隱藻藻藍蛋白與類囊體膜的動態結合模型
2015-04-11 03:26:24徐偉,梁源,陳敏,王寧
海洋科學 2015年4期
徐 偉, 梁 源, 陳 敏, 王 寧
(煙臺大學 生命科學學院, 山東 煙臺264005)
在光合系統中捕光復合物(light-harvesting complex, LHC)的功能是負責吸收和傳遞光能, 并將激發能經特定的方式和途徑傳遞給反應中心用于光合作用。綠色植物和雜色植物的LHC分別為脂溶性的葉綠素(Chl)a/b-蛋白復合物和 Chl a/c-蛋白復合物, 而紅藍植物為水溶性的藻膽蛋白, 可以確定它們均以不同的方式結合于葉綠體的類囊體膜上[1]。但雜色植物隱藻卻非常特殊, 除了膜上的 Chl a/c2-蛋白復合物外, 還含有水溶性的藻膽蛋白。與紅、藍藻中的藻膽蛋白相比, 隱藻藻膽蛋白有許多特異之處: 只含有隱藻藻紅蛋白(Cr-PE)或隱藻藻藍蛋白(Cr-PC), 不含異藻藍蛋白(APC); 通常以二聚體(α1α2ββ)形式存在, 而非三聚體或六聚體; 不形成藻膽體樣結構等等。而最突出的差別是隱藻藻膽蛋白存在于類囊體腔內, 而不像藻膽體那樣附著于類囊體膜外表面[2-3]。但目前對于隱藻藻膽蛋白在膜腔內的存在狀態, 以及與膜的關系卻爭議很大。主要有以下幾種看法: (1)緊密結合在類囊體膜的內表面上[4]; (2)一部分與膜結合, 其余游離或者散布在類囊體腔中[5]; (3)以某種方式結合并橫跨膜腔內部, 邊緣與光合系統 II(PSII)接觸并為其提供能量[6]; (4)均勻地填充于類囊體腔中,彼此之間以及與膜上 Chl-蛋白復合物之間沒有傾向性的結合關系[7-8]。
目前, 與研究較為透徹的紅、藍藻藻膽蛋白及藻膽體相比, 對隱藻藻膽蛋白的了解要少得多, 尤其是隱藻藻膽蛋白與膜上另一套捕光復合物 Chl a/c2-蛋白復合物以及PSI和PSII反應中心之間的相互接觸方式不明, 使能量傳遞機制更無法最終確定。作者以含有隱藻藻藍蛋白 PC645的藍隱藻(Chroomonas placoidea)為材料, 根據室溫和低溫光譜測定和分析結果, 提出對這一問題的看法。
1 材料與方法
1.1 藍隱藻的培養及藻體細胞的收集
藍隱藻在實驗室采用F/2培養基自行培養, 溫度20~25℃, 持續通氣、光照培養, 16W日光燈, 光照強度約2 000 lx。取培養至4~5 d達到對數生長期的培養液, 于3 500g離心10 min收集藻體細胞, 用分離介質(pH7.2的50 mmol/L磷酸緩沖液, 含有0.3 mol/L的蔗糖, 0.1 mol/L NaCl和MgCl2)洗滌兩次。
1.2 藍隱藻類囊體膜的制備
在清洗后的藻體細胞中加入懸浮液(50 mmol/L,pH8.0的 Tricine-NaOH)后, 用 French Pressure Cell Press(美國 SLM Aminco)破碎細胞, 相對壓強為6.897 MPa。破碎后的溶液在1 000g離心10 min去除殘余細胞及大顆粒, 于5 000g離心10 min收集類囊體膜。以懸浮液懸浮, 小心攪拌, 清洗2~3次, 以除去膜中殘留的游離 PC, 直至離心上清液中不再有藍色出現為止。加入含 50% 甘油的懸浮液, 參照Jeffrey公式[9]調整葉綠素濃度為0.8~1.5 g/L, 于–80℃冰箱凍存備用。以上所有操作在4℃避光進行。
1.3 藻藍蛋白PC645的制備
細胞破碎后的溶液離心收集去除類囊體膜后的上清液于12 000g再次離心10 min, 上清液為含有PC645的粗提液。采用50%飽和度硫酸銨沉淀去除葉綠素等組分, 80%飽和度下得到PC645沉淀, 12 000g離心15 min收集沉淀, 加入少量PBS緩沖液溶解備用。經Sephadex G-100分子篩層析純化, 洗脫液為50 μL/L的PBS緩沖液得到初步純化的PC645。
1.4 室溫吸收光譜測定
藍隱藻藻體、類囊體膜用懸浮液適當稀釋后, 使用TU-1900雙波長雙光束紫外可見分光光度計(北京普析通用)測定室溫吸收光譜。
1.5 室溫熒光光譜測定
加入懸浮液將藍隱藻藻體、類囊體膜適當稀釋,使用 LS55 熒光分光光度計(Perkin Elmer)測定室溫熒光光譜。掃描速度500 nm/min, 激發和發射狹縫寬度10.0 nm。
