PI3K/Akt抑制劑對雞朊蛋白過表達DF-1細胞增殖、侵襲和凋亡的影響
2015-03-07 08:05:20萬學瑞朱曼玲楊潤霞劉桂林
畜牧獸醫學報 2015年10期
關鍵詞:檢測
萬學瑞,朱曼玲,楊潤霞,劉桂林,劉 磊,吳 潤
(甘肅農業大學動物醫學院,蘭州730070)
PI3K/Akt信號轉導通路廣泛存在于細胞中,Akt是此通路中的一個關鍵分子,活性主要由PI3K的產物調控。胞外信號可以通過激活受體酪氨酸激酶(RTK)或G蛋白偶聯受體而激活PI3K,其產物使Akt磷酸化,p-Akt可在細胞質或細胞核內磷酸化多種蛋白質分子,通過改變下游分子的磷酸化狀態參與細胞的生長、增殖和存活。PI3K/Akt信號轉導通路是抑制細胞凋亡,促進細胞生長增殖的重要因素,使細胞維持周期運行[1]。渥曼青霉素(wortmannin,WM)是絲狀真菌繩狀青霉(Penicillium funiculosum)的代謝產物,可特異性、不可逆的抑制PI3K活性,阻斷PI3K/Akt信號通路。
細胞型朊蛋白(cellular prion protein,Pr PC)是由細胞自身基因編碼,普遍表達在真核細胞膜上的GPI錨定蛋白[2]。哺乳動物細胞型朊蛋白錯誤折疊形成異常癢病型朊蛋白(scrapie prion protein,Pr PSC)可致傳染性海綿狀腦病,但在非哺乳動物體內卻未見發病[3]。多項研究已證明哺乳動物細胞型朊蛋白具有多種生理功能,參與調節細胞增殖、分化、凋亡、侵襲、黏附、信號轉導、銅離子代謝和氧化應激等過程[4-10],但具體機制還不清楚。本課題組前期的研究發現雞細胞型朊蛋白(ChPr PC)表達及分布規律同哺乳動物Pr PC相似[11],通過構建雞細胞型朊蛋白過表達的雞成纖維(DF-1)穩定細胞系(DF-1-PrP)證實其可促進DF-1細胞黏附、增殖和侵襲,抑制其凋亡[12],進一步應用實時熒光定量PCR檢測發現雞細胞型朊蛋白過表達DF-1細胞的Akt基因表達量明顯高于DF-1細胞,推測雞細胞型朊蛋白可能參與Akt的激活,進而促進細胞生長與抑制細胞凋亡[13]。為了證實這一推測,本研究擬利用渥曼青霉素阻斷PI3K/Akt信號通路,探討其對雞細胞型朊蛋白過表達DF-1細胞黏附、增殖、侵襲和凋亡的影響及其可能的作用機制。
1 材料與方法
1.1 材料
1.1.1 細胞 DF-1細胞、DF-1-Pr P細胞(Ch Pr PC過表達DF-1細胞)和DF-1-NC細胞(導入空表達載體pCDNA3.1的DF-1細胞),均由本實驗室保存或構建。
