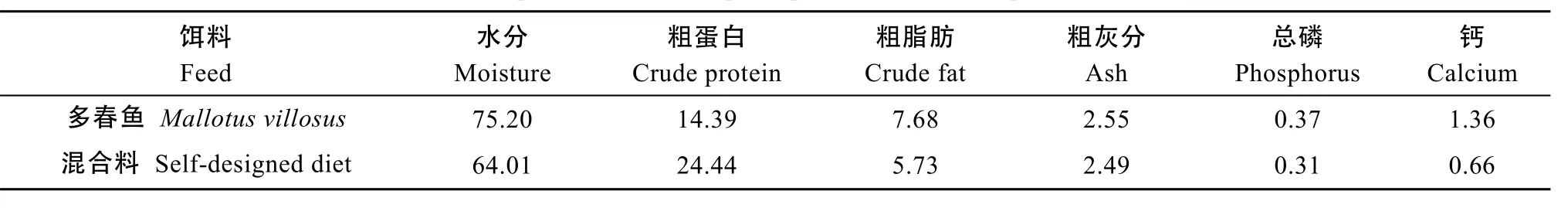
表1 產(chǎn)后親本餌料的營養(yǎng)成分Tab. 1 The composition of feed of post-spawned cultured Acipenser sinensis (%)
實驗測定的19項血清指標(biāo)包括: 血清促甲狀腺激素(Serum thyroid-stimulating hormone, TSH)、血清總T3 (Serum T3, TT3)、血清總T4 (Serum T4, TT4)、血清游離 T3 (Serum free T3, FT3)、血清游離 T4 (Serum free T4, FT4)、鈣(Calcium, Ca2+)、無機(jī)磷(Phosphorus, P3+)、鎂(Magnesium, Mg2+)、丙氨酸氨基轉(zhuǎn)移酶(Glutamate-pyruvate, ALT)、天冬氨酸氨基轉(zhuǎn)移酶(Aspartate aminotransferase, AST)、堿性磷酸酶 (Alkaline phosphatase, ALP)、 膽 堿 酯 酶(Cholinesterase, CHE)、總蛋白(Total protein, TP)、白蛋白(Albumin, ALB)、球蛋白(Globulin, GLB)、甘油三酯(Triglycerides, TG)、膽固醇(Total cholestero, CHO)、血糖(Glucose, GLU)。使用英科新創(chuàng)(廈門)科技有限公司生產(chǎn)的試劑盒在全自動生化分析儀(OLYMPUS AU400)上測定生化指標(biāo), 使用西門子生產(chǎn)的試劑盒在全自動化學(xué)發(fā)光免疫分析儀(ADVIA Centaur CP)上測定激素指標(biāo)。
1.4 數(shù)據(jù)處理與計算公式
體長生長率: (Growth rate of length, GRL): GRL=(l2-l1)/l1
體重生長率: (Growth rate of weight, GRW): GRW=(W2-W1)/W1
式中, l1、l2為單位時間內(nèi)始、末體長(cm); W1、W2為單位時間內(nèi)始、末體重(kg)。
參照組數(shù)據(jù)統(tǒng)計分析用SPSS 13.0軟件完成。用單因素方差分析(ANOVA)參照組血液指標(biāo)差異, 以P<0.05為顯著水平。參照組結(jié)果為平均值±標(biāo)準(zhǔn)差。
2 結(jié)果
2.1 攝食行為和攝食量
在人工催產(chǎn)繁殖后, 采用產(chǎn)前培育期的主要餌料多春魚誘導(dǎo)開口, 產(chǎn)后親本 2周后攝食, 每日僅食少量多春魚, 攝食過程中反復(fù)吞吐餌料, 并從鰓孔溢出大量餌料碎渣。在轉(zhuǎn)入馴養(yǎng)池后(12月28日),潛水員通過多春魚攜帶混合餌料方式伴隨游動, 馴化喂食(攜帶量逐漸增加), 20d后產(chǎn)后親本可直接攝食混合料, 攝食量逐步提高, 攝食過程中吞吐餌料及從鰓孔溢出餌料碎渣行為逐漸減少。雌性親本從4月開始, 攝食期間降至池體下層進(jìn)入喂食區(qū)域?qū)な? 攝食積極性和攝食量明顯提高, 游泳平穩(wěn)性逐漸轉(zhuǎn)好, 于 5月可直接吞咽餌料。雄性攝食行為恢復(fù)快于雌性, 于4月可直接吞咽餌料(表2)。雌性恢復(fù)培養(yǎng)初期攝食期間行為明顯異常, 主要表現(xiàn)為隨吞吐餌料時鰓孔溢出大量氣泡, 身體明顯下沉, 然后沖出水面吸氣調(diào)節(jié)身體平衡。隨著4月后攝食量持續(xù)提高, 此行為明顯減輕至消失。培養(yǎng)后期雌性攝食量增高幅度明顯大于雄性親本。從轉(zhuǎn)入馴養(yǎng)池至培育期末, 攝食量占體質(zhì)量比例為雌性 0.52%—2.04%, 雄性0.39%—1.60%。
2.2 生長效果
雌雄親本的體長和體質(zhì)量產(chǎn)后培育期間變化趨勢相似(圖 1), 至培育期末, 體長分別為 189和192 cm, 體質(zhì)量為55.5和63.5 kg。從轉(zhuǎn)入馴養(yǎng)池到培育期末, 體長增長率分別為5.00%和3.23%, 體質(zhì)量增長率分別為 44.16%和 23.30%。雌、雄參照組在培育期體長和體質(zhì)量的增長率分別為 2.91%、2.65%和15.73%、5.56%, 明顯低于產(chǎn)后雌雄親本。2.3 血清中甲狀腺指標(biāo)和血液理、化指標(biāo)的變化
雌性親本多項血液指標(biāo)培育初期低于參照組平均水平, 經(jīng)產(chǎn)后培育, 15項指標(biāo)可恢復(fù), 其中6項指標(biāo), Ca2+、P3+、AST、CHE、GLU、RBC在較短時間(產(chǎn)后5個月左右)可恢復(fù), 9項指標(biāo)TT4、FT4、ALP、TP、ALB、GLB、CHO、TG、Hb產(chǎn)后較長時間(產(chǎn)后8—11個月左右)恢復(fù)至參照組平均水平。

表2 產(chǎn)后親本在馴養(yǎng)池的攝食行為Tab. 2 The feeding behaviour of post-spawned cultured Acipenser sinensis during the recovering in the aquarium
