PRRSV新亞型毒株JL580的基因組變異特征研究
2015-04-27 00:49:48常曉博趙款葉超姜成剛王淑杰蔡雪輝張輝安同慶
中國(guó)獸藥雜志 2015年5期
常曉博,趙款,葉超,姜成剛,王淑杰,蔡雪輝,張輝,安同慶?
(1.吉林農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院,長(zhǎng)春130118;2.中國(guó)農(nóng)業(yè)科學(xué)院哈爾濱獸醫(yī)研究所獸醫(yī)生物技術(shù)國(guó)家重點(diǎn)實(shí)驗(yàn)室,哈爾濱150001)
PRRSV新亞型毒株JL580的基因組變異特征研究
常曉博1,2,趙款2,葉超2,姜成剛2,王淑杰2,蔡雪輝2,張輝1?,安同慶2?
(1.吉林農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院,長(zhǎng)春130118;2.中國(guó)農(nóng)業(yè)科學(xué)院哈爾濱獸醫(yī)研究所獸醫(yī)生物技術(shù)國(guó)家重點(diǎn)實(shí)驗(yàn)室,哈爾濱150001)
為了解豬繁殖與呼吸綜合征病毒(PRRSV)新分離株JL580遺傳變異特征,研究擴(kuò)增得到了該病毒的全基因組序列,并對(duì)GP5和NSP2序列進(jìn)行了比對(duì)分析。結(jié)果表明:JL580株屬于美洲株,且不同于中國(guó)現(xiàn)流行的其他毒株,與中國(guó)代表毒株CH-1a的核苷酸同源性是84.9%,與HP-PRRSV代表株JXA1、HuN4、TJ的核苷酸同源性分別是84.8%、84.9%、85%。對(duì)基因組的進(jìn)一步分析表明,JL580株GP5蛋白與CH-1a、VR-2332、HuN4相比,R13→Q13,V27→A27,H38→N38,R151→K151;且NSP2存在3處缺失,與HP-PRRSV的30個(gè)氨基酸的缺失有明顯的不同。JL580分離株的出現(xiàn)表明中國(guó)出現(xiàn)了新的PRRSV亞型,這對(duì)于PRRSV的防治和新疫苗的研制提供了科學(xué)依據(jù)。
豬繁殖與呼吸綜合征病毒;JL580;基因組;遺傳變異
豬繁殖與呼吸綜合征(porcine reproductive and respiratory syndrome,PRRS)是由豬繁殖與呼吸綜合征病毒(PRRS virus,PRRSV)引起的豬的一種急性、高度接觸性傳染病。PRRS于1987年[1]首次發(fā)生于美國(guó)的衣阿華、北卡羅來納等州,隨后迅速在美洲蔓延,我國(guó)于1996年[2]首次分離到PRRSV,目前該病在全球范圍內(nèi)暴發(fā)和流行[3]。PRRSV有兩種基因型,分別為歐洲型(EU-type)和北美洲型(NA-type),我國(guó)主要流行毒株為北美洲型[4]。PRRSV在自然界中不斷變異,不斷出現(xiàn)新的病毒株,給PRRS的防治帶來了巨大的挑戰(zhàn)。PRRSV基因組中ORF5是高變區(qū)之一,其編碼的蛋白是PRRSV主要的宿主保護(hù)性抗原蛋白[5],PRRSV的變異可以導(dǎo)致PRRS疫苗株交叉保護(hù)率低下。編碼PRRSV的非結(jié)構(gòu)蛋白基因中,位于ORF1a的NSP2基因最易發(fā)生變異,主要表現(xiàn)為氨基酸的突變、缺失和插入。2006年,我國(guó)南方暴發(fā)的HP-PRRSV基因組中NSP2存在30個(gè)不連續(xù)氨基酸的缺失[6];Darwich等[7]通過分析PRRSV基因I型毒株的基因和免疫多樣性發(fā)現(xiàn)他們所研究的基因I型PRRSV中NSP2存在74個(gè)氨基酸的缺失。因此,針對(duì)GP5和NSP2的研究有助于了解整個(gè)基因組的遺傳衍化特征。