1.1.2 主要試劑 胎牛血清(FBS,Hy Clone公司),鼠尾膠原(杭州生友生物技術有限公司);G418、DMEM、無血清培養基(GIBCO公司),渥曼青霉素(Alexis公司),噻唑藍(MTT)、臺盼藍(上海酶聯生化試劑有限公司),Annexin V-FICT/PI雙染細胞凋亡檢測試劑盒(上海貝博生物公司),Transwell小室(Costar公司),牛血清白蛋白(BSA,西安依科生物技術有限公司),其他試劑均為國產分析純。
1.1.3 主要儀器 MyCyclerTMThermal Cycler EN-61010 PCR儀(美國BIO-RDA公司),Light Cycler 480Ⅱ熒光定量PCR儀(美國Roche公司);酶標儀680(美國BIO-RAD公司),FACSCalibur flow cytometer(美國BD公司)等。
1.2 方法
1.2.1 細胞培養 DF-1-PrP、DF-1-NC和DF-1細胞用含10%胎牛血清、青霉素(100 U·m L-1)和鏈霉素(100 U·m L-1)的DMEM培養基置于37℃、5%CO2培養箱培養,當培養瓶中的細胞覆蓋率達到80%以上時,0.25%胰蛋白酶消化傳代,傳3代穩定后進行后續試驗。
1.2.2 黏附試驗 將鼠尾膠原用6 mmol·L-1無菌乙酸配制成6.25 mg·L-1的溶液,50 μL·孔-1加入96孔培養板,4℃過夜制備基底膜,吸出孔中殘余液體,加入50μL無血清培養基37℃孵育30 min水化基底膜。將對數生長期的DF-1-Pr P、DF-1-NC和DF-1細胞,胰蛋白酶消化后,用培養液重懸細胞,調整細胞數為1×105·m L-1,100 μL·孔-1分別接種于包被鼠尾膠原的孔中,再加入渥曼青霉素使其終濃度分別為0、10、20、50、100、200 nmol·L-1,每組4個重復,37℃5%CO2環境下培養細胞,以包被牛血清白蛋白(BSA)為對照。細胞培養1 h后,用PBS溶液小心的洗細胞3次,按照MTT比色法測定培養板各孔的光吸收值(A值),試驗重復3次。應用如下公式分別計算各組細胞的黏附率:黏附率(%)=[(處理組A值/BSA對照組A值)-1]×100%。