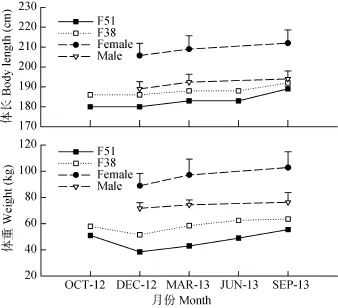
圖1 產(chǎn)后培育期間親本體長和體質(zhì)量的變化Fig. 1 Changes in the body length and the body weight of postspawned cultured Acipenser sinensis during the recovering
4項指標(biāo)培育期末未恢復(fù), TT3、FT3、ALT高于參照組平均值, TT3、FT3近 3倍于參照組平均水平, ALT約1.7倍于參照組平均水平, Ht低于參照組平均值。TSH、Mg、A/G培育期間無明顯變化, 接近參照組平均水平(表3)。雄性親本產(chǎn)后培育期間血液指標(biāo)恢復(fù)與雌性親本有所不同(表 4)。12項指標(biāo)可恢復(fù), Ca2+、AST、GLB、RBC 4項指標(biāo)在較短時間可恢復(fù), FT4、TT4、TP、ALB、A/G、CHO、Ht、Hb 8項指標(biāo)產(chǎn)后較長時間恢復(fù), TT3、FT3變化趨勢與雌性親本較相似, 約2倍于參照組平均值, ALP低于參照組平均值。TSH、Mg2+、P3、ALT、CHE、GLU、TG 7項指標(biāo)培育初期接近參照組平均水平, 以后變化不明顯。與雌性親本相比, 雄性親本培育初期多項指標(biāo)穩(wěn)定, GLB值升高速度快于雌性, 至培育期末, 多數(shù)指標(biāo)值恢復(fù)。這表明至培育期末, 產(chǎn)后親本生理情況基本恢復(fù), 雄性親本在初期恢復(fù)效果優(yōu)于雌性親本。
3 討論
3.1 人工條件下可實現(xiàn)養(yǎng)殖中華鱘產(chǎn)后康復(fù)和繼續(xù)生長
本文產(chǎn)后養(yǎng)殖親本為長江水產(chǎn)研究所 2012年初次全人工繁殖的個體, 產(chǎn)后身體虛弱, 通過本次康復(fù)培養(yǎng)實驗, 逐漸恢復(fù)攝食, 甲狀腺激素分泌水平提高, 血液理化指標(biāo)值趨向于同齡未產(chǎn)組平均水平, 生長優(yōu)于未產(chǎn)組, 雌性異常行為恢復(fù)正常, 說明產(chǎn)后養(yǎng)殖親本可以獲得康復(fù)并繼續(xù)生長, 這也使得中華鱘養(yǎng)殖親本再次成熟后生殖力的提高成為可能, 對養(yǎng)殖中華鱘資源的重復(fù)利用具有重要意義。
3.2 養(yǎng)殖中華鱘產(chǎn)后的攝食行為
在魚類繁殖后, 恢復(fù)攝食的效果和攝入的營養(yǎng)水平是影響親本成活率及再次繁殖周期長短的主要因素[8]。在天然水域中, 產(chǎn)后中華鱘食物可獲得性受到環(huán)境限制, 存活率低, 再次繁殖的間隔時間是2年甚至更長時間[9]; 而蓄養(yǎng)產(chǎn)后野生中華鱘可攝食營養(yǎng)豐富的食物, 存活率高, 產(chǎn)后7—12個月行為恢復(fù),再次發(fā)育的間隔時間多為2—3年[5]。在本研究中, 產(chǎn)后親本攝食欲差, 特別是雌性身體虛弱, 提供營養(yǎng)豐富的適口性飼料并促進(jìn)其快速攝食是康復(fù)成功的關(guān)鍵。雌性親本產(chǎn)后初期攝食期間泳層與游泳區(qū)域不固定, 不能直接游至下層尋食, 且游動平穩(wěn)性差,通過潛水員伴隨游動投遞食物方式, 提高了食物的可獲得性, 降低攝食時的體能消耗。隨體力逐漸加強(qiáng), 攝食積極性增強(qiáng), 攝食期間可降低泳層主動尋食, 攝食量也逐步提高。中華鱘對食物有較長時間記憶[5], 攝取食物后, 主要靠口腔觸覺和味覺判斷適口性, 最終吞入或摒棄食物, 對部分食物的選擇性可以被逐步馴化[4, 10]。在本文中, 先采用產(chǎn)前主要食物多春魚誘導(dǎo)產(chǎn)后親本開口, 然后混合以多春魚為原料的自制鮮餌繼續(xù)馴化, 利于親本快速選食[11]。由于雄性親本產(chǎn)前即攝食混合餌料, 因此適應(yīng)混合料的時間明顯短于雌性。相較于野生產(chǎn)后中華鱘通過灌食促進(jìn)恢復(fù)攝食而言, 養(yǎng)殖中華鱘較容易恢復(fù)攝食, 但采用多春魚馴化攝食混和餌料也需要20d[4],與野生中華鱘灌食到自主攝食的時間相似, 這可能與中華鱘消化有關(guān)的酶類及消化有關(guān)的神經(jīng)內(nèi)分泌系統(tǒng)需要一段時間平衡有關(guān)[8]。另外, 野生中華鱘主動攝食后泳層即下降, 雌性親本則遠(yuǎn)長于野生, 可能受其虛弱體質(zhì)影響所致[4, 5]。混和餌料的營養(yǎng)均衡性優(yōu)于多春魚, 利于促進(jìn)產(chǎn)后親本快速康復(fù)。
行為是動物對外界環(huán)境和內(nèi)部環(huán)境變化的外在反應(yīng)。本文雌性親本產(chǎn)后初期攝食行為明顯異常。分析認(rèn)為, 一方面, 養(yǎng)殖個體類似于天然水域野生個體, 在性腺發(fā)育過程中停食或少食, 依靠體內(nèi)的脂肪完成性腺的最后成熟[9], 但是由于缺少野生中華鱘繁殖洄游前的育肥階段, 性腺發(fā)育過程又要消耗大量營養(yǎng)物質(zhì)(特別是雌性), 致使產(chǎn)后體內(nèi)營養(yǎng)物質(zhì)積累不足, 易導(dǎo)致體質(zhì)虛弱。其次, 大型雌性動