1.6 77K熒光光譜測定
將藍隱藻藻體或類囊體膜離心去除懸浮液后,加入含 60%甘油的懸浮液懸浮, 或者于懸浮液中加入甘油至 60%(V/V), 混勻后置于低溫石英管中, 用LS55 熒光分光光度計在液氮溫度下測定熒光光譜。掃描速度 500 nm/min, 測定發射光譜時激發狹縫15.0 nm, 發射狹縫10.0 nm; 測定激發光譜時發射狹縫20.0 nm, 激發狹縫10.0 nm。
1.7 SDS-PAGE
將低溫保存的類囊體膜解凍后, 用懸浮液清洗,去除游離的PC后, 用懸浮液重新懸浮, 調整葉綠素質量濃度0.8 g/L。參照Laemmli[10]方法加入等體積樣品裂解液, 于37℃增溶30 min, 36 000g離心10 min去除膜碎片。上清液加入 80% 丙酮/乙醇(V/V)洗滌抽提脂類和葉綠素, 三氟乙酸沉淀蛋白, 40 000g離心15 min收集沉淀。沉淀及PC樣品參照Laemmli[10]方法進行 SDS-PAGE, 只是采用 10%~20%線性梯度膠分離。電泳結束后先用硫酸鋅染色觀察有熒光的條帶并拍照, 然后以 7% 冰乙酸脫色至背景透明后再用考馬斯亮藍染色, 0.5 mol/L NaCl脫色后拍照。
2 實驗結果
2.1 吸收光譜
在藍隱藻完整藻體細胞中(圖1), 436(438)nm和677(679) nm的吸收峰源自Chl a; 495 nm處吸收峰來自類胡蘿卜素; 而 586、647 nm 兩個吸收主峰和 1個626 nm的肩峰都屬于PC(581、626和645 nm), 為典型的Cr-PC645類型。463 nm左右的吸收峰與Chl c的藍區吸收有關, 但其在紅區吸收峰(630 nm附近)被PC645的吸收所掩蔽。在活體細胞中, PC吸收峰的高度超過Chl a, 表明PC在活體細胞中含量很高,是藻體內主要的捕光復合物。Chl c在紅區的吸收可由PC取代, 但在藍區的吸收比Chl a更靠近綠光區。這與隱藻多生長于以藍綠光為主的深水環境相適應,大量的PC以及一定量的Chl c的存在, 有利于藻體捕獲水體弱光環境中的有效光能, 以彌補Chl a吸收的不足[11]。充分洗滌過的類囊體膜中(圖2)Chl a紅區吸收紅移至679 nm, 在580~650 nm附近依然能看到屬于PC的吸收峰存在(對應于導數光譜中的690、634和650 nm倒置峰), 但與Chl a吸收相比, 相對強度明顯降低。這是由于葉綠體破碎后存在于類囊體腔中的PC大部分流失造成的。這種保留有藻膽蛋白的類囊體膜樣品, 在以往的報道中都未有提及。只是這部分PC的存在狀態, 單純通過吸收光譜不能確定,可能與膜結合, 但也可能處于游離狀態與類囊體膜片共存。
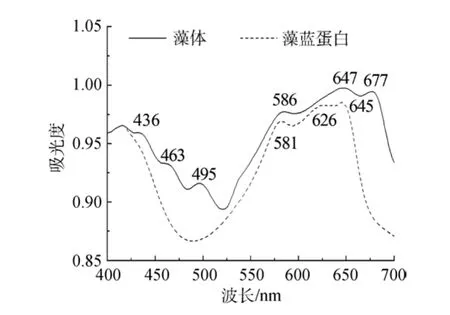
圖1 藻體和藻藍蛋白室溫吸收光譜Fig.1 Absorption spectra of cells and PC645 at room temperature
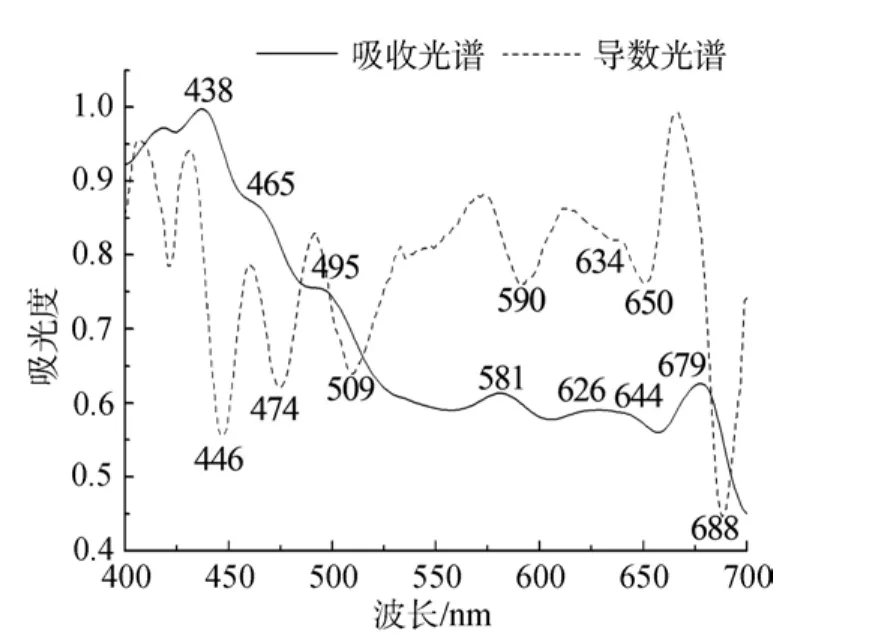
圖2 類囊體膜室溫吸收及其導數光譜Fig.2 Absorption and derivative spectra of thylakoids at room temperature
2.2 藻體室溫熒光光譜
如圖3A藻體的室溫發射光譜中, 當給予435 nm(對應 Chl a的吸收)、460 nm(對應 Chl c的吸收)激發光時, 均只發射源于PSII的Chl a的686~687 nm附近熒光, 沒有640 nm附近源于Chl c的熒光發射, 說明活體狀態下Chl c吸收的能量幾乎100%傳遞給Chl a。Chl c作為輔助色素通常與 Chl a及膜上蛋白結合,組裝形成Chl a/c-捕光蛋白復合物, 圍繞于反應中心外部, 上述結果說明, 膜上脂溶性的 Chl-蛋白復合物與反應中心之間的能量傳遞效率是極高的。當給予 580 nm(對應于藍隱藻 PC吸收)的激發光時, 在686 nm產生一個屬于Chl a的熒光發射。說明PC吸收的能量可以傳遞給Chl a。