1.2.3 侵襲試驗 用6.25 mg·L-1鼠尾膠原溶液包被Transwell小室,4℃過夜風干,吸出殘余液體,加入50μL無血清培養基,37℃孵育30 min。用0.25%的胰蛋白酶消化處于對數生長期的DF-1-Pr P、DF-1-NC和DF-1細胞,無血清培養液重懸細胞(1×105·m L-1)。在下室中加入400μL按1∶1混合的完全培養液和條件培養液(DF-1細胞生長至80%以上融合,換無血清培養液培養24 h后收集培養上清液,過濾除菌),上室中加入100μL細胞懸液,再加入渥曼青霉素使其終濃度分別為0、10、20、50、100、200 nmol·L-1,每組4個重復,37℃5%的CO2環境中培養24 h后,棄培養基,擦凈上室細胞,取出Transwell用PBS洗2次,95%的乙醇溶液固定細胞15 min,風干;加4 g·L-1的臺盼藍溶液染色20 min,PBS洗3次,倒置顯微鏡下隨機計數10個視野的細胞,取平均數,試驗重復3次。
1.2.4 MTT法檢測細胞增殖 取對數生長期的DF-1-PrP、DF-1-NC和DF-1細胞(1× 105·m L-1),100μL·孔-1接種96孔培養板中,加入渥曼青霉素使其終濃度分別為0、10、20、50、100、200 nmol·L-1,以DMEM培養液為空白對照,每組3個重復。細胞培養12、24、36、48、60 h后,每孔加入20μL MTT(5 mg·m L-1)溶液,置于CO2培養箱中孵育4 h后吸出孔內液體,再加入150μL DMSO振蕩10 min。用酶標儀檢測,讀取OD490nm處的吸光值。
1.2.5 流式細胞儀檢測細胞凋亡 在對數生長期的DF-1-PrP、DF-1-NC和DF-1細胞中加入渥曼青霉素使其終濃度分別為0、10、20、50、100、200 nmol·L-1,作用24 h后,消化收集細胞(1× 107·m L-1),按Annexin V-FICT/PI雙染細胞凋亡檢測試劑盒說明書進行操作,冷PBS洗細胞2次,加入200μL結合緩沖液重懸細胞,再加入10 μL Annexin V-FITC和5μL PI,輕搖混勻,孵育15 min;上述溶液中加入300μL結合緩沖液充分混勻,立即進行流式細胞儀檢測。
1.2.6 不同渥曼青霉素濃度下PRNP基因的轉錄量檢測 在對數生長期的DF-1-Pr P、DF-1-NC和DF-1細胞中加入渥曼青霉素使其終濃度分別為0、10、20、50、100 nmol·L-1,作用24 h后,收集細胞提取總RNA。利用本實驗室建立的雞PrPC基因mRNA定量RT-PCR檢測方法,檢測PRNP基因mRNA表達量[13]。
1.2.7 統計學分析 采用SPSS17.0軟件One-Way分析(Anova)或t檢驗進行統計學處理,數據以±s表示,P<0.05,P<0.01為差異有統計學意義。
2 結 果