2013年,吉林省某豬場(chǎng)發(fā)生疑似PRRS,臨床表現(xiàn)為高熱,耳朵發(fā)紺,24 h全身發(fā)紺,最后死亡,病程較短,發(fā)病率較高。通過病料采集、檢測(cè)及病毒的分離鑒定,我們將新分離株命名為JL580。為弄清JL580新分離株的基因組變異特征,特別是ORF5和NSP2的變異規(guī)律,我們著重對(duì)JL580基因組中的GP5和NSP2進(jìn)行了序列比對(duì)分析,從而了解JL580株的遺傳變異情況。
1 材料和方法
1.1 樣品和細(xì)胞 樣品:采自吉林省某豬場(chǎng)疑似PRRSV感染豬的肺臟和淋巴結(jié);細(xì)胞:Marc-145細(xì)胞由中國(guó)農(nóng)業(yè)科學(xué)院哈爾濱獸醫(yī)研究所動(dòng)物疫病診斷及技術(shù)服務(wù)中心提供。
1.2 主要試劑 QIAamp Viral RNA Mini Kit購自QIAGEN公司;Reverse Transcriptase M-MLV、Recombinant RNase Inhibitor、TaKaRa EX Taq Hot Start Version、rTaq DNA聚合酶、3’-Full RACE Core Set Ver.2、Clontech SMARTerTMRACE cDNA Amplification Kit、pMD18-T Vector Kit、E.coli DH5a感受態(tài)均購自寶生物工程(TaKaRa)有限公司;膠回收試劑盒購自O(shè)MEGA公司。
1.3 全基因組分析
1.3.1 引物及參考序列 根據(jù)GenBank中已登錄的國(guó)內(nèi)外經(jīng)典毒株序列的保守區(qū)及GP5測(cè)序結(jié)果,參考美國(guó)NADC30株[8](GenBank no.JN654459)設(shè)計(jì)并合成9對(duì)引物(表1)。

表1 JL580全基因組序列擴(kuò)增所需引物
1.3.2 RNA提取與PCR擴(kuò)增、克隆及測(cè)序 按照QIAamp Viral RNA Mini Kit操作手冊(cè)提取病毒總RNA,反轉(zhuǎn)錄成cDNA后用于目的片段的擴(kuò)增,基因5’端和3’端序列分別參照Clontech SMARTerTMRACE cDNA Amplification Kit和3’-Full RACE Core Set Ver.2使用說明書進(jìn)行擴(kuò)增。利用膠回收試劑盒回收PCR產(chǎn)物后分別與pMD18-T載體16℃過夜連接,轉(zhuǎn)化并篩選重組陽性質(zhì)粒,每個(gè)片段選取5個(gè)單克隆菌落,重復(fù)進(jìn)行3次測(cè)序。
1.4 全基因組序列的分析 應(yīng)用DNAStar軟件中的Seqman程序拼接序列,Megalign進(jìn)行序列比對(duì)及同源性分析;應(yīng)用MEGA5.2軟件對(duì)該毒株的全基因組序列進(jìn)行遺傳進(jìn)化分析。
2 結(jié)果