盡管Chl a在580 nm附近也有一小部分吸收, 但 580 nm 激發光所產生的Chl a熒光發射強度遠遠高于435 nm激發光(相對熒光強度之比可達1.5~1.8倍), 表明這部分Chl a熒光發射主要來自于PC的激發而非Chl a。此外, 580 nm光激發下還產生1個662~667 nm的源自PC的特征熒光峰, 其相對熒光強度接近甚至超過Chl a的發射峰(F667/F686=1.05~1.2), 說明藻體內的 PC可以把吸收的能量傳遞給Chl a, 但在測定時的激發光強度下, 也有超過半數的激發能以剩余熒光形式發射掉了。由于藻膽蛋白是在沒有足夠用受體的情況下才會以發射熒光的方式消耗激發能, 而據報道隱藻藻膽蛋白與 Chl a之間的能量傳遞效率很高, 可達98%~99%[7], 因此活體細胞中發射這部分660~667 nm剩余熒光的 PC應該是以某種方式中斷了與光合系統之間的能量傳遞關系。
激發光譜反映的是熒光物質在不同波長的激發光照射下, 對所產生的發射波長的熒光強度的貢獻,往往與該物質的吸收光譜中的峰形變化一致, 可以反映色基之間的能量偶聯情況[12]。如圖3B所示, 在700 nm發射波長下, 藻體細胞的激發光譜與吸收光譜很相似, 在藍區436、458和495 nm(導數光譜447、473和502 nm)處可看到小的源自于Chl a、Chl c和類胡蘿卜素的激發峰, 只有導數處理后的結果中可看到位于679 nm的Chl a紅區激發峰, 說明Chl c及類胡蘿卜素和Chl a之間存在能量偶聯, 只是對該熒光貢獻較小。活體細胞激發譜中, 最強的582 nm和646 nm(導數光譜592和662, 631 nm對應于625 nm吸收肩峰)的激發峰顯然源自 PC, 說明藻體內對700 nm長波熒光發射貢獻比例最高的是 PC。在520~550 nm(導數光譜529 nm和543 nm)的變化, 據報道與PC內部偶聯的色基對相互作用有關[13-14]。
2.3 藻體77K熒光光譜
室溫下通常只能觀測到屬于PSII的熒光, PSI在室溫下不產生顯著的熒光峰[15], 因而采用 77K液氮低溫條件測定, 以區分 PSI和 PSII的差別。活體細胞 77K低溫熒光發射譜與室溫相比, 峰形、峰位和相對強度都有所變化(圖3C), 其譜峰十分特別, 表現在以下幾個方面: (1)當給予435 nm和460 nm激發光時, 發射主峰在屬于Chl a的694~696 nm附近, 與高等植物和綠藻相似, 是源自于 PSII內周天線的典型發射峰[16]。此結果與以往對隱藻的報道不同, 含有PC的藍隱藻ChroomonasCCMP270以及含有PE的RhodomonasCS24的活體細胞在 77K低溫下屬于PSII的熒光峰都在683或者686 nm處, 這一現象被認為是由于隱藻 PSII光合系統特異而導致的[7,17],但作者的測定結果說明, 不同種屬的隱藻, 其 PSII的熒光特性可能是多樣化的。在這兩種激發波長下,700 nm以上的長波發射基本看不到, 說明Chl a和Chl c激發后的能量似乎更傾向于傳遞給PSII或者短波長Chl a。但Chl c激發后只產生Chl a的熒光, 說明77K低溫沒有影響Chl c向Chl a之間的能量傳遞效率。
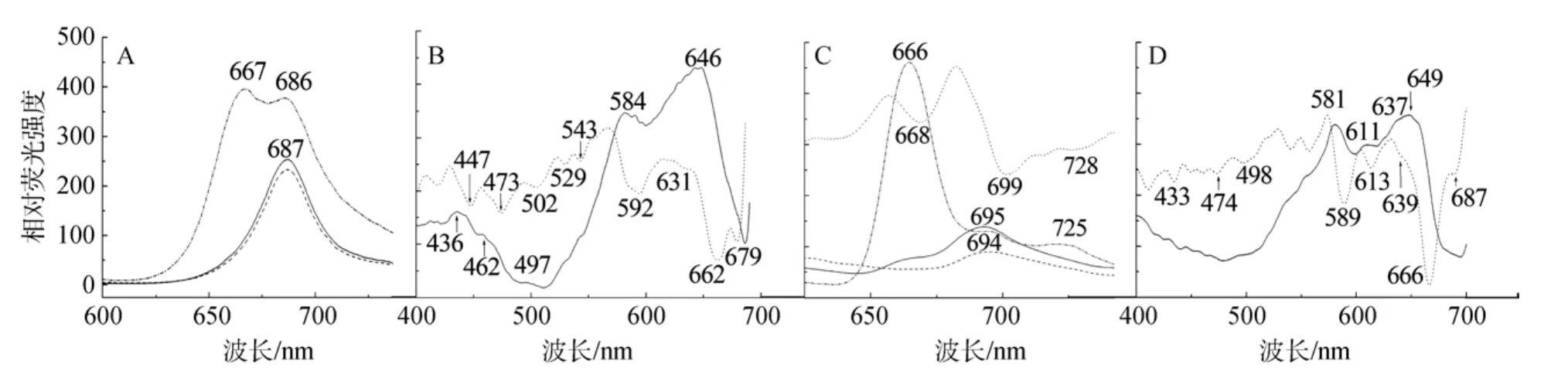
圖3 藻體細胞室溫(A, B)和77K(C, D)熒光發射(A, C), 激發(B, D)及其導數光譜Fig.3 Fluorescence emission (A, C), excitation (B, D) and derivative spectra of cells at room temperature (A, B) and77 K (C, D)
(2) 當給予580 nm對應于藍隱藻PC的激發光時, 除了產生695 nm源自PSII的熒光發射之外, 還產生 720~725 nm熒光, 該熒光發射被認為源于PSI[18-19]。此結果與Mimuro等[16]報道的不同隱藻的活體細胞中 PSI的 77K特征熒光發射峰通常位于715~730 nm相一致。說明藍隱藻完整藻體細胞中的PC可將激發能同時傳遞給PSII和PSI反應中心附近的Chl a。藻體中對700 nm以上長波熒光的貢獻主要來自于 PC, 說明 PC將相當一部分激發能傳遞給了長波Chl a。