2.1 渥曼青霉素以劑量依賴方式抑制DF-1-PrP細胞黏附
細胞黏附試驗結果如表1所示,渥曼青霉素濃度為0 nmol·L-1時,各細胞黏附率均最高,且DF-1-Pr P細胞黏附率比DF-1-NC細胞和DF-1細胞分別高26.04%和26.18%,差異極顯著(P<0.01),隨著渥曼青霉素濃度的升高,各細胞的黏附率均顯著下降。渥曼青霉素濃度10~50 nmol·L-1時,DF-1-Pr P細胞黏附率極顯著(P<0.01)高于DF-1和DF-1-NC細胞;100 nmol·L-1時,DF-1-Pr P細胞黏附率顯著(P<0.05)高于DF-1和DF-1-NC細胞;渥曼青霉素濃度達到200 nmol·L-1時,DF-1-PrP、DF-1-NC和DF-1細胞的黏附率無顯著差異;當渥曼青霉素濃度200 nmol·L-1時,3種細胞的黏附率分別被抑制了87.46%、72.65%和84.32%。
2.2 渥曼青霉素以劑量依賴方式抑制DF-1-Pr P細胞侵襲
細胞侵襲試驗結果如表2所示,渥曼青霉素濃度為0 nmol·L-1時,各細胞的侵襲穿膜細胞數最多,其侵襲力最強,隨著渥曼青霉素濃度的升高,細胞的侵襲力顯著下降,但同一渥曼青霉素濃度下DF-1-Pr P細胞侵襲穿膜細胞數顯著高于DF-1-NC和DF-1細胞(P<0.05或P<0.01)。當渥曼青霉素濃度高于100 nmol·L-1時,DF-1-Pr P、DF-1-NC和DF-1細胞均不見侵襲穿膜細胞。
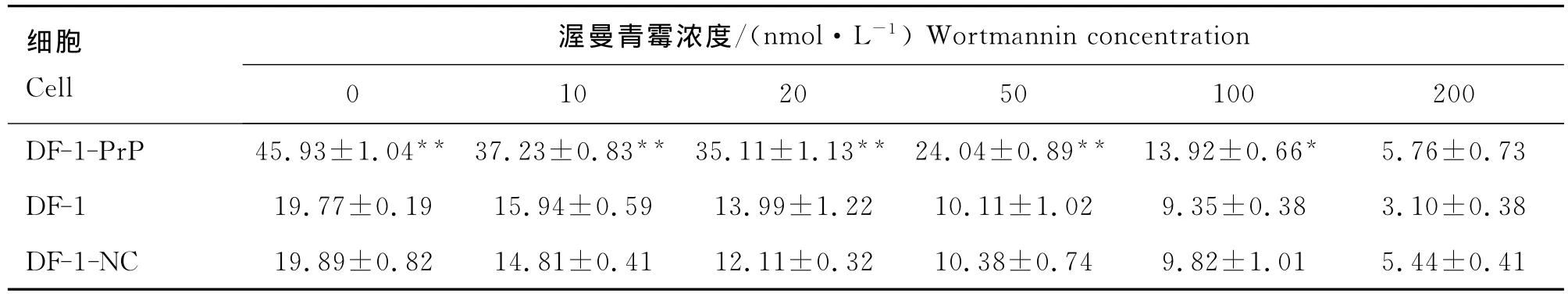
表1 不同濃度渥曼青霉素對細胞黏附率的影響Table 1 Effects of different concentration wortmannin(WM)on cell adhesion rates %

表2 不同濃度渥曼青霉素對細胞侵襲(侵襲細胞數)的影響Table 2 Effects of different concentration wortmannin on cell invasion(Cell numbers of invasion)
2.3 渥曼青霉素以劑量和時間依賴方式抑制DF-1-Pr P細胞增殖
應用MTT法檢測細胞的增殖,結果顯示(圖1),不加渥曼青霉素時,DF-1-Pr P、DF-1-NC和DF-1細胞增殖活性均高于其渥曼青霉素處理的增殖活性;隨著渥曼青霉素濃度的增加和作用時間的延長,增殖抑制作用明顯增強,呈現明顯的時效和量效關系。同時在渥曼青霉素濃度低于50 nmol·L-1時,DF-1-Pr P細胞具有抗渥曼青霉素的能力,與DF-1-NC和DF-1細胞相比有統計學差異(P<0.01);而當渥曼青霉素濃度大于50 nmol·L-1時其幾乎完全抑制了各細胞的增殖。