2.1 JL580全基因組序列擴(kuò)增結(jié)果 將JL580病毒提取的RNA反轉(zhuǎn)錄成cDNA后,利用9對(duì)引物對(duì)其進(jìn)行擴(kuò)增,得到9條與預(yù)期大小相符的片段(圖1)。將測(cè)序所獲得的9個(gè)首尾重疊的目的基因片段進(jìn)行拼接,測(cè)定的基因組全長(zhǎng)為15019 bp。

圖1 PRRSV全基因組PCR擴(kuò)增產(chǎn)物
2.2 JL580全基因組序列分析結(jié)果 遺傳進(jìn)化樹(圖2)結(jié)果顯示,JL580株和NADC30株不同于CH-1a株、VR-2332株和HuN4株,它們形成了一個(gè)新的獨(dú)立分支。JL580分離株與18株國(guó)內(nèi)外參考株(表2)相比,核苷酸同源性在83.2%~92.9%,氨基酸同源性在70.8%~83.1%。與美國(guó)NADC30株相比核苷酸同源性為92.9%,氨基酸同源性為83.1%;與經(jīng)典株CH-1a、VR-2332株的核苷酸同源性為84.9%和84.3%,推導(dǎo)的氨基酸同源性為74.1%和72.9%;與HP-PRRSV中的代表株HuN4、JXA1、TJ株的核苷酸同源性分別為84.9%、84.8%、85%,氨基酸同源性為75.7%、75.3%、75.7%。
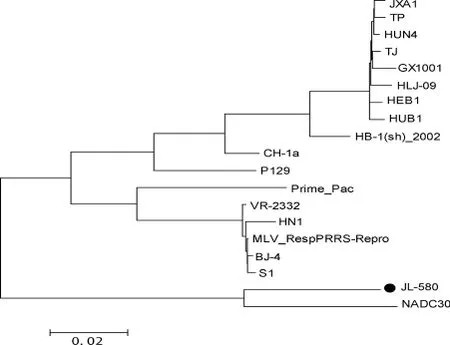
圖2 PRRSV全基因組核苷酸序列的系統(tǒng)進(jìn)化樹
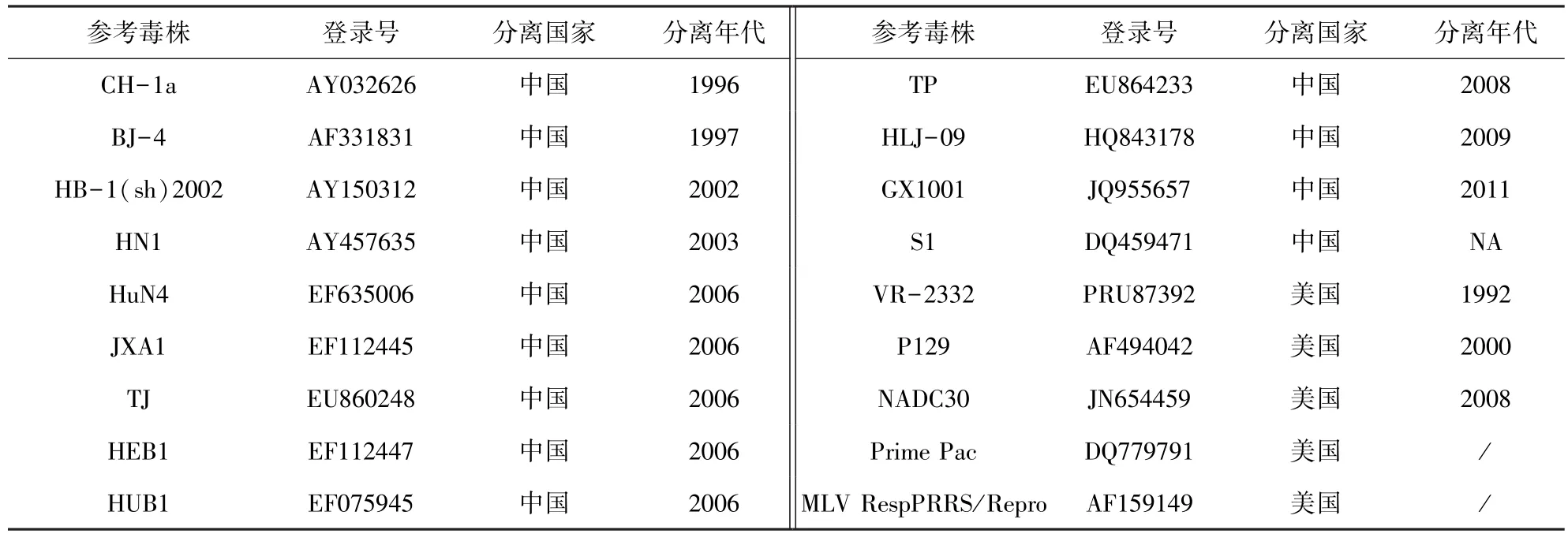
表2 參考毒株信息
2.3 JL580株ORF5遺傳特征分析 JL580株與CH-1a株相比,GP5核苷酸同源性為87.4%,氨基酸同源性為86.1%,與HP-PRRSV中的HuN4毒株相比,核苷酸同源性為85.6%,氨基酸同源性為84.6%。此外,JL580病毒株GP5蛋白與CH-1a、VR-2332、HuN4相比發(fā)生了顯著變異,主要變異位點(diǎn)分別是R13→Q13,V27→A27,H38→N38,R151→K151。
2.4 JL580株NSP2遺傳特征分析 JL580株與經(jīng)典株CH-1a株相比,NSP2基因核苷酸同源性為72.2%,氨基酸同源性為68.6%,與HuN4株相比核苷酸同源性為71.5%,氨基酸同源性為69.1%(表3)。JL580分離株存在3處連續(xù)堿基的缺失,分別位于基因組2304~2636、2779~2781和2852~2908處;這3處缺失導(dǎo)致NSP2編碼的蛋白在322~432位缺失111個(gè)氨基酸、481位缺失1個(gè)氨基酸、504~522位缺失19個(gè)氨基酸(圖3)。

表3 JL580株GP5和NSP2與部分毒株氨基酸同源性分析

圖3 NSP2氨基酸序列比對(duì)
3 討論
豬繁殖與呼吸綜合征自1995年在我國(guó)發(fā)現(xiàn)以來已有20年,雖然已研制多種疫苗防治,但PRRS并未根除。PRRSV為單股正鏈RNA病毒[9],在自然界以及免疫壓力[10]下不斷變異。2006年北美[11]和亞洲出現(xiàn)的高致病性PRRSV表明病毒的潛在變異導(dǎo)致PRRSV出現(xiàn)了新的變異株。本研究通過對(duì)JL580分離株進(jìn)行分子流行病學(xué)及遺傳進(jìn)化分析發(fā)現(xiàn),JL580病毒株屬于北美洲型PRRSV,并且與國(guó)內(nèi)PRRSV經(jīng)典株CH-1a和HP-PRRSV株HuN4處于遺傳進(jìn)化樹的不同分支中,同源性較低;而與美國(guó)NADC30株的全基因組序列具有較高的同源性。