(3) 580 nm光激發后的發射主峰位于 666 nm,是屬于 PC的特征發射峰, 該熒光峰比來自 PSI和PSII的 Chl a的發射峰高的多(F666/F695平均可達5.0, F666/F725=6.0), 而相對熒光發射強度是 Chl a激發時的4~5倍。與室溫熒光發射光譜相比較, 屬于PC的發射比例大幅度增加。藍隱藻細胞中這種發射超高比例藻膽蛋白熒光的現象與某些隱藻相似[7,17],但與藍藻或紅藻相比則顯得特異[20-21]。其原因可能是由于低溫導致了 PC與膜相關的能量傳遞途徑阻礙, 致使 PC激發后的大部分激發能沒有傳遞下去,表現為666 nm發射強度相對Chl a的發射強度大幅度升高。由于低溫條件下阻礙的是活體細胞內的生物過程或者動態過程, 而光物理和光化學過程仍可進行。推測生理條件下PC與膜組分之間的能量傳遞可能借助了某些低溫下可被阻斷的動態相互作用,而紅、藍藻藻膽體因為是附著在光合膜上的, 因而受低溫的影響相對要小。
PC激發對長波熒光的貢獻, 在發射波長為720 nm時藻體細胞 77K 激發光譜(圖3D)中體現得更為明顯。此時, 在440~470 nm之間屬于Chl a和Chl c的激發峰幾乎不可見, 只在導數處理的譜峰中可以看到687nm處對應于長波Chl a的激發貢獻。主要的激發峰在580~650 nm, 激發峰形與PC645的吸收譜圖很相似(圖1)。而77K下PC的激發光譜更精細,呈現581、611、632和 649 nm多個PC的激發峰, 在導數處理后更為明顯。對于隱藻PC645吸收光譜中3個典型的吸收譜峰與藻膽素色基之間的對應關系一直存在爭議, 但最近的一些報道傾向于認為,PC645所含有的色基吸收區域為: DBV(二氫膽綠素)?580~585 nm, MBV(中膽綠素)?610~622nm,PCB(藻藍膽素)?630~640 nm, PCB(β82)?645~650 nm,位于β亞基82位Cys上的PCB為最低能量色基[22-23]。因此上述4個激發峰應該分別對應于DBV(581 nm)、MBV(611 nm)、PCB(632 nm)和 PCBβ82(649 nm)。
2.4 類囊體膜室溫熒光光譜
葉綠體破碎之后制備的類囊體膜在室溫條件下以 435和 460 nm作為激發光照射時, 都只產生在688~689 nm附近的來自PSII的Chl a發射峰(圖4A),說明在分離得到的類囊體膜上Chl a和Chl c依然保持著良好的天然狀態, 吸收的光能均可有效的傳遞給Chl a。若以580 nm光激發時, 同樣在685~686 nm附近產生源于Chl a的熒光, 但相對于藻體細胞, 由于PC的流失, 相對熒光強度低于435 nm激發(相對強度比值為0.2~0.3), 甚至低于Chl c激發時的熒光(相對強度比值為0.7~0.8)。除此之外, 在659~660 nm處(比活體狀態藍移約5~6 nm)仍可看到有1個屬于PC的發射峰, 與Chl a峰的比值F660/F685= 1.4~1.6。說明經過充分洗滌的類囊體膜中仍有和 Chl-蛋白特異結合的PC。

圖4 類囊體膜在室溫(A, B)和77K(C, D)下的熒光發射(A, C), 激發(A, C)及導數光譜Fig.4 Fluorescence emission (A, C), excitation (B, D) and derivative spectra of thylakoid membrane at room temperature(A, B)and 77 K (C, D)
在 700 nm發射波長下的室溫激發光譜(圖4B)顯示, 在去除大部分PC的類囊體膜中, Chl a對此長波熒光的貢獻相對突出, 除了紅區435 nm激發峰外,在673 nm的藍區激發峰成為主峰; 但460 nm附近原屬于Chl c激發峰幾乎消失, 表明此時Chl c對此長波熒光的貢獻微乎其微; 而583, 632, 642 nm的激發峰明顯與 PC有關, 進一步說明在樣品中存在的 PC是與類囊體膜以良好的狀態結合著的, 且PC和Chl a之間依然保持著能量傳遞關系。
2.5 類囊體膜77K熒光光譜