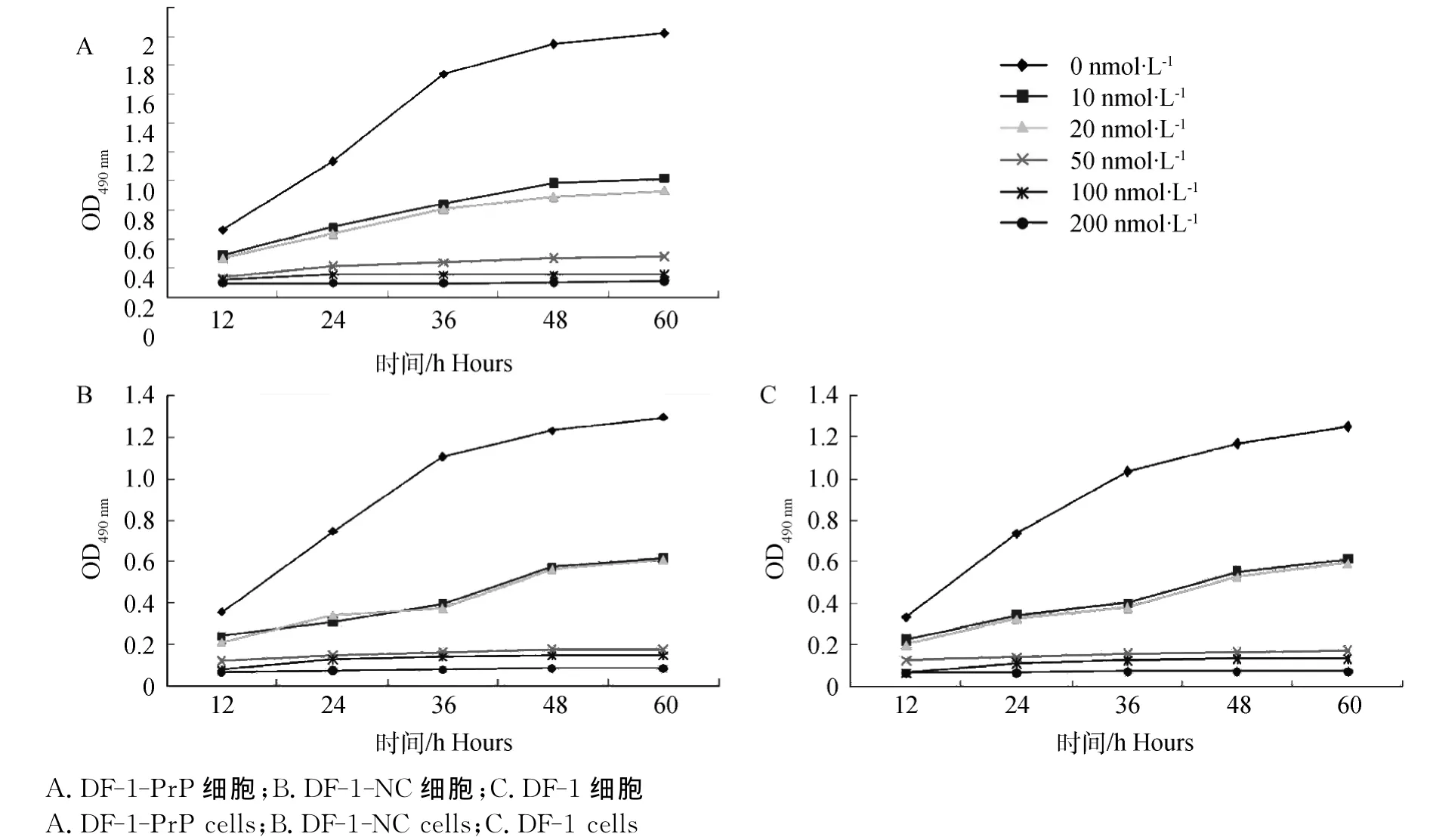
圖1 不同渥曼青霉素濃度對細胞增殖的影響Fig.1 Effects of different concentration wortmannin on cell proliferation
2.4 渥曼青霉素以劑量依賴方式促進DF-1-Pr P細胞凋亡
細胞經Annexin-V-FITC/PI雙染后流式細胞儀檢測,結果顯示(圖2),在無渥曼青霉素處理時,DF-1-Pr P細胞總凋亡率(5.96%)低于DF-1-NC細胞(10.27%)和DF-1細胞(13.85%)。隨著渥曼青霉素濃度的增加,各細胞的總凋亡率均升高,而且呈現一定的劑量效應關系;渥曼青霉素濃度低于20 nmol·L-1以下時,對各細胞的凋亡率影響較小,當渥曼青霉素濃度高于50 nmo·L-1時,渥曼青霉素濃度對各細胞的凋亡率影響非常大。在同一渥曼青霉素濃度下,DF-1-PrP細胞的凋亡率低于DF-1-NC和DF-1細胞;尤其渥曼青霉素濃度為100、200 nmol·L-1時,DF-1-NC和DF-1細胞幾乎全部凋亡,但DF-1-Pr P細胞仍有56.63%和15.54%的細胞存活,具有抗渥曼青霉素的能力。
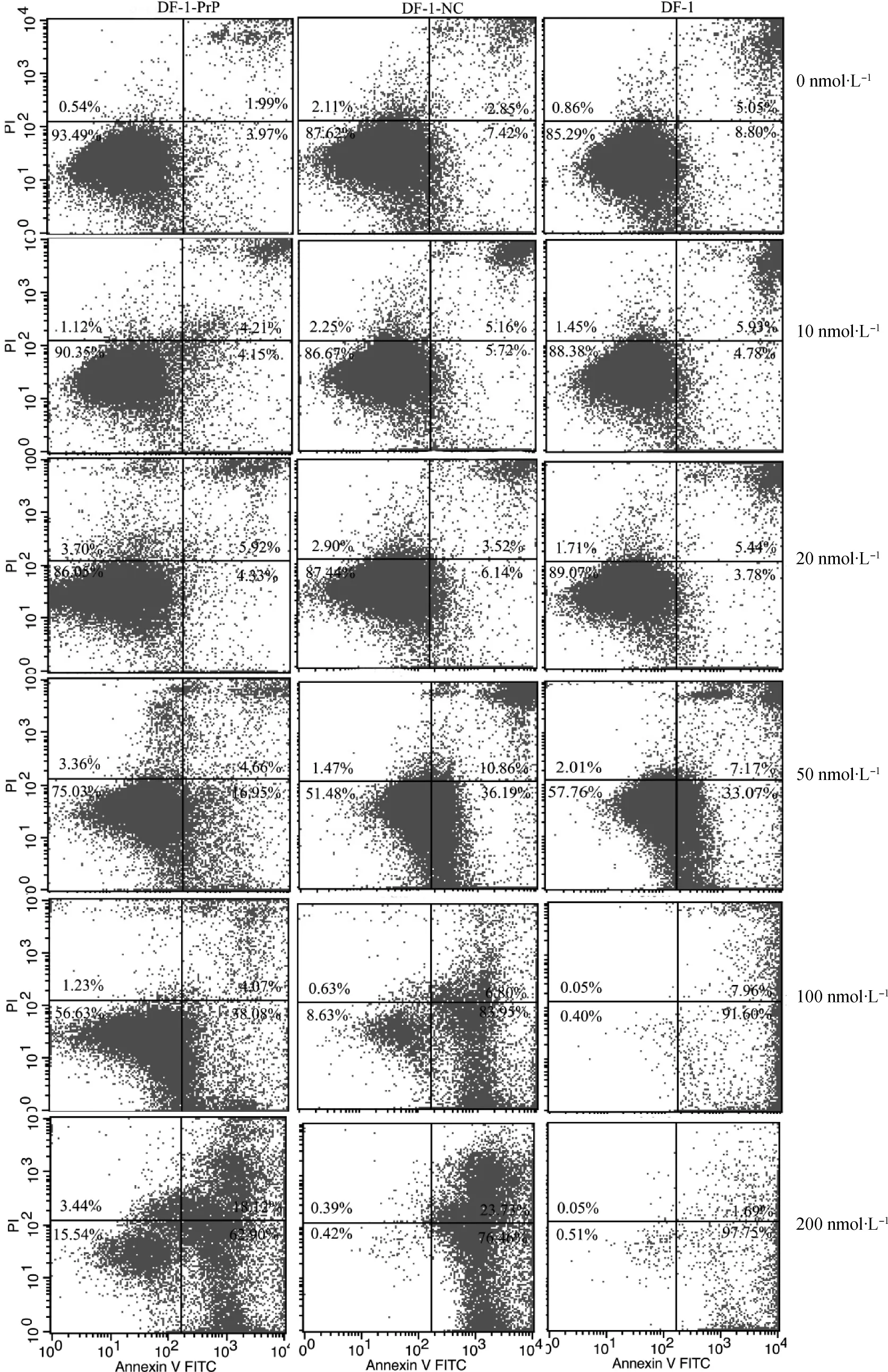
圖2 不同濃度渥曼青霉素對細胞凋亡的影響Fig.2 Effects of different concentration wortmannin on cell apoptosis
2.5 不同渥曼青霉素濃度下PRNP基因的轉錄量分析
利用本實驗室建立的雞PRNP基因m RNA實時熒光定量RT-PCR檢測方法,檢測DF-1-Pr P、DF-1-NC和DF-1細胞PRNP基因轉錄水平表達量,結果顯示(圖3),加入渥曼青霉素時各細胞PRNP基因的mRNA拷貝數均出現變化;隨著渥曼青霉素濃度的增加,PRNP基因的mRNA拷貝數呈現出下降的趨勢。在同一渥曼青霉素濃度下,DF-1-PrP細胞PRNP基因的mRNA拷貝數均顯著高于DF-1-NC和DF-1細胞。
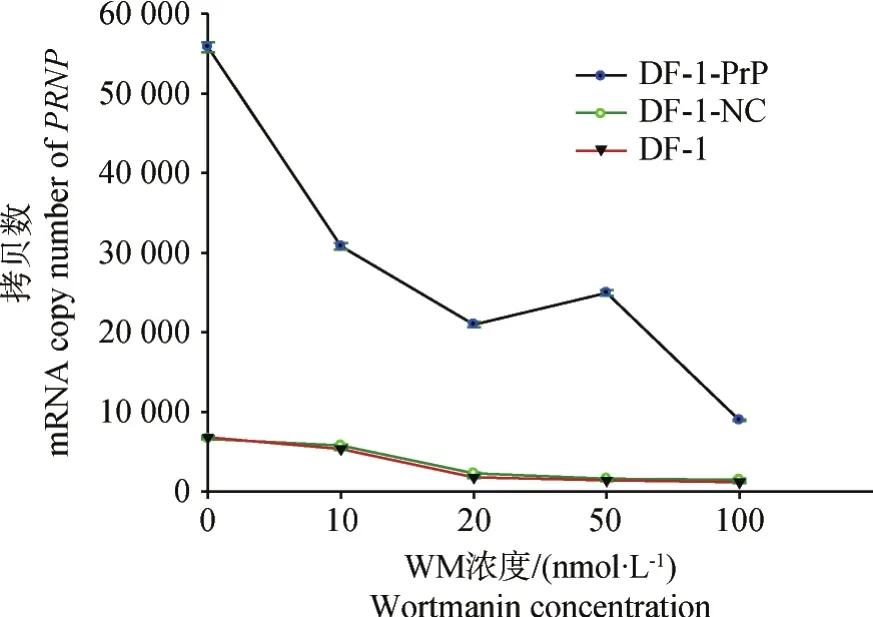