物一般在人工繁殖過程中受到的損傷和應(yīng)激大于雄性, 這也容易導(dǎo)致產(chǎn)后胃腸神經(jīng)紊亂和虛弱, 致厭食甚至死亡等[12]。中華鱘靠氣鰾保持浮力, 氣鰾開口于食道后段, 健康個體可以將吞入的空氣通過鰾管送入鰾內(nèi), 也可以通過氣鰾周圍肌肉的收縮經(jīng)鰾管把空氣排出以調(diào)節(jié)浮力[13]。雌性親本產(chǎn)后體弱,可能胃腸神經(jīng)紊亂, 吞吐餌料時食道和氣鰾周圍肌肉不能協(xié)調(diào)運動, 攝食時浮力控制出現(xiàn)異常。隨攝食逐漸恢復(fù), 攝入營養(yǎng)量提高, 雌性親本產(chǎn)后7個月體力明顯增強(qiáng), 行為恢復(fù)正常, 與蓄養(yǎng)野生產(chǎn)后中華鱘行為恢復(fù)正常的時間相似[5]。這說明, 在人工養(yǎng)殖條件下, 營養(yǎng)豐富的適口餌料和促進(jìn)攝食的方法可以提高養(yǎng)殖親本(特別是雌性)的產(chǎn)后恢復(fù)效果,這也是本文研究獲得成功的關(guān)鍵。本文混合餌料僅是初步嘗試, 深入探討親本在性腺進(jìn)入快速發(fā)育階段前的營養(yǎng)強(qiáng)化方式, 研究產(chǎn)后恢復(fù)培養(yǎng)時具有誘食效果的強(qiáng)化餌料, 可以成為中華鱘親魚培育技術(shù)新的研究方向。
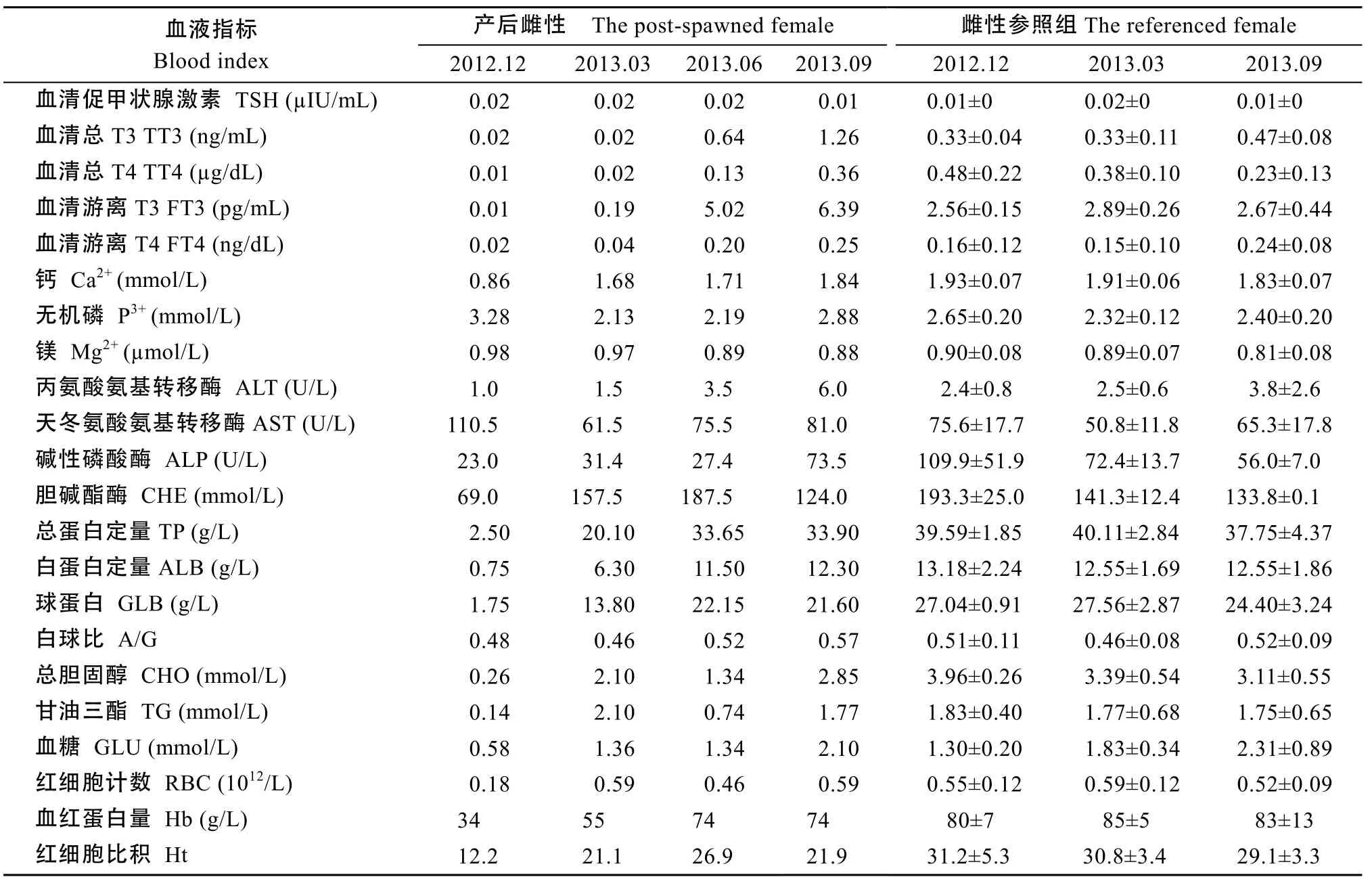
表3 雌性親本產(chǎn)后培育期間血液指標(biāo)與參照組比較Tab. 3 Comparison of the blood indices between the post-spawned female and the referenced female during recovery (n=4)
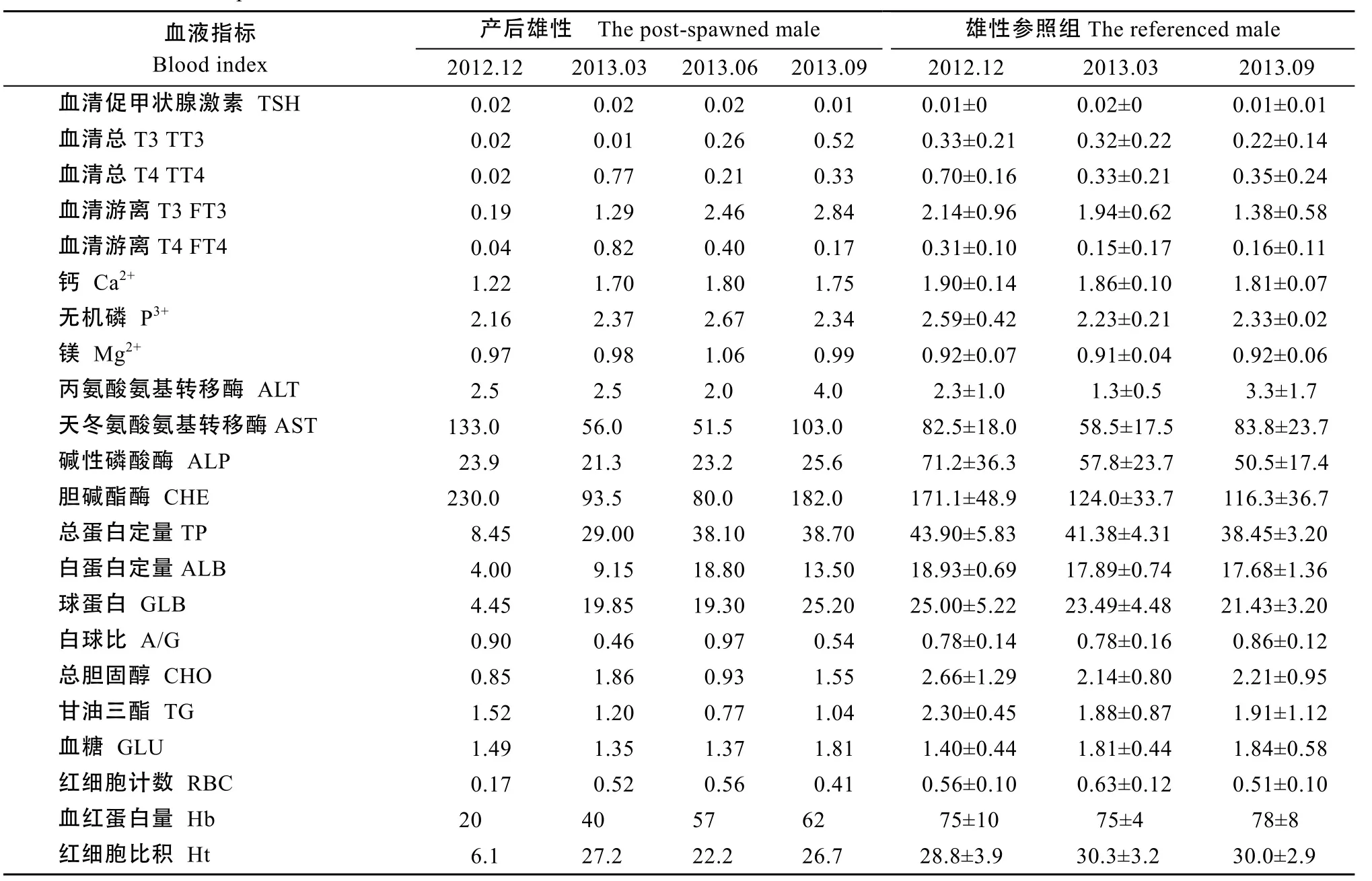
表4 雄性親本產(chǎn)后培育期間血液指標(biāo)與參照組比較Tab. 4 Comparison of the blood indices between the post-spawned male and the referenced male during recovery (n=4)
3.3 初次繁殖的養(yǎng)殖中華鱘產(chǎn)后生長特點
危起偉等[2]研究了初次性成熟的養(yǎng)殖中華鱘生物學(xué)情況, 發(fā)現(xiàn)雌性年齡小于記錄到的最小野生雌性中華鱘性成熟年齡, 性成熟體長和體質(zhì)量值遠(yuǎn)小于長江野生雌性最小成熟個體值, 認(rèn)為這不利于中華鱘的保護(hù)和規(guī)模化培育。本文親本出現(xiàn)產(chǎn)后體長繼續(xù)生長的特點, 與天然水域產(chǎn)后親本可持續(xù)生長的特點相同[9]。養(yǎng)殖親本生長速度均優(yōu)于未進(jìn)入性腺快速發(fā)育的同齡未產(chǎn)組, 這可能是長期饑餓后攝入豐富的食物時出現(xiàn)的補(bǔ)償生長現(xiàn)象[13]。另外, 水深、空闊、水質(zhì)清新的大型養(yǎng)殖空間有利于大體型成體中華鱘的生長[14]。這說明, 在人工條件下, 通過加強(qiáng)產(chǎn)后培養(yǎng), 可以提高親本的生長率, 這使得初次繁殖親本個體小的情況得以改善成為可能, 對于提高再次成熟后的生殖力具有現(xiàn)實意義。
3.4 產(chǎn)后親本的血清甲狀腺指標(biāo)、血液理化指標(biāo)與攝食、健康情況及生長性能的關(guān)系
魚類的甲狀腺素主要是T3 和T4 [T3是由T4在外周組織中(主要是肝臟)脫碘轉(zhuǎn)化而來的], 對代謝活動、生長、行為等有重要作用, T3的生物活性比 T4強(qiáng)[15]。饑餓會影響下丘腦—垂體—甲狀腺軸,使甲狀腺對TSH刺激的敏感性降低, T4向T3的轉(zhuǎn)化率下降[16]。在本文培養(yǎng)期間, 產(chǎn)后親本TSH持平于參照組, 而食物攝取量提高后甲狀腺激素水平明顯升高, 表明食物攝取量對甲狀腺激素的分泌影響較大。這與對黑鯛(Sparus macroehhalus)、虹鱒(Oncorhynchus mykiss)等研究結(jié)果較為一致[17]。食物的營養(yǎng)成分(特別是蛋白質(zhì))是影響甲狀腺激素分泌的重要因子[18], 本文產(chǎn)后培養(yǎng)采用的主要飼料中蛋白質(zhì)含量較高, 可能也更有利于促進(jìn)甲狀腺激素的產(chǎn)生。雄性親本產(chǎn)后初期TT3、FT3、TT4水平和體質(zhì)量增長率大于雌性, 而6月以后, 雌性TT3、FT3水平和體質(zhì)量增長率明顯高于雄性。另外, 相比于參照組, 雌、雄產(chǎn)后親本實驗?zāi)┢?TT3、FT3和雌性體長、體質(zhì)量及雄性體長均處于較高水平, 說明了甲狀腺激素與生長關(guān)系密切。這在其他魚的生長研究中也有類似報道[17, 18]。
RBC與Ht培養(yǎng)期間快速增高, Hb升高速度滯后于紅細(xì)胞增生的速度。魚類紅細(xì)胞由脾臟等造血器官產(chǎn)生, 主要有攜帶和運輸氧氣及排除二氧化碳的功能。隨攝食逐漸恢復(fù), 紅細(xì)胞快速增生, 說明紅細(xì)胞生成過程受營養(yǎng)水平影響大, RBC值增高顯示機(jī)體代謝活動增強(qiáng)。