在圖4C類囊體膜的77K熒光發射光譜中, 與活體細胞相似, 在435 nm和460 nm的激發光照射下,主要可使Chl a和Chl c激發, 此時類囊體膜的熒光發射主峰均為源自PSII內周天線Chl a的692 nm熒光(比活體狀態藍移約3~4 nm); 但在700 ~730 nm區域還呈現不明顯的肩峰區, 這一部分發射波長長于PSI的反應中心葉綠素P700, 因而一般認為來自PSI,說明類囊體膜中Chl a和Chl c激發后都可將能量傳遞給PSI。有報道認為, 隱藻PSI也有屬于自己的Chl a/c2捕光復合物 LHCI存在[24-25], 因此, 光譜中這一部分激發能可能來自LHCI中的Chl a和Chl c, 但是顯然份額較少。此外, 當給予類囊體膜580 nm的激發光時, 類囊體膜除了呈現比藻體時強度低的多的662 nm發射峰外, 還呈現出源自PSI的703、717 nm的長波Chl a的熒光峰(比藻體藍移7 ~8 nm), 再次說明PC和兩個光合系統的Chl a間均存在能量傳遞關系, 并且向長波Chl a傳遞的份額比Chl c更高。
圖4D類囊體膜的77K激發光譜中也顯示, 發射波長為720 nm時, 對這一長波熒光產生貢獻的主要是Chl a(431~448 nm、682 nm)和 PC(580~646 nm),Chl c(470 nm)以及類胡蘿卜素(494 nm)的激發不明顯。而 77K下 PC的激發光譜更精細, 除了 581和649 nm峰之外, 還呈現582、613、632和 646 nm處多個PC藻膽素色基的激發峰。
可見, 盡管類囊體膜制備過程中, 包裹在類囊體腔中的PC不可避免會流失, 但在實驗條件下所得的膜片上依然存在一定比例的 PC, 并且其上結合的Chl -蛋白復合物以及 PC的色基之間的能量傳遞關系仍與活體細胞狀態相似。
2.6 類囊體膜多肽SDS-PAGE
充分洗滌后的類囊體膜多肽分析結果見圖5。多肽的分布呈現一定的規律: 即主要分布在 20~22 ku(捕光復合物)、28~35 ku(反應中心)、45 ku 附近(PSII內周天線)和 66 ku附近(PSI核心)4個區域附近[25](圖5b)。結果經鋅染后在紫外光下觀察(圖5c), 發現在考染圖譜中豐度最高的 20 ku多肽附近有一條粉紅色熒光條帶, 與PC的β亞基遷移率相當(圖5d)。由于類囊體膜樣品在多肽組成分析之前經過丙酮/乙醇處理, 與蛋白質非共價結合的葉綠素和類胡蘿卜素被去除, 只有與蛋白質共價結合的藻膽素可被保留, 因此 PAGE圖譜中呈現粉紅色熒光的蛋白帶只能屬于含藻膽素的藻膽蛋白亞基, 說明類囊體膜樣品中確有PC存在。并且由于膜樣品在電泳前經過深度洗滌, 因此圖譜中觀察到的多肽組分(包括 PC)都應是附著或者結合于膜上的。多肽分析結果還顯示,PC645的β亞基總能被檢測到, 而α亞基則只有在銀染條件下偶爾可以見到(結果未顯示), 盡管α亞基可能因為分子量小而容易流失, 但這一結果暗示了PC645的β亞基似乎與膜結合更為緊密。既然目前的研究傾向于認為, PC645產生末端發射的基團(662 nm熒光發色團)是位于 β亞基的 PCBβ82, 激發能在PC645內部傳遞后最終到達PCBβ82, 后者再將能量傳遞給位于膜上的Chl a[22-23]。因此, 從功能上來說,PC645的β亞基應是與膜上葉綠素相互作用的位點,因此結構上由 β亞基與膜相接觸, 這一結果以往未有報道, 但作者認為應該是合理的。
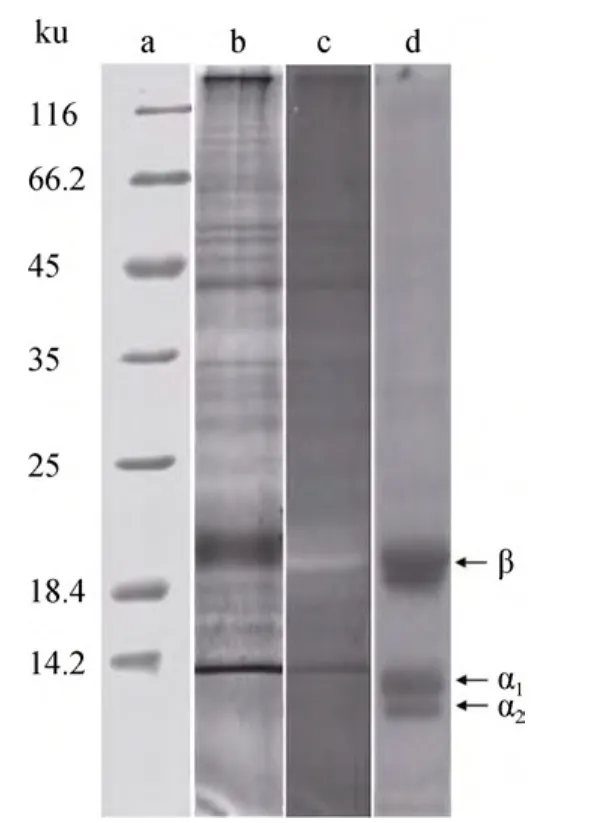
圖5 類囊體膜多肽以及PC亞基SDS-PAGE圖譜Fig.5 The SDS-PAGE profiles of thylakoid membrane peptides and PC subunits
3 討論與結論
目前對隱藻藻膽蛋白的了解遠不及紅、藍藻藻膽蛋白透徹, 而隱藻藻膽蛋白的研究又大多以游離的異二聚體或者更小的單體本身為對象[22-24,26-28],對于異二聚體之間的相互作用, 以及作為隱藻主要的捕光復合物與類囊體膜上 Chl-蛋白之間的接觸機制卻了解有限, 甚至于是否有結構上的接觸都還眾說紛紜, 存在爭議。
