乳酸乳球菌生產γ-氨基丁酸條件的優化
2015-03-17 05:36:22楊詩穎柯崇榕黃建忠
食品科學 2015年7期
陳 佳,楊詩穎,柯崇榕,黃建忠*
(福建師范大學生命科學學院,工業微生物發酵技術國家地方聯合工程研究中心,福建 福州 350117)
?
乳酸乳球菌生產γ-氨基丁酸條件的優化
陳 佳,楊詩穎,柯崇榕,黃建忠*
(福建師范大學生命科學學院,工業微生物發酵技術國家地方聯合工程研究中心,福建 福州 350117)
摘 要:采用析因設計和中心組合試驗設計對乳酸乳球菌FJNU-GA1304產γ-氨基丁酸(γ-aminobutyric acid,GABA)的條件進行優化。完全析因設計優化后的細胞轉化條件為:pH 3.5、反應溫度40 ℃、反應時間24 h,谷氨酸鈉質量濃度20 g/L和濕菌體質量濃度25 g/L;在單因素試驗的基礎上,通過篩選設計確定谷氨酸鈉、玉米漿粉和葡萄糖質量濃度為主效因子。采用三因素三水平的中心組合試驗對主效因子的交互作用進行分析,結果表明:最佳的培養基組成為谷氨酸鈉9.50 g/L、玉米漿粉12.50 g/L、葡萄糖5.74 g/L、酵母膏5.00 g/L、K2HPO41.2 0 g/L、MgSO40.60 g/L。在最佳轉化條件和發酵培養基組合下,GABA產量最高達9.06 g/L,比優化前4.80 g/L提高了88.8%。
關鍵詞:γ-氨基丁酸;乳酸乳球菌;析因設計;篩選設計;中心組合試驗設計
γ-氨基丁酸(γ-aminobutyric acid,GABA)是由L-谷氨酸或其衍生物在谷氨酸脫羧酶的作用下脫羧而成的一種非蛋白質組成的天然氨基酸,廣泛存在于微生物、植物和動物細胞中[1-3]。GABA是哺乳動物中樞神經系統的抑制性傳遞物質,具有降血氨、抗焦慮、降血壓、治療糖尿病、改善腎機能和肝機能,促進乙醇代謝和消臭,以及高效減肥等生理功能,廣泛應用于食品和醫藥行業中[4-9]。此外,GABA也作為飼料添加劑應用于畜牧業中[10-12]。
目前,GABA生產主要是通過微生物進行發酵[13]。乳酸菌是國際公認的食品安全級細菌,2009年12月國家衛生部已批準GABA用于食品生產加工,因此大部分研究集中于利用乳酸菌發酵生產GABA[14]。然而,乳酸菌屬于兼性厭氧菌,生物量較低,直接以谷氨酸鈉為底物進行發酵生產,底物殘留較多,增加了下游提取純化難度。林謙等[15]以10 g/L的谷氨酸鈉為底物利用Lactobacillus fermentum YS2液體發酵生產GABA,產量僅為4.37 g/L;Ratanaburee等[16]以10 g/L的谷氨酸鈉為底物利用Lactobacillus plantarum DW12液體發酵生產GABA,產量也只有4.00 g/L。同時,乳酸菌是以有機培養基進行發酵,發酵液中還有微生物菌體、代謝物、蛋白和色素等多種雜質,產品分離純化步驟繁雜、設備要求高、成本高。針對此問題,張婷[17]探討了兩步法合成GABA。第一步是發酵收集菌體,第二步是全細胞催化GABA。相對于發酵液而言,反應液成分簡單,不含大量有機成分,使產物分離純化較簡單。同時具有生產周期較短、轉化率高及節約成本等優點。因此,本實驗采用兩步法生產GABA,探討細胞轉化條件對GABA產量和培養基組分對谷氨酸脫羧酶活性的影響,從而為規模化生產GABA提供一定的理論基礎。
1 材料與方法
1.1材料與試劑