產(chǎn)后親本Ca2+、Mg2+、P3+3月恢復(fù)至參照組平均水平。Ca2+、P3+在骨骼形成和維持酸堿平衡等代謝中起重要作用, Mg2+除了參與骨鹽形成外, 還是很多酶的激活劑。3種離子含量快速恢復(fù), 表明隨著恢復(fù)攝食, 產(chǎn)后親本可通過自身的調(diào)節(jié)快速維持內(nèi)環(huán)境的穩(wěn)定。較雄性親本相比, 雌性的 Ca2+及鈣磷比值在12月時均明顯低于參照組水平, 這可能受到卵巢發(fā)育過程中卵黃蛋白原合成的影響所致[19]。
ALT主要存在于肝細(xì)胞胞漿中, 肝細(xì)胞受損,細(xì)胞通透性增強(qiáng), 血清ALT活性上升。AST也存在于肝臟, 也可反映肝臟的生理狀態(tài)。CHE主要由肝臟合成, 故能靈敏地評價肝細(xì)胞的合成能力, 與ALT、AST可做為指示肝臟功能的酶學(xué)指標(biāo)[20, 21]。本文產(chǎn)后親本3月時ALT、AST、CHE接近參照組平均水平, 說明肝臟的的代謝活動已接近健康的未產(chǎn)個體。培育末期, 雌性產(chǎn)后親本ALT增高至參照組1.7倍的水平, 這主要與其后期快速生長, 甲狀腺功能亢進(jìn)有關(guān)[22]。ALP主要來源于肝臟、腎臟、成骨細(xì)胞等臟器和組織, 是反映成骨細(xì)胞活性、骨生成狀況和鈣、磷代謝及腎臟功能的重要生化指標(biāo),甲狀腺激素影響ALP活性[22, 23]。雌性親本培育末期ALP快速升高, 與甲狀腺激素分泌水平和繼續(xù)加速生長趨勢一致。另外, ALP值接近參照組平均水平也說明腎臟功能逐漸恢復(fù)。雄性ALP在恢復(fù)培養(yǎng)期間較穩(wěn)定, 至培育末期略有提高, 仍然低于參照組平均水平。這與其體長增長6月后才快于參照組較為一致。
GLU是機(jī)體組織生化活動所需能量的來源, CHO、TG是脂肪的代謝產(chǎn)物, 與TP同受營養(yǎng)水平的影響顯著[24, 25]。產(chǎn)后親本 TP、ALB、GLB值隨攝食量提高持續(xù)增高, 表明受營養(yǎng)水平影響明顯,與對施氏鱘(Acipenser schrenckii)[26]的研究結(jié)果較為一致。魚的生長主要是依靠蛋白質(zhì)在體內(nèi)構(gòu)成組織和器官, 攝入蛋白質(zhì)總量提高后, 機(jī)體蛋白質(zhì)代謝增強(qiáng), 合成速率提高, 因此TP、ALB、GLB與體質(zhì)量表現(xiàn)出一致的增長趨勢, 這也說明TP、ALB、GLB能較好地反映機(jī)體的營養(yǎng)水平。雌性親本TP、ALB、GLB值 12月遠(yuǎn)低于參照組, TG、CHO、GLU也有類似表現(xiàn), 而雄性相對差別較小, 表明雌性性腺發(fā)育和繁殖過程消耗的營養(yǎng)和能量物質(zhì)多, 易造成產(chǎn)后體內(nèi)營養(yǎng)積累不足, 導(dǎo)致體質(zhì)虛弱。這也提示,加強(qiáng)產(chǎn)后營養(yǎng)護(hù)理是促進(jìn)雌性親本康復(fù)的關(guān)鍵。
綜上所述, 本次養(yǎng)殖親本產(chǎn)后康復(fù)培養(yǎng)初次獲得成功, 盡管親本獲得較好的康復(fù)和生長效果, 然而由于本研究主要依據(jù) 2012年全人工繁殖的產(chǎn)后親魚恢復(fù)培養(yǎng)實驗數(shù)據(jù)得來, 受研究樣本量限制,尚存在偶然因素影響, 產(chǎn)后初期數(shù)據(jù)完整性也存在不足, 還需繼續(xù)增加實驗樣本, 積累多批次恢復(fù)培養(yǎng)數(shù)據(jù)。
[1] Wang J H, Wei Q W, Zou Y C. Conservation strategies for the Chinese sturgeon, Acipenser sinensis, an overview on 30 years of practices and future needs [J]. Journal of Applied Ichthyology, 2011, 27(2): 176—180
[2] Wei Q W, Li L X, Du H, et al. Research on technology for controlled propagation of cultured Chinese sturgeon (Acipenser sinensis) [J]. Journal of Fishery Science of China, 2013, 20(1): 1—11 [危起偉, 李羅新, 杜浩, 等. 中華鱘全人工繁殖技術(shù)研究. 中國水產(chǎn)科學(xué), 2013, 20(1): 1—11]
[3] Webb M A, Doroshov S I. Importance of environmental endocrinology in fisheries management and aquaculture of sturgeons [J]. General and Comparative Endocrinology, 2011, 170(2): 313—321