一方面, Wit等[7]提出的隱藻光合膜結構模型,是現有模型中最新的, 他們認為隱藻藻膽蛋白的α1α2ββ異二聚體形式是穩定的生理結構形式, 似乎不傾向于形成高度的聚合狀態; 藻膽蛋白異二聚體高密度均勻地填充于類囊體腔中, 彼此之間以及與膜上 Chl-蛋白復合物之間都沒有傾向性的結合; 二聚體蛋白可在不相互接觸、也不與膜接觸的情況下以 F?ster共振方式直接將吸收的能量在彼此之間傳遞, 并最終傳遞給位于類囊體膜上的Chl a。由于按照此模型計算, 藻膽蛋白與膜上能量的傳遞效率比活體狀態低得多, 因此 Doust等[8]認為, 離體狀態的藻膽蛋白其光譜學測定結果可能不能代表活體狀態下的準確情況。因此也說明處于游離狀態的隱藻藻膽蛋白似乎不是完全的天然狀態。
另一方面, Lichtlé等[25]曾經報道分到一種活性的 PE-葉黃素-PSII復合物組分; 而作者在前期的實驗中則分離到一種特殊的 PC-Chl a/c2-蛋白復合物,并且該復合物中 PC吸收的能量可直接傳遞給復合物內的Chl a, 無需經由Chl c2的介導[29]。這些結果直接說明, 隱藻藻膽蛋白在結構上是與膜上的 PSII或者Chl a/c-蛋白復合物直接相連的, 并且在功能上存在能量傳遞關系。
此外, 曾有活體細胞的熒光動力學研究顯示,隱藻Cryptomonas ovata細胞也存在不是很典型的光誘導的狀態轉變過程, 在PSII過激發時, 會導致PE與Chl a/c2以及PSII核心的偶聯中斷, 從而改變藻膽蛋白向PSII能量傳遞比率, 并且認為這是PSII的一種光保護機制[30]。隱藻類囊體膜雖然沒有基粒結構,但也有一定的垛疊[3], PSI和PSII的分布也是區域化的, 因此其激發能在兩個光系統之間的再分配以及光保護的實現, 可能有賴于藻膽蛋白與膜之間關系的某種動態變化。由于藻膽蛋白在類囊體腔內是密集存在的[7-8], 因此總有一部分藻膽蛋白不能接觸到類囊體膜的內表面。所以實現解除偶聯的方式有兩種可能: 一是藻膽蛋白為結合狀態的, 此時可通過某種結構或構象變化而與受體葉綠素解除偶聯[30-31], 比如一部分與類囊體膜的脫離; 另一種可能是在藻體細胞的天然狀態下就以游離狀態存在, 此時可能表現為 PSI或 PSII附近藻膽蛋白區域化的聚集程度的變化。
由于活體細胞中色基之間的能量傳遞過程非常復雜, 且實驗結論多有不同。因此, 本實驗改換思路,同時制備了充分洗滌但證實仍然結合有藻膽蛋白的類囊體膜為材料, 得到的實驗結果說明: (1)藍隱藻的類囊體膜上始終緊密的結合著一定量的 PC, 這樣的類囊體膜樣品在以往的報道中均沒有提及, 這一點充分說明PC不應是完全游離于膜腔內的; (2)SDSPAGE顯示, 深度洗滌過的類囊體膜上結合的主要是PC645的β亞基, 因此PC645有可能借助β亞基與膜接觸, 這一發現為近年來 PC645內部能量傳遞最終發射基團位于β亞基而非α亞基的觀點提供了直接的證據[22-23,26], 同時說明PC與膜結合時并非完全沒有傾向性; (3)PC既可將激發能傳遞給PSII, 但也可傳遞給PSI, 說明PC與膜上兩個光合系統之間都有功能上的關聯, 這結果與近年來的活體細胞時間分辨動力學研究一致[17,22]; (4)相較于Chl a和Chl c,PC在PSI長波熒光中的貢獻更大, 而Chl c吸收的光能, 更傾向于傳遞給PSII或者短波長Chl a; (5)藍隱藻細胞中表現出超比例的PC熒光發射, 說明PC在類囊體腔內應該有一部分是與膜不關聯而游離存在的,處于游離狀態下的 PC吸收的光能主要以發射自身熒光而耗散掉, 沒有將激發能全部傳遞給反應中心。
根據以上各方面的實驗結果, 作者提出隱藻藻膽蛋白在葉綠體中可能的存在和能量傳遞模式: 隱藻葉綠體內可能存在超過光合功能需要的藻膽蛋白,與其生長的深水中特定的弱光環境相適應; 藻膽蛋白在類囊體腔中與膜的結合狀態有可能是動態的,一部分為膜結合型, 另一部分為游離狀態(Morisset等[32]曾報道PC645可以形成不同的兩種晶型), 處于結合狀態的藻膽蛋白參與光能向反應中心的傳遞,而游離狀態的藻膽蛋白對于捕光功能來說可能暫時是冗余的, 而當需要的時候, 游離的藻膽蛋白還可與類囊體膜內表面重新結合或聚集; 藻膽蛋白與膜的結合部位包括PSI和PSII的定位區, 與PSII的接觸和能量傳可能通過LHCII中的Chl a/c蛋白復合物介導, 但能量是直接傳遞給Chl a而不經過Chl c的中轉, Chl c和藻膽蛋白向反應中心供能時各自遵循不同的能量傳遞路徑; 而藻膽蛋白在與 PSI的接觸中, 則更多地以某種方式與含有長波Chl a的組分相互作用; 藻膽蛋白可借助與膜的結合和脫離的動態方式, 以類似于高等植物的移動天線 LHCII的原理,調節激發能在光合系統之間的傳遞比例, 或者實現對反應中心的光保護。