乳酸乳球菌(Lactococcus lactis)FJNU-GA1304,由工業微生物發酵技術國家地方聯合工程研究中心(福建師范大學)保藏。
γ-氨基丁酸標準品(純度≥99%)美國Sigma公司;異硫氰酸苯酯美國Aladdin公司;乙腈(色譜純) 美國Sinence公司;其余試劑均為國產分析純。
1.2儀器與設備
U-3000高效液相色譜系統(配有DAD檢測器)美國Thermo Scientific公司。
1.3培養基
種子培養基(g/L):牛肉膏10、酵母膏5、葡萄糖20、蛋白胨10、K2HPO42、無水乙酸鈉 5、檸檬酸三銨2、MgSO4·7H2O 0.58、MnSO4·4H2O 0.25,吐溫-80 1 mL,pH 6.8。
初始發酵培養基(g/L):酵母膏10、葡萄糖10、K2HPO41、MgSO4·7H2O 1、谷氨酸鈉5,pH 7.0。
1.4液體培養條件
按體積分數5%的初始接種量將種子液接入發酵培養基中,裝液量為35 mL/150 mL,溫度為30 ℃,靜置培養12 h。
1.5GABA產量的測定
發酵液離心,收集菌體,稱取濕菌體質量,計算對應的生物量。采用含有10 g/L谷氨酸鈉的醋酸-醋酸鈉緩沖溶液重懸菌體,使最終轉化液中濕菌體質量濃度為4 g/100 mL,在一定條件下反應后,以沸水浴10 min結束反應,10 000×g離心5 min,取上清液測定GABA的質量濃度,是以異硫氰酸苯酯(phenylisothiocyanate,PITC)作為衍生劑通過高效液相色譜進行測定[18]。
色譜條件:Hypersil GOLD C18色譜柱(250 mm× 4.6 mm,5 μm),檢測波長254 nm,流動相:V(醋酸鈉緩沖液)∶V(乙腈)=4∶1,柱溫40 ℃,流速0.8 mL/min,進樣量5 μL。
1.6細胞轉化條件的優化
使用N=20完全析因試驗對影響細胞轉化的pH值、反應溫度、反應時間、谷氨酸鈉和濕菌體的質量濃度5 個因素進行優化,見表1。

表1 細胞轉化條件因素Table 1 Level and code of conversion conditions used for full factorial design
1.7培養基組分的優化
1.7.1 單因素試驗和篩選設計
以初始發酵培養基為基礎,分別以10 g/L的可溶性淀粉、蔗糖、丁二酸鈉、葡萄糖、乳糖和果糖作為單一碳源,確定最佳碳源;再分別以10 g/L的硝酸鈉、硫酸銨、玉米漿粉、酵母膏、蛋白胨、牛肉膏和胰蛋白胨作為單一氮源,確定最佳氮源。根據最適碳氮源修改初始發酵培養基組分,采用JMP中的N=16的篩選設計分析培養基組分中影響GABA產量的主效因素。
1.7.2 中心組合試驗設計
通過最陡爬坡試驗確定3 個主效因素的中心點,利用SAS 9.2軟件根據響應面曲面設計原理,采用三因素三水平的中心組合試驗設計,確定主效因素的最佳水平。
2 結果與分析
2.1細胞轉化條件優化

圖1 細胞轉化條件優化實驗結果Fig.1 Results of optimal conversion conditions