[4] Zhang X Y, Liu J Y, Wei Q W, et al. Recovery of foraging ability of wild female Chinese sturgeon after artificial propagation in aquarium by different feeding methods [J]. Chinese Journal of Zoology, 2007, 42(2): 142—146 [張曉雁,劉鑒毅, 危起偉, 等. 不同喂食方法對野生產(chǎn)后中華鱘的攝食促進(jìn). 動物學(xué)雜志, 2007, 42(2): 142—146]
[5] Wei Q W, Zhang X Y, Zhang X F, et al. Acclimating and maintaining Chinese sturgeon Acipenser sinensis in a large public aquarium environment [J]. Journal of Applied Ichthyology, 2011, 27(2): 533—540
[6] Guo B F, Chang J B, Xiao H, et al. The productive biology of first filial generation of Acipenser sinensis growing up in the freshwater environment [J]. Acta Hydrobiologica Sinica, 2011, 35(6): 940—945 [郭柏福, 常劍波, 肖慧, 等. 中華鱘初次全人工繁殖的特性研究. 水生生物學(xué)報, 2011, 35(6): 940—945]
[7] Zhang X Y. Blood biochemistry and its applications in Chinese sturgeon (Acipenser sinensis) [D]. Thesis for Master of Science. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan. 2012 [張曉雁. 中華鱘血液生化學(xué)的研究與應(yīng)用. 博士學(xué)位論文, 中國科學(xué)院水生生物研究所,武漢. 2012]
[8] Liu L, Zhang J M, Guo F, et al. Effects of renewal cultivation and renewal reproduction after spawning in Japanese eel Anguilla japonica [J]. Journal of Fishery Science of China, 2004, 11(1): 54—58 [柳凌, 張潔明, 郭峰, 等. 日本鰻鱺人工催產(chǎn)后親魚恢復(fù)培養(yǎng)與再催產(chǎn)效果. 中國水產(chǎn)科學(xué), 2004, 11(1): 54—58]
[9] Chen X H. Biology and Resources Status of Acipenseriformes [M]. Beijing: Ocean Press. 2007, 96—99 [陳細(xì)華.鱘形目魚類生物學(xué)與資源現(xiàn)狀. 北京: 海洋出版社. 2007, 96—99]
[10] Zhuang P, Zhang L Z, Luo G, et al. Function of sense organs to the feeding behavior of juveniles Chinese sturgeon captured from the Yangtze estuary [J]. Acta Hydrobiologica Sinica, 2008, 32(4): 475—481 [莊平, 章龍珍, 羅剛, 等.長江口中華鱘幼魚感覺器官在攝食行為中的作用. 水生生物學(xué)報, 2008, 32(4): 475—481]
[11] Kasumyan A Q. Sturgeon food searching behaviour evoked by chemical stimuli: a reliable sensory mechanism [J]. Journal of Applied Ichthyology, 2002, 18(4—6): 685—690
[12] Yu Y Q. Prevention methods of anorexia in postpartum sows [J]. Shandong Journal of Animal Science and Veterinary Medicine, 2012, 33(22): 34—35 [于彥強(qiáng). 母豬產(chǎn)后不食癥的綜合防治方法. 山東畜牧獸醫(yī), 2012, 33(22): 34—35]
[13] Xie C X. Ichthyology [M]. Beijing: China Agriculture Press. 2010, 304 [謝從新. 魚類學(xué). 北京: 中國農(nóng)業(yè)出版社. 2010, 304]