[1] Watanabe M, Ikeuchi M.Phycobilisome: architecture of a light-harvesting supercomplex[J].Photosynth Res,2013, 116(2-3): 265-276.
[2] Glazer A N, Wedemayer G J.Cryptomonad biliproteinsan evolutionary perspective[J].Photosynth Res, 1995,46(1-2): 93-105.
[3] Gantt E, Edwards M R, Provasoli L.Chl oroplast structure of the Cryptophyceae.Evidence for phycobiliproteins within intrathylakoidal spaces[J].J Cell Biol, 1971,48(2): 280-290.
[4] Ludwig M, Gibbs S P.Localization of phycoerythrin at the lumenal surface of the thylakoid membrane inRhodomonas lens[J].J Cell Biol, 1989, 108(3): 875-884.
[5] Lichtlé C, Mckay R M L, Gibbs S P.Immunogold localization of photosystem I and photosystem II light-harvesting complexes in cryptomonad thylakoids[J].Biology of the Cell, 1992, 74(0): 187-194.
[6] Ingram K, Hiller R G.Isolation and characterization of a major Chl orophyll a/c2light-harvesting protein from aChroomonasspecies (Cryptophyceae)[J].Biochim Biophys Acta - Bioenergetics, 1983, 722(2): 310-319.
[7] van der Weij-De Wit C D, Doust A B, van Stokkum I H,et al.How energy funnels from the phycoerythrin antenna complex to photosystem I and photosystem II in cryptophyteRhodomonasCS24 cells[J].J Phys Chem B, 2006, 110(49): 25066-25073.
[8] Doust A B, Wilk K E, Curmi P M G, et al.The photophysics of cryptophyte light-harvesting[J].Journal of Photochemistry and Photobiology A: Chemistry,2006, 184(1-2): 1-17.
[9] Jeffrey S, Humphrey G.New spectrophotometric equations for determining Chl orophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton[J].Biochem Physiol Pflanzen, 1975, 167: 191-194.