細胞轉化生產GABA實際上是利用細胞內的谷氨酸脫羧酶(glutamate decarboxylase,GAD)將運輸通過細胞壁進入胞內的谷氨酸進行脫羧形成。如圖1所示,通過標準最小二乘法構建模型的線性擬合表明,pH值是最主要的影響因素,當pH值為5.5時,GABA產量均低于0.124 g/L;其次是谷氨酸鈉,在pH 3.5的基礎上,當谷氨酸鈉的質量濃度為20 g/L時,GABA產量均大于3 g/L;且當pH 3.5時,底物質量濃度20 g/L時,反應溫度為40 ℃更有利于提高GABA的產量。這是因為任何酶都需要在合適的pH值和溫度條件下才能體現出其最大的活性,而且在酸性條件下細菌需要通過將胞外的谷氨酸運入胞內脫羧生成氨基丁酸并消耗一質子來維持自身pH值[19]。因此,較低的pH值和高質量濃度的谷氨酸鈉有利于GABA的生成。此外,較高的溫度能夠提高細胞壁的通透性,有利于谷氨酸進入細胞內。因而最終確定pH 3.5、反應溫度40 ℃、反應時間24 h、谷氨酸鈉質量濃度20 g/L和濕菌體質量濃度25 g/L作為細胞轉化GABA的條件。
2.2碳氮源的選擇
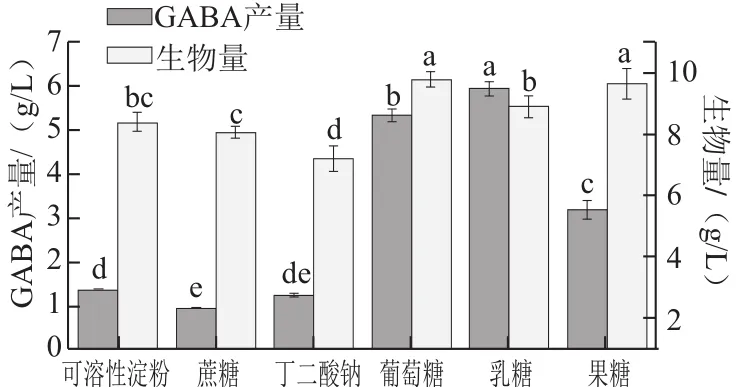
圖2 不同碳源對GABA產量的影響Fig.2 Effect of carbon sources on GABA production
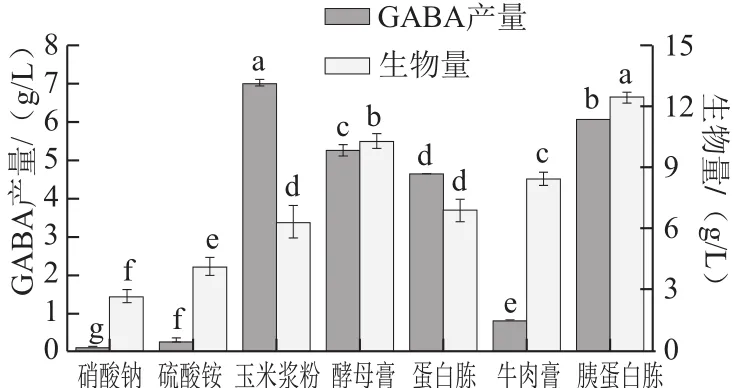
圖3 不同氮源對GABA產量的影響Fig.3 Effect of nitrogen sources on GABA production
細胞內單位細胞的轉化效率受培養基組分的影響,因此需要對培養基組分進行優化。首先探討最適的碳氮源,如圖2所示,以乳糖為碳源最有利于提高單位細胞轉化效率,GABA產量最高達到5.93 g/L;其次為葡萄糖5.33 g/L。通過方差分析和多重比較表明,不同碳源對GABA的產量以及生物量的影響差異顯著。考慮到葡萄糖有利于菌體生長且價格低廉,故選定葡萄糖作為碳源。
以葡萄糖為碳源,分析不同氮源對GABA產量的影響。如圖3所示,以玉米漿粉、胰蛋白胨或酵母膏作為氮源時,GABA產量較高,分別為6.98、6.02 g/L和5.23 g/L。黃桂東等[20]發現采用復合氮源有利于菌體內單位細胞轉化效率的提高,從而提高GABA的產量。由于胰蛋白胨價格高昂,因此選取玉米漿粉和酵母膏作為復合氮源。考慮到玉米漿粉最有利于提高GABA產量,故確定玉米漿粉和酵母膏的質量濃度分別為7 g/L和3 g/L。
2.3篩選設計
培養基組分較多,需先對各個組分進行擬合分析,篩選出主效組分。在單因素試驗的基礎上,采用N=16的篩選設計,結果如表2所示。
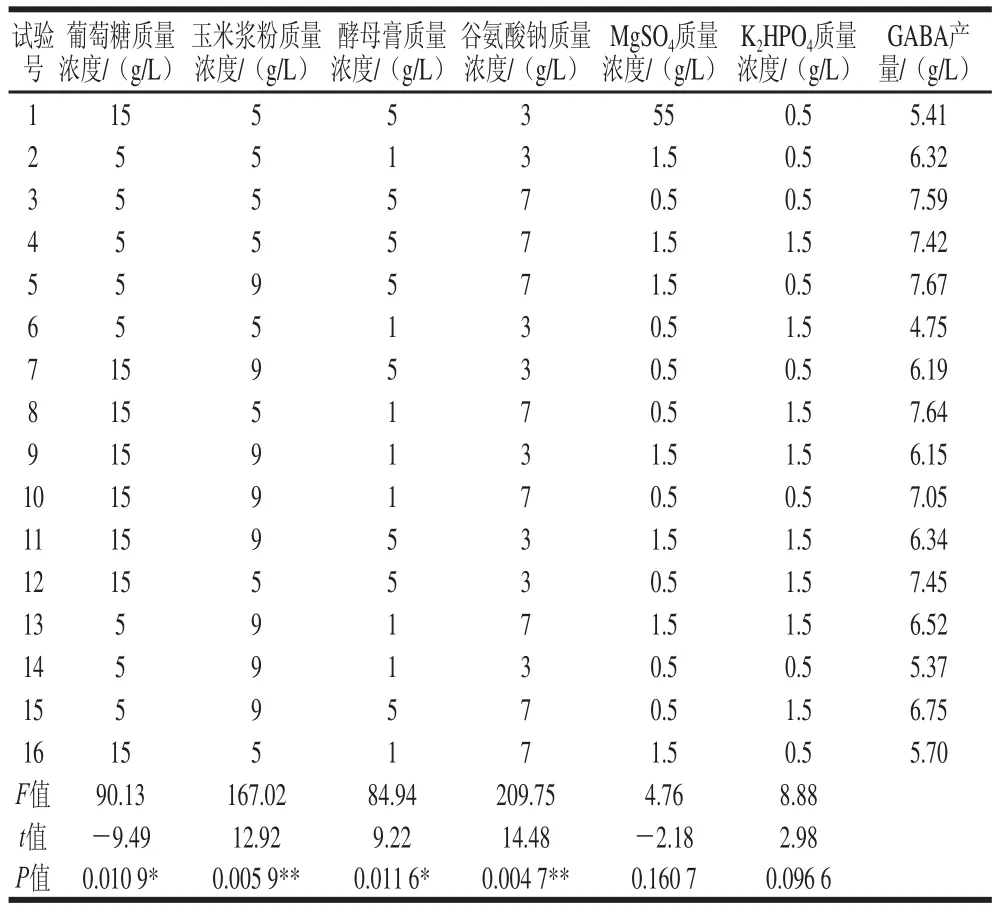
表2 主效組分的篩選設計與結果Table 2 Screening design of medium components and results
利用JMP 11.0進行數據處理和因素效應分析,結果表明,谷氨酸鈉和玉米漿粉質量濃度對GABA產量影響最大(P<0.01),其次為葡萄糖和酵母膏質量濃度(P<0.05)。這是由于GABA是由谷氨酸脫羧酶(GAD)脫羧形成的,而GAD是誘導酶,需要一定質量濃度的谷氨酸鈉作為誘導物。此外,回歸分析表明葡萄糖和MgSO4為負效應因素,適當減少其質量濃度有利于提高GABA產量。
2.4最陡爬坡試驗設計
根據篩選設計結果,確定谷氨酸鈉、玉米漿粉和葡萄糖為主效因子進行最陡爬坡試驗,逼近GABA的最大產量區,而酵母膏、K2HPO4和MgSO4的質量濃度分別設為5、1.2 g/L和0.6 g/L。由表3可知,第5組試驗的GABA產量最高,達到8.61 g/L。因此,選擇葡萄糖、玉米漿粉和谷氨酸鈉質量濃度分別為6、11.2 g/L和11 g/L時作為中心組合試驗設計的中心點。