圖3 不同濃度渥曼青霉素對PRNP基因mRNA拷貝數的影響Fig.3 Effects of different wortmannin concentration on mRNA copy number of PRNP
3 討 論
Akt的活性與抗細胞凋亡以及細胞生長、增殖和能量代謝有關。在腫瘤細胞中Akt的表達量較正常細胞高,有加速白血病細胞增殖和抑制細胞程序性死亡的作用[14],激活Akt還可改變細胞遷移和侵襲,提高卵巢腫瘤細胞的侵襲能力[15]。敲除小鼠的PrPC基因可明顯降低p-Akt表達量,加劇腦缺血后的神經損傷[16]。Pr PC和p-Akt在胃癌組織中協同表達,p-Akt有可能在Pr PC介導的胃癌惡性表型中發揮作用[17]。但雞細胞型朊蛋白的生理功能及其機制還未見報道,為此作者構建了Ch Pr PC過表達的雞成纖維細胞系DF-1-Pr P及空載體細胞系DF-1-NC。本試驗中,作者應用PI3K/Akt抑制劑渥曼青霉素阻斷該信號通路,探討其對Pr PC介導的細胞增殖、黏附、侵襲和凋亡過程的影響。結果顯示,渥曼青霉素濃度為0 nmol·L-1時,DF-1-Pr P、DF-1-NC和DF-1細胞的增殖、黏附、侵襲能力均最高,總凋亡率均最低;且DF-1-Pr P細胞增殖、黏附、侵襲能力均高于DF-1-NC細胞和DF-1細胞,而總凋亡率卻低于DF-1-NC細胞和DF-1細胞,差異顯著(P<0.05),說明ChPrPC的過量表達可促進DF-1-Pr P細胞增殖、黏附、侵襲,抑制其凋亡,這和PrPC在哺乳動物中的功能相似[7]。隨著渥曼青霉素濃度的增加,DF-1-Pr P、DF-1-NC和DF-1細胞各自的PRNP基因mRNA表達量均減少,其增殖、黏附、侵襲能力相應下降,而總凋亡率均升高,且呈現一定的劑量效應關系,這證實了文獻報道的PI3K/Akt信號轉導通路是調節細胞存活最重要的途徑之一[18],ChPrPC至少部分是通過PI3K/Akt信號通路對DF-1細胞的增殖、黏附、侵襲和凋亡進行調節的。但在低于100 nmol·L-1的同一渥曼青霉素濃度下,DF-1-Pr P細胞增殖、黏附、侵襲能力始終高于DF-1-NC細胞和DF-1細胞,而總凋亡率均低于DF-1-NC細胞和DF-1細胞,差異顯著(P<0.05),說明ChPrPC的過量表達具有抗渥曼青霉素的能力。尤其在流式細胞術檢測細胞凋亡的試驗中,渥曼青霉素濃度為100和200 nmol·L-1時,DF-1-NC和DF-1細胞幾乎全部凋亡,但DF-1-Pr P細胞仍有56.63%和15.54%的細胞存活,提示ChPrPC還可通過其他途徑調節DF-1細胞的凋亡。