[14] Yang D G, Wei Q W, Wang K, et al. Three types of culture Chinese sturgeon (Acipenser sinensis) and analysis of their effects by comparison [J]. South China Fisheries Sciences, 2006, 2(2): 1—5 [楊德國, 危起偉, 王凱, 等. 淡水養(yǎng)殖中華鱘成魚的三種方式及效果比較. 南方水產(chǎn), 2006, 2(2): 1—5]
[15] Lin H R. Fish Physiology [M]. Guangzhou: Guangzhou Higher Education Press. 2004, 222 [林浩然. 魚類生理學(xué).廣州: 廣州高等教育出版社. 2004, 222]
[16] Pedro N D, Delgado M J, Gancedo B, et al. Changes in glucose, glycogen, thyroid activity and hypothalamic catecholamines in tench by starvation and refeeding [J]. Journal of Comparative Physiology B, 2003, 173(6): 475—481
[17] Deng L, Zhang W M, Lin H R. Effects of starvation on the contents of growth hormone and triiodothyronine in serum, and lipid and protein of white muscles and livers in black seabream [J]. Zoological Research, 2003, 24(2): 94—98 [鄧?yán)? 張為民, 林浩然. 饑餓對黑鯛血清生長激素、甲狀腺激素以及白肌和肝臟脂肪、蛋白質(zhì)含量的影響. 動物學(xué)研究, 2003, 24(2): 94—98]
[18] Wang G Q, Zhou G Q, Chen J M, et al. Effect of dietary protein on growth and hormone of cultural burnus juveniles [J]. Acta Hydrobiologica Sinica, 2008, 32(4): 543—550 [王桂芹,周洪琪, 陳建明, 等. 鲌飼料蛋白對翹嘴紅 生長和內(nèi)分泌激素的影響. 水生生物學(xué)報, 2008, 32(4): 543—550]
[19] Matsche M A, Rosemary K M, Brundage H M. Hematology and plasma chemistry of wild shortnose sturgeon Acipenser brevirostrum from Delawarw River, USA [J]. Journal of Applied Ichthyology, 2013, 29(1): 6—14
[20] Zhong Q J, Zhang S F. The Analysis Technique of Biochemistry [M]. Wuhan: Huazhong University of Science and Technology Press. 2012, 196—200 [仲其軍, 張淑芳.有機(jī)化學(xué)檢驗技術(shù). 武漢: 華中科技大學(xué)出版社. 2012, 196—200]
[21] Wang Z D, Yang J J, Li L A, et al. Effects of different intensities of swimming stress on serum ALT, AST and AIP activities in mice [J]. Heilongjiang Animal Science and Veterinary Medicine, 2013, (11): 164—166 [王轉(zhuǎn)丁, 楊晶晶, 李留安, 等. 不同強(qiáng)度游泳應(yīng)激對小鼠血清 ALT、AST和 ALP活性的影響. 黑龍江畜牧獸醫(yī), 2013, (11): 164—166]
[22] Su H X, Zhong Y H, Gao B H. Detection and clinical value of liver function for the patients with hyperthyroidism [J]. Journal of Guangxi Medical University, 2002, 22(1): 91—92[蘇會璇, 鐘宇華, 高碧華. 甲狀腺功能亢進(jìn)癥者肝功能的檢測及其臨床意義. 廣西醫(yī)科大學(xué)學(xué)報, 2002, 22(1): 91—92]
[23] Wang S Y, Yan S M. Research progress of alkaline phosphatase in bone metabolism of animal [J]. Feed Review, 2009, (4): 14—16 [王石瑩, 閆素梅. 堿性磷酸酶在動物骨骼代謝中的研究進(jìn)展. 飼料博覽, 2009, (4): 14—16]