從JL580與國(guó)內(nèi)之前報(bào)道的經(jīng)典PRRSV和HP-PRRSV的遺傳距離上來看,JL580屬于國(guó)內(nèi)出現(xiàn)的一個(gè)新亞型毒株。本研究結(jié)果表明我國(guó)PRRSV出現(xiàn)了新的遺傳分支,揭示了我國(guó)PRRSV存在著更為復(fù)雜的遺傳多樣性。
GP5是PRRSV的主要結(jié)構(gòu)蛋白,也是最易發(fā)生變異的蛋白,R13和R151是毒力相關(guān)位點(diǎn)[12],非中和表位A(aa27~aa30)和中和表位B(aa37~aa45)是影響免疫的關(guān)鍵位點(diǎn)。研究發(fā)現(xiàn)JL580株GP5蛋白與CH-1a、VR-2332、HuN4相比,糖基化位點(diǎn)并未發(fā)生變異,但是中和表位、非中和表位及毒力位點(diǎn)發(fā)生了顯著的變異,主要表現(xiàn)在R13→Q13,V27→A27,H38→N38,R151→K151。NSP2是PRRSV非結(jié)構(gòu)蛋白中最易發(fā)生變異的蛋白,歐洲型和北美洲型NSP2高變區(qū)氨基酸缺失的報(bào)道[13]表明NSP2對(duì)于病毒的復(fù)制不具有決定性作用但在宿主免疫方面可能具有非常重要的作用。JL580株NSP2編碼的蛋白存在3處不連續(xù)氨基酸的缺失,與經(jīng)典株CH-1a相比NSP2共缺失了131個(gè)氨基酸;與2006年暴發(fā)的HP-PRRSV基因組中NSP2存在30個(gè)不連續(xù)氨基酸的缺失相比,JL580株NSP2基因編碼的蛋白共缺失101個(gè)氨基酸,除此之外,JL580株和HuN4株在481位具有一個(gè)相同的缺失位點(diǎn)。另外,JL580病毒株NSP2的缺失情況與2006年Han等[11]和2014年Choi等[14]所研究的PRRSV分離株NSP2缺失131個(gè)氨基酸情況相似。因此,JL580病毒株是目前國(guó)內(nèi)存在的一種新的變異株,其GP5和NSP2基因的變異相對(duì)于以往國(guó)內(nèi)發(fā)現(xiàn)的變異株具有顯著的差異性。至于JL580新分離株GP5中和表位、非中和表位及毒力位點(diǎn)的變異和基因組中3處不連續(xù)氨基酸的缺失是否會(huì)影響其免疫原性和致病性,還有待進(jìn)一步研究。
[1] Keffaber K K.Reproductive failure of unknown etiology[J].Am Assoc Swine Pract Newslett,1989,(2):1-9.
[2] 郭寶清,陳章水,劉文興,等.從疑似PRRSV感染的流產(chǎn)胎兒分離PRRSV的研究[J].中國(guó)畜禽傳染病,1996,2:1-4.
[3] Rowland R R,Lunney J,Dekkers J.Control of porcine reproductive and respiratory syndrome(PRRS)through genetic improvements in disease resistance and tolerance[J].Front Genet,2012,3:260.