T.W.Poh等研究渥曼青霉素對PI3K/Akt信號通路的作用,結果表明渥曼青霉素能負調控PI3K/Akt信號通路中的多種分子,包括PI3K和m TOR,使Akt去磷酸化從而下調Akt激酶活性[18-19]。作者采用實時熒光定量RT-PCR法檢測發現,隨著渥曼青霉素濃度的增加,DF-1-Pr P、DF-1-NC和DF-1細胞中PRNP基因mRNA表達量均減少,而在同一渥曼青霉素濃度下,DF-1-Pr P細胞中PRNP基因mRNA表達量均高于DF-1-NC細胞和DF-1細胞,表明Akt基因表達量與PRNP基因表達量相關聯,Akt對PrPC可能存在反饋調節,但其機制尚不明確。
總之,ChPrPC的過量表達可促進DF-1細胞增殖、黏附和侵襲,抑制其凋亡;渥曼青霉素以劑量依賴方式抑制DF-1細胞中PRNP基因mRNA表達及其細胞的增殖、黏附和侵襲,誘導其凋亡,表明PI3K/Akt信號通路可能在Ch Pr PC介導DF-1細胞增殖、黏附、侵襲和凋亡過程中具有重要的作用。Ch Pr PC的過量表達可抗渥曼青霉素引起的細胞凋亡,提示PI3K/Akt信號通路并不是ChPrPC調節細胞凋亡的唯一途徑。本研究結果為進一步闡明ChPrPC生理功能的分子機制奠定基礎。
(
):
[1] OSAKI M,OSHIMURA M,ITO H.PI3K-Akt pathway:its functions and alterations in human cancer[J].Apoptosis,2004,9(6):667-676.
[2] PRUSINER S B.Novel proteinaceous infectious particles cause scrapie[J].Science,1982,216(4542):136-144.
[3] JI H F,ZHANG H Y.A comparative molecular dynamics study on thermostability of human and chicken prion proteins[J].Biochem Biophys Res Commun,2007,359(3):790-794.
[4] BROWN D R,CLIVE C,HASWELL S J.Antioxidant activity related to copper binding of native prion protein[J].J Neurochem,2001,76(1):69-76.