[24] Qiang J, Yang H, Wang H, et al. Effects of different dietary protein levels on serum biochemical indices and expression of liver HSP70 mRNA in gift tilapia (Oreochromis niloticus) under low temperature stress [J]. Acta Hydrobiologica Sinica, 2013, 37(3): 434—443 [強(qiáng)俊, 楊弘, 王輝, 等. 飼料蛋白水平對低溫應(yīng)激下吉富羅非魚血清生化指標(biāo)和HSP70 mRNA表達(dá)的影響. 水生生物學(xué)報, 2013, 37(3): 434—443]
[25] De Pedro N, Guijarro A I, Lpez-patio M A, et al. Daily and seasonal variations in hematological and blood biochemical parameters in the tench, Tinva tinca Linnaeus, 1758 [J]. Aquaculture Research, 2005, 36(12): 1185—1196
[26] Zhang L Z, Zhuang P, Zhang T, et al. Hematological biochemical norm of different age groups of cultured Amur sturgeon (Acipenser schrenckii) [J]. Journal of Fisheries of China, 2007, 31(2): 159—164 [章龍珍, 莊平, 張濤, 等. 人工養(yǎng)殖不同年齡施氏鱘的血液生化指標(biāo). 水產(chǎn)學(xué)報, 2007, 31(2): 159—164]
THE POST-SPAWNED RECOVERY OF CULTURED CHINESE STURGEON ACIPENSER SINENSIS
ZHANG Xiao-Yan1, 2, DU Hao1, WEI Qi-Wei1, ZHANG Yan-Zhen2, WANG Yan-Peng2, CAI Jing-Jiang2,
LIU Zhi-Gang1, QIAO Xin-Mei1and XIONG Wei1
(1. Key Laboratory of Freshwater Biodiversity Conservation, Ministry of Agriculture of China, Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Wuhan 430223, China; 2. Beijing Xinwoda Marine Technology Co. Ltd (Beijing Aquarium), Beijing 100081, China)
In this study, we recorded the feeding behavior, the growth indicators and the blood indices for 11 months after the spawning of cultured Chinese sturgeon. The growth indicators and the blood samples of the 2 referenced groups were also analyzed, and these 2 groups were in the slow gonad development stage and the same age as the female and male post-spawned individuals. We tested 22 blood indices that included red blood cells (RBC), hematocritvalue (Ht), Hemoglobin (Hb), serum calcium (Ca2+), phosphorus (P3+), magnesium (Mg2+), glutamate-pyruvate (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), cholinesterase (CHE), magnesium (Mg), total protein (TP), albumin (ALB), globulin (GLB), albumin/ globulin (A/G), total cholestero (CHO), triglycerides (TG), glucose (GLU), serum thyroid-stimulating hormone (TSH), serum T3 (TT3), serum T4 (TT4), serum free T3 (FT3), and serum free T4 (FT3). We found that the re-feeding of the post-spawned sturgeons can be induced with Mallotus villosus, the preferred feed of the two sturgeons before the controlled reproduction. Then