[4] 朱佳毅,任曉峰.豬繁殖與呼吸綜合征病毒檢測(cè)方法的研究進(jìn)展[J].中國(guó)預(yù)防獸醫(yī)學(xué)報(bào),2014,36(1):77-80.
[5] Pirzadeh B,Dea S.Monoclonal antibodies to the ORF5 product of porcine reproductive and respiratory syndrome virus define linear neutralizing determinants[J].J Gen Virol,1997,78(8):1867-1873.
[6] Zhou Y J,Hao X F,Tian Z J,et al.Highly pathogenic porcine reproductive and respiratory syndrome virus emerged in China[J].Transbound Emerg Dis,2008,55(3/4):152-164.
[7] Darwich L,Gimeno M,Sibila M,et al.Genetic and immunobiological diversities of porcine reproductive andrespiratorysyndrome genotype I strains[J].Vet Microbiol,2011,150(1/2):49-62.
[8] Brockmeier S L,Loving C L,Vorwald A C,et al.Genomic sequence and virulence comparison of four type 2 porcine reproductive and respiratory syndrome virus strains[J].Virus Res,2012,169(1):212-221.
[9] Rossow K D.Porcine reproductive and respiratory syndrome[J].Vet Pathol,1998,35(1):1-20.
[10]Liu D,Zhou R,Zhang J,et al.Recombination analyses between two strains of porcine reproductive and respiratory syndrome virus in vivo[J].Virus Res,2011,155(2):473-486.
[11]Han J,Wang Y,F(xiàn)aaberg K S.Complete genome analysis of RFLP 184isolatesofporcinereproductiveandrespiratory syndrome virus[J].Virus Res,2006,122(1/2):175-182.
[12]Allende R,Kutish G F,Laegreid W,et al.Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype[J].Arch Virol,2000,145(6):1149-1161.
[13]Rascon-Casteio E,Burgara-Estrella A,Mateu E,et al.Immunological features of the non-structural proteins of porcine reproductive and respiratory syndrome virus[J].Viruses,2015,7(3):873-886.
[14]Choi H W,Nam E,Lee Y J,et al.Genomic analysis and pathogenic characteristics of type 2 porcine reproductive and respiratory syndrome virus nsp2 deletion strains isolated in Korea[J].Vet Microbio,2014,170(3/4):232-245.
(編輯:李文平)
Genetic Diversity of Porcine Reproductive and Respiratory Syndrome Virus of New Subtype JL580 Strain
CHANG Xiao-bo1,2,ZHAO Kuan2,YE Chao2,JIANG Cheng-gang2,WANG Shu-jie2,CAI Xue-hui2,ZHANG Hui1?,AN Tong-qing2?
(1.College of Animal Science and Technology,Jilin Agricultural University,Changchun 130118,China;2.State Key Laboratory of Veterinary Biotechnology,Harbin Veterinary Research Institute,Chinese Academy of Agricultural Sciences,Harbin 150001,China)
To investigate the variation of pathogenic porcine reproductive and respiratory syndrome(PRRSV)of JL580,the full-length genomic sequence of the virus was amplified,and the GP5 and NSP2 gene were analyzed,respectively.Results showed that JL580 strain shared 84.9%nucleotide similarity of complete sequence with CH-1a strain,84.8%,84.9%,85%compared with JXA1,HuN4 and TJ strains,and clearly different from other prevalent strains in China,which belongs to North American genotype PRRSV.The GP5 protein sequence was variable in the newly isolated PRRSV when compared to CH-1a,VR-2332,HuN4,which were R13→Q13,V27→A27,H38→N38,R151→K151.In addition,the full-length genomic sequence of the virus revealed three distinct deletions in NSP2 which had significant difference with the 30 amino acid of HP-PRRSV.Therefore,the study of JL580 indicated that new subtype and new branch of PRRSV have emerged in China,which would provide scientific basis for the development of new vaccine and comprehensive prevention and control of PRRSV.
PRRSV;JL580;genome;genetic variation
2015-02-06
A
1002-1280(2015)05-0012-05
S858.28
國(guó)家自然科學(xué)基金(31270045);吉林省科技廳基礎(chǔ)研究項(xiàng)目(20140101028JC)
常曉博,碩士研究生,從事PRRSV分子流行病學(xué)研究。
張輝,E-mail:huizhang01525@163.com;安同慶,E-mail:antongqing@163.com