表3 最陡爬坡試驗設計及結果Table 3 Steepest aasscent design with experimental values of GABA yield
2.5中心組合試驗設計
2.5.1 中心組合試驗結果與分析
葡萄糖、玉米漿粉和谷氨酸鈉的質量濃度響應面試驗方案及結果見表4。16 組試驗點中有14 個析因點,2 個用以估算試驗誤差的零點。利用SAS 9.2軟件對數據進行回歸分析,發現GABA響應值均在95%的置信區間內,且殘差在±5 RSD之間,正態分布呈一直線,表明該試驗結果可信、沒有噪音數據。
2.5.2 中心組合試驗顯著性檢驗
方差分析(表5)表明,不同組分間的差異顯著,表明試驗方法準確可靠,通過擬合回歸構建二階響應模型是可行的,回歸模型P值小于0.01且相關系數R2=96.40%(>90%),表明該模型極顯著且擬合良好,可用于分析預測不同組分的質量濃度配比對GABA產量的影響。

表5 二次回歸模型的方差分析結果Table 5 Analysis of variance for the fitted quadratic regression model
由表5中的P值可知,葡萄糖、玉米漿粉及其與谷氨酸鈉的交互作用對GABA產量有極顯著影響(P<0.01);谷氨酸鈉和葡萄糖二次項對GABA產量有顯著影響(P<0.05),表明不同質量濃度的葡萄糖、玉米漿粉和谷氨酸鈉對GABA產量的影響呈曲面關系,交互項作用顯著,二次項影響較小。根據模型預測分析,當葡萄糖為5.74 g/L、玉米漿粉為12.5 g/L和谷氨酸鈉為9.50 g/L時,GABA最高產量可達9.47 g/L。
2.6驗證實驗
根據中心組合試驗確定的最優培養基配方進行搖瓶發酵驗證,以優化前的條件做對照,實驗重復3 次。3 次實驗優化后的GABA平均產量為9.06 g/L,與預測產量9.47 g/L相差不到5%,說明模型較好地擬合了不同組分配比對GABA產量的影響。
3 結 論
本研究對一株具有產GABA能力的乳酸乳球菌(Lactococcus lactis)FJNU-GA1304進行了細胞轉化條件和培養基組分的優化。通過完全析因設計對細胞轉化條件優化,確定了有利于提高GABA產量的細胞轉化條件是pH 3.5、反應溫度40 ℃、反應時間24 h、谷氨酸鈉質量濃度20 g/L和濕菌體質量濃度25 g/L。GABA的產量直接受谷氨酸脫羧酶影響[21]。要提高GABA的產量,首先應獲得較高產量的GAD,提高單位細胞轉化率,即培養基組分必須有利于細胞的增殖,同時菌體還應具有較高的GAD活性。因此,在優化后的細胞轉化條件的基礎上,通過單因素試驗結合中心組合試驗,對發酵培養基組分進行優化,以期提高單位質量菌體內的GAD產量,最終提高GABA的產量。優化后得到有利于提高GABA生成量的發酵培養基組分為谷氨酸鈉9.50 g/L、玉米漿粉12.50 g/L、葡萄糖5.74 g/L、酵母膏5.00 g/L、K2HPO41.20 g/L、MgSO40.60 g/L。在最佳轉化條件和發酵培養基組合下,轉化液中GABA產量為9.06 g/L,比優化前4.80 g/L提高了88.8%。
參考文獻:
[1]LIN Qian. Submerged fermentation of Lactobacillus rhamnosus YS9 for gamma-aminobutyric acid (GABA) production[J]. Brazilian Journal of Microbiology, 2013, 44(1): 183-187.
[2]MATSUYAMA A, YOSHIMURA K, SHIMIZU C, et al. Characterization of glutamate decarboxylase mediating γ-amino butyric acid increase in the early germination stage of soybean[J]. Journal of Bioscience and Bioengineering, 2009, 107(5): 538-543.
[3]RICHARDSON G, DING H, ROCHELEAU T, et al. An examination of aspartate decarboxylase and glutamate decarboxylase activity in mosquitoes[J]. Molecular Biology Reports, 2010, 37(7): 3199-3205.
[4]WAAGEPETERSEN H S, SONNEWALD U, SCHOUSBOE A. The GABA paradox: multiple roles as metabolite, neurotransmitter, andneurodifferentiative agent[J]. Journal of Neurochemistry, 1999, 73(4): 1335-1342.
[5]江波. GABA(γ-氨基丁酸): 一種新型的功能食品因子[J]. 中國食品學報, 2008, 8(2): 1-4.
[6]DEFEUDIS F. γ-Aminobutyric acid and cardiovascular function[J]. Experientia, 1983, 39(8): 845-849.
[7]HAYAKAWA K, KIMURA M, K ASAHA K, et al. Effect of γ-aminobutyric acid-enriched dairy product on the blood pressure of spontaneously hypertensive and normotensive Wistar-Kyoto rats[J]. British Journal of Nutrition, 2004, 92(3): 411-417.
[8]COHEN I, NAVARRO V, CLEMENCEAU S, et al. On the origin of interictal activity in human temporal lobe epilepsy in vitro[J]. Science, 2002, 298: 1418-1421.
[9]KRNJEVI? K. Chemical nature of synaptic transmission in vertebrates[J]. Physiological Reviews, 1974, 54(2): 418-540.