[5] RACHIDI W,MANGéA,SENATOR A,et al.Prion infection impairs copper binding of cultured cells[J].J Biol Chem,2003,278(17):14595-14598.
[6] PAN Y,ZHAO L,LIANG J,et al.Cellular prion protein promotes invasion and metastasis of gastric cancer[J].FASEB J,2006,20(11):1886-1888.
[7] MEHRPOUR M,CODOGNO P.Prion protein:From physiology to cancer biology[J].Cancer Lett,2010,290(1):1-23.
[8] DIARRA-MEHRPOUR M,ARRABAL S,JALIL A,et al.Prion protein prevents human breast carcinoma cell line from tumor necrosis factor alpha-induced cell death[J].Cancer Res,2004,64(2):719-727.
[9] ROUCOU X,GIANNOPOULOS P N,ZHANG Y,et al.Cellular prion protein inhibits proapoptotic Bax conformational change in human neurons and in breast carcinoma MCF-7 cells[J].Cell Death Differ,2005,12(7):783-795.
[10] KIM B H,LEE H G,CHOI J K,et al.The cellular prion protein(Pr PC)prevents apoptotic neuronal cell death and mitochondrial dysfunction induced by serum deprivation[J].Brain Res Mol Brain Res,2004,124(1):40-50.
[11] DIAO X L,WU R,LIU L,et al.Expression patterns of PrP gene during chicken embryo development[J].Asian J Anim Vet Adv,2011,7:199-204.
[12] 刁小龍.雞朊蛋白參與DF-1細胞增殖粘附的功能研究[D].蘭州:甘肅農業大學,2012:60.DIAO X L.The function research of ChPrPCinvolved in the processes of DF-1 cells proliferation and adhesion[D].Lanzhou:Gansu Agricultural University,2012:60.(in Chinese)
[13] 朱曼玲,吳 潤,于宏偉,等.雞Akt基因m RNA SYBR GreenⅠ實時熒光定量RT-PCR檢測方法的建立[J].中國獸醫科學,2014,44(7):758-764.ZHU M L,WU R,YU H W,et al.Establishment of SYBR GreenⅠreal-time RT-PCR for detection of chicken Akt gene m RNA[J].Chinese Veterinary Science.2014,44(7):758-764.(in Chinese)
[14] 王曉南,吳 青,張連生,等.PI3K/Akt抑制劑渥曼青霉素對白血病細胞增殖和凋亡的影響[J].中國腫瘤生物治療雜志,2010,17(1):82-87.WANG X N,WU Q,ZHANG L S,et al.Effect of PI3K/Akt inhibitor wortmannin on proliferation and apoptosis of leukemia K562 cells[J].Chinese Journal of Cancer Biotherapy,2010,17(1):82-87.(in Chinese)
[15] LEE S,CHOI E J,JIN C,et al.Activation of PI3K/Akt pathway by PTEN reduction and PIK3CA mRNA amplification contributes to cisplatin resistance in an ovarian cancer cell line[J].Gynecol Oncol,2005,97(1):26-34.
[16] WEISE J,SANDAU R,SCHWARTING S,et al.Deletion of cellular prion protein results in reduced Akt activation,enhanced postischemic caspase-3 activation,and exacerbation of ischemic brain injury[J].Stroke,2006,37(5):1296-1300.
[17] 葛伏林,呂 焱,吳本儼,等.朊蛋白Pr PC及磷酸化Akt在胃癌中共表達及其臨床意義[J].軍醫進修學院學報,2011,32(9):962-964.GE F L,LV Y,WU B Y,et al.Co-expression of PrPCand p-Akt in gastric cancer and its clinical significance[J].Journal of Chinese PLA Postgarduate Medical School,2011,32(9):962-964.(in Chinese)
[18] OHTA T,OHMICHI M,HAYASAKA T,et al.Inhibition of phosphatidylinositol 3-kinase increases efficacy of cisplatin in in vivo ovarian cancer models[J].Endocrinology,2006,147(4):1761-1769.
[19] POH T W,PERVAIZ S.LY294002 and LY303511 sensitize tumor cells to drug-induced apoptosis via intracellular hydrogen peroxide production independent of the phosphoinositide 3-kinase-Akt pathway[J].Cancer Res,2005,65(14):6264-6274.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