the sturgeons started to feed on self-designed diet composed of Mallotus villosus, Scomberomorus niphonius, Loligo chinensis and Penaeus vanname. The feeding of the post-spawned female was initially abnormal due to the weakness, and then the feeding quantity increased to 2.04% of the body weight (to 1.60% of body weight in the post-spawned male at the same time) and the feeding turned back to normal 7 months post-spawning. As the feeding quantity increased over 9 months of recovery, the post-spawned cultured sturgeons showed the higher growth rates of the body length (5.00% and 3.23% for the female and the male respectively) and the body weight (44.16% and 23.30% for the female and the male respectively) than those of the two referenced groups. The concentrations of TT3, TT4, FT3, FT4 and many other blood indices of the two post-spawned sturgeons were lower than those of the two referenced groups in the early recovery, then most of the indices increased nearly to the average levels in the referenced groups, and the levels of TT3 and FT3 were much higher than the average values in the two referenced groups. These implied that the two post-spawned sturgeons were recovered. Our data suggested that refined culture technique could effectively improve the recovery and elevate the growth rate of the post-spawned cultured sturgeon (especially for the female). Our study is also crucial for the reuse of cultured Chinese sturgeon resources and for improving the fecundity after re-maturation.
Acipenser sinensis; After-spawning; Feeding behavior; Growth; Serum thyroid indicators; Blood cell indices; Serum biochemical indices
S965.2
A
1000-3207(2015)04-0705-09
10.7541/2015.93
2014-06-09;
2014-11-20
公益性行業(yè)(農(nóng)業(yè))科技專項(201203086); 國家自然科學(xué)基金項目(31172413); 973計劃課題(2015CB150702)資助
張曉雁(1973— ), 女, 山東棲霞人; 高級工程師, 博士; 主要從事瀕危魚類生理生態(tài)學(xué)和水族館生態(tài)學(xué)方向的研究。E-mail: zn7@sina.com
危起偉(1960—), 男, 江西南昌人; 研究員, 博導(dǎo); 主要研究方向為珍稀魚類保護(hù)。E-mail: weiqw@yfi.ac.cn
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫(yī)科大學(xué)學(xué)報(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結(jié)合肝病雜志(2020年2期)2020-10-27 02:18:50
少年文藝·開心閱讀作文(2018年4期)2018-05-02 07:08:56
科學(xué)大眾(中學(xué))(2018年2期)2018-02-01 12:53:52
少年文藝·開心閱讀作文(2018年1期)2018-01-19 09:05:26
小獼猴學(xué)習(xí)畫刊(2017年12期)2017-12-26 12:08:34
中國火炬(2014年3期)2014-07-24 14:44:39