[10]ZHANG Min, ZOU Xiaoting, LI Hui, et al. Effect of dietary γ-aminobutyric acid on laying pe rformance, egg quality, immune activity and endocrine hormone in heat-stressed Roman hens[J]. Animal Science Journal, 2012, 83(2): 141-147.
[11]鄭藝梅, 劉麗妹, 符穩群. 家禽生產中γ-氨基丁酸的應用[J]. 糧食與飼料工業, 2014(1): 46-49.
[12]楊勝遠, 陸兆新, 呂風霞, 等. γ-氨基丁酸的生理功能和研 究開發進展[J]. 食品科學, 2005, 26(9): 546-551.
[13]LI Haixing, CAO Yusheng. Lactic acid bacterial cell factories for gamma-aminobutyric acid[J]. Amino Acids, 2010, 39(5): 1107-1116.
[14]食品與發酵工業編輯部. 衛生部批準γ-氨基丁酸等6種新資源食品用于食品生產加工[J]. 食品與發酵工業, 2010, 36(2): 144.
[15]林謙, 姜康怡, 韋素娟, 等. 產GABA發酵乳 桿菌的篩選、發酵條件優化及其谷氨酸脫羧酶基因的克隆[J]. 廣東農業科學, 2014(8): 192-197.
[16]RATANABUREE A, KANTACHOTE D, CHARERNJIRATRAKUL W, et al. Enhancement of γ-aminobutyric acid in a fermented red seaweed beverage by starter culture Lactobacillus plantarum DW12[J]. Electronic Journal of Biotechnology, 2011, 14(3): 1-14.
[17]張婷. HPLC法測定轉化液中γ-氨基丁酸含量及轉化液 的預處理研究[D]. 鄭州: 鄭州大學, 2012.
[18]許建軍, 江波, 許時嬰. 比色法快速測定乳酸菌谷氨酸脫羧酶活力及其應用[J]. 微生物學通報, 2004(2): 66-71.
[19]KAN JEE U, HOURY W A. Mechanisms of acid resistance in Escherichia coli[J]. Annual Review of Microbiology, 2013, 67: 65-81.
[20]黃桂東, 毛健, 姬中 偉, 等. 一株產γ-氨基丁酸植物乳桿菌mj0301培養基的優化[J]. 食品科學, 2013, 34(17): 165-170. doi: 10.7506/spkx1002-6630-201317036.
[21]FAN Euyu, HUANG Jun, HU Sheng, et al. Cloning, sequencing and expression of a glutamate decarboxylase gene from the GABA-producing strain CG MCC 1306[J]. Annals of Microbiology, 2012, 62(2): 689-698.
Optimization of Conditions for the Production of γ-Aminobutyric Acid by Lactococcus lactis
CHEN Jia, YANG Shiying, KE Chongrong, HUANG Jianzhong*
(National-Local United Engineering Research Centre of Industrial Microorganism Fermentation Technology, College of Life Science Fujian Normal University, Fuzhou 350117, China)
,
Abstract:Factorial design and central composite design were applied to optimize the yield of γ-aminobutyric acid (GABA) produced by cultured Lactococcus lactis cells from monosodium glutamate in a buffer solution. A full factorial design was used to optimize the conversion conditions as 40 ℃, pH 3.5 and 24 h. Using single factor designs, sodium glutamate, corn steep powder and glucose concentration were identified as major factors affecting GABA production. The interaction effect of the major factors at three levels each was analyzed by central composite design. The results showed that the optimal medium for Lactococcus lactis consisted of 9.5 g/L sodium glutamate, 5.74 g/L glucose, 12.50 g/L corn steep powder, 5.00 g/L yeast extract, 1.20 g/L K2HPO4and 0.60 g/L MgSO4. Under these conditions, the yield of GABA was 9.06 g/L, which was increased by 88.8% when compared with that (4.80 g/L) before optimization.
Key words:γ-aminobutyric acid; Lactococcus lactis; factorial design; screening design; central composite design
doi:10.7506/spkx1002-6630-201507027
中圖分類號:TQ922
文獻標志碼:A
文章編號:1002-6630(2015)07-0148-05
*通信作者:黃建忠(1966—),男,教授,博士,研究方向為微生物功能基因。E-mail:hjz@fjnu.edu.cn
作者簡介:陳佳(1990—),女,碩士研究生,研究方向為食品微生物。E-mail:happierjia@163.com
基金項目:福建省自然科學基金重點項目(2012N0013)
收稿日期:2014-05-04