[10] Laemmli U K.Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J].Nature, 1970, 227(5259): 680-685.
[11] Green B R, Parson W W.Advances in photosynthesis and respiration Volume13: Light-harvesting antennas in photosynthesis[M].The Netherlands: Kluwer Academic Publisher, 2003: 1-28.
[12] 丘冠英, 彭銀祥.生物物理學[M].武漢: 武漢大學出版社, 2000: 137-138.
[13] Maccoll R, Williams E C, Eisele L E, et al.Chromophore topography and exciton splitting in phycocyanin 645[J].Biochemistry, 1994, 33(21):6418-6423.
[14] Maccoll R, Eisele L E, Marrone J.Fluorescence polarization studies on four biliproteins and a bilin model for phycoerythrin 545[J].Biochim Biophys Acta,1999, 1412(3): 230-239.
[15] van der Weij-De Wit C D, Ihalainen J A, van Grondelle R, et al.Excitation energy transfer in native and unstacked thylakoid membranes studied by low temperature and ultrafast fluorescence spectroscopy[J].Photosynth Res, 2007, 93(1-3): 173-182.
[16] Mimuro M, Tamai N, Murakami A.Multiple pathways of excitation energy flow in the photosynthetic pigment system of a cryptophyte,Cryptomonassp.(CR-1)[J].Phycological Research, 1998, 46(3): 155-164.
[17] van der Weij-De W C, Doust A B, van Stokkum I H M,et al.Phycocyanin sensitizes both photosystem I and photosystem II in cryptophyteChroomonasCCMP270 cells[J].Biophys J, 2008, 94(6): 2423-2433.
[18] Wientjes E, van Stokkum I H, van Amerongen H, et al.Excitation-energy transfer dynamics of higher plant photosystem I light-harvesting complexes[J].Biophys J,2011, 100(5): 1372-1380.
[19] Kuang T Y, Argyroudi-Akoyunoglou J H, Nakatani H Y,et al.The origin of the long-wavelength fluorescence emission band (77 degrees K) from photosystem I[J].Arch Biochem Biophys, 1984, 235(2): 618-627.
[20] Mcconnell M D, Koop R, Vasil'Ev S, et al.Regulation of the distribution of Chl orophyll and phycobilinabsorbed excitation energy in cyanobacteria.A structure-based model for the light state transition[J].Plant Physiol, 2002, 130(3): 1201-1212.
[21] Joshua S, Mullineaux C W.Phycobilisome diffusion is required for light-state transitions in cyanobacteria[J].Plant Physiol, 2004, 135(4): 2112-2119.
[22] Marin A, Doust A B, Scholes G D, et al.Flow of excitation energy in the cryptophyte light-harvesting antenna phycocyanin 645[J].Biophys J, 2011, 101(4):1004-1013.
[23] MacColl R, Williams E C, Eisele L E, et al.Chromophore topography and exciton splitting in phycocyanin 645[J].Biochemistry, 1994, 33(21):6418-6423.
[24] Janssen J, Rhiel E.Evidence of monomeric photosystem I complexes and phosphorylation of Chl orophyll a/c-binding polypeptides inChroomonassp.strain LT (Cryptophyceae) [J].Int Microbiol, 2008,11(3): 171-178.
[25] Lichtlé C, Duval J C, Lemoine Y.Comparative biochemical, functional and ultrastructural studies of photosystem particles from a Cryptophycea:Cryptomonas rufescens;isolation of an active phycoerythrin particle[J].Biochim Biophys Acta (BBA) - Bioenergetics,1987, 894(1): 76-90.
[26] Novoderezhkin V I, Doust A B, Curutchet C, et al.Excitation dynamics in phycoerythrin 545: modeling of steady-state spectra and transient absorption with modified redfield theory[J].Biophys J, 2010, 99(2):344-352.
[27] Collini E, Wong C Y, Wilk K E, et al.Coherently wired light-harvesting in photosynthetic marine algae at ambient temperature [J].Nature, 2010, 463: 644-647.
[28] Huo P F, Coker D F.Theoretical study of coherent excitation energy transfer in cryptophyte phycocyanin 645 at physiological temperature [J].J Phys Chem Lett,2011(5): 825-833.
[29] Chen M, Li S H, Sun L.A novel phycocyanin-Chl a/c2-protein complex isolated from Chl oroplasts ofChroomonas placoidea[J].Chinese Chemical Letters,2007, 18: 1374-1378.
[30] Synder U K, Biggins J.Excitation-energy redistribution in the cryptomonad algaCryptomonas ovata[J].Biochim Biophys Acta - Bioenergetics, 1987, 892:48-55.
[31] Bruce D, Biggins J, Steiner T.Excitation energy transfer in the cryptophytes.Fluorescence excitation spectra and picosecond time-resolved emission spectra of intact algae at 77K [J].Photochem Photobiol, 1986,44(4): 425-554.
[32] Morisset W, Wehrmeyer W, Schirmer T, et al.Crystallization and preliminary x-ray diffraction data of the cryptomonad biliprotein phycocyanin-645 from aChroomonasspec[J].Archives of Microbiology, 1984,140(2-3): 202-205.
