Fusarium mangiferae侵染對(duì)凱特杧內(nèi)源激素含量的影響
2015-05-30 02:33:24柳鳳歐雄常詹儒林韋繼光常金梅
熱帶作物學(xué)報(bào) 2015年7期
柳鳳 歐雄常 詹儒林 韋繼光 常金梅
摘 要 為探討杧果感染畸形病過(guò)程中頂芽?jī)?nèi)源激素含量的變化規(guī)律,以凱特杧為試驗(yàn)材料,測(cè)定了接種病菌(Fusarium mangiferae)后頂芽?jī)?nèi)生長(zhǎng)素(IAA)、赤霉素(GA3)和脫落酸(ABA)含量的變化情況。結(jié)果表明:接種病菌后,杧果頂芽?jī)?nèi)赤霉素和生長(zhǎng)素含量急速上升,且在整個(gè)試驗(yàn)階段明顯高于健康處理,脫落酸含量在試驗(yàn)后期不斷升高;分析發(fā)現(xiàn),F(xiàn). mangiferae侵染能夠顯著影響杧果頂芽?jī)?nèi)源激素的含量,改變IAA/ABA和GA3/ABA的平衡,致使杧果的生長(zhǎng)發(fā)育受到影響,進(jìn)而引發(fā)畸形病的產(chǎn)生。
關(guān)鍵詞 杧果;畸形病;Fusarium mangiferae;內(nèi)源激素
中圖分類號(hào) S667.7 文獻(xiàn)標(biāo)識(shí)碼 A
Abstract In order to study the effect of pathogens causing malformation on the endogenous hormone contents in Mangifera indica Keitt, we detected the changes of IAA, GA3 and ABA contents in buds affected by Fusarium mangiferae. The results showed that the contents of endogenous IAA and GA3 rose rapidly and were significantly higher than the control at at whole period after inoculation; ABA content rose at the late period after inoculation. The pathogens could affect the contents of IAA, GA3 and ABA in mango significantly and disturbed the balance of IAA/ABA and GA3/ABA, which affected the growth and development of mango, related to the mango malformation disease causing.
Key words Mangifera indica; Mango malformation; Fusarium mangiferae; Endogenous hormone
doi 10.3969/j.issn.1000-2561.2015.07.009
病原菌的入侵會(huì)強(qiáng)烈干擾植物體的正常代謝,內(nèi)源激素作為植物體內(nèi)重要的微量信號(hào)分子,必不可少的參與此過(guò)程,內(nèi)源激素作為抗性因子,啟動(dòng)或抑制與抗病相關(guān)蛋白的基因表達(dá),起到誘導(dǎo)或阻遏抗病性的產(chǎn)生,最終表現(xiàn)出感病或抗病[1-3]。脫落酸(ABA)、赤霉素(GA3)和生長(zhǎng)素(IAA)是調(diào)節(jié)植物細(xì)胞生長(zhǎng)代謝的重要內(nèi)源激素,在植物生長(zhǎng)發(fā)育中充當(dāng)著重要而又復(fù)雜的角色,其含量及變化規(guī)律決定著細(xì)胞與組織的生長(zhǎng)狀況[4]。正常生長(zhǎng)情況下,這些內(nèi)源激素之間處于某種動(dòng)態(tài)平衡狀態(tài),調(diào)節(jié)植物的生長(zhǎng)發(fā)育[5]。病原物的入侵打破了植物體內(nèi)各種內(nèi)源激素之間原有的平衡狀態(tài),改變了各種內(nèi)源激素之間的比值,從而使植株生長(zhǎng)量減小,生長(zhǎng)勢(shì)弱,很快發(fā)生病變,甚至衰亡[6-7]。杧果畸形病是一種危害杧果正常生長(zhǎng)的世界性病害,植株感病后嫩葉變細(xì)而脆,嫩芽、花序簇生,最后干枯,因此杧果染病后幾乎不座果。國(guó)內(nèi)該病主要分布在云南、四川海拔較高的杧果晚熟地區(qū)[8-10]。有研究報(bào)道患病組織的形成與杧果內(nèi)源激素的失調(diào)有著十分密切的關(guān)系,但關(guān)于病原菌接種后杧果內(nèi)內(nèi)源激素含量的變化情況目前還無(wú)人測(cè)定[11]。本研究以中國(guó)杧果晚熟地區(qū)主栽品種‘凱特杧為材料,通過(guò)比較人工接種F. mangiferae后,健康與接種杧果內(nèi)內(nèi)源激素含量的差別,旨在闡明F. mangiferae侵染對(duì)杧果內(nèi)源激素含量變化的影響,豐富杧果畸形病病原菌的致病機(jī)理研究。
1 材料與方法
1.1 材料
試驗(yàn)設(shè)病原菌接種處理和健康(對(duì)照)兩組。供試MG06菌株為杧果畸形病病原菌F. mangiferae。以健康的一年生凱特杧實(shí)生苗為材料,采用組織塊接種法接種。將病菌在PDA培養(yǎng)基上培養(yǎng)7 d后,菌落邊緣打孔,取直徑0.8 cm的菌餅作為接種體。利用解剖刀在頂芽下1~2 cm處刺傷,將菌餅菌絲面貼于刺傷處,保濕培養(yǎng)。每間隔5 d接種1次,共接種5次作為平行試驗(yàn)。接種0、15、30、45、60、75、90和105 d后取樣。每處理每次取15個(gè)芽,在液氮冷凍中磨成粉狀進(jìn)行混勻,分開(kāi)包裝后置于-80 ℃冰箱中保存。
1.2 方法
1.2.1 內(nèi)源激素的提取與純化 每次取1.0 g杧果頂芽粉狀物加10 mL提取液(80%色譜級(jí)甲醇,內(nèi)含1 mmol/L的BHT二叔丁基對(duì)甲苯酚),4 ℃過(guò)夜后離心,沉淀中加5 mL提取液抽提2 h,收集所有的上清液測(cè)定總體積,37 ℃減壓蒸發(fā)甲醇,留水相,之后加0.2 g的PVPP吸附酚類及色素,震蕩10 min后離心取上清液,往上清液中加入等體積石油醚脫色2次,留水相,冰乙酸調(diào)pH至2.5~2.9,用等體積乙酸乙酯萃取3次,合并酯相,減壓蒸干,用2 mL甲醇溶解。過(guò)C-18柱固相萃取柱進(jìn)行純化,具體步驟是:80%甲醇(1 mL)平衡柱→上樣→收集樣品→移開(kāi)樣品后用100%甲醇(5 mL)洗柱→100%乙醚(5 mL)洗柱→100%甲醇(5 mL)洗柱→循環(huán)。將過(guò)柱后的樣品轉(zhuǎn)入5 mL塑料離心管中,真空濃縮干燥,加1.5 mL甲醇溶解,過(guò)0.45 μm有機(jī)系針頭式過(guò)濾器過(guò)濾,1.5 mL棕色液相上機(jī)瓶收集濾液,備用。
1.2.2 標(biāo)準(zhǔn)溶液的配制 將IAA、GA3、和ABA 3種標(biāo)準(zhǔn)樣用色譜級(jí)甲醇溶解,分別配置為100 mg/L標(biāo)準(zhǔn)溶液,再將上述3種溶液配置成混合標(biāo)準(zhǔn)溶液待用。
1.2.3 標(biāo)準(zhǔn)曲線的制備及回收率測(cè)定 將各種激素標(biāo)樣在1.2.4的色譜條件下分別進(jìn)樣,進(jìn)樣量分別為5、10、15、20、25、30和35 μL。記錄保留時(shí)間,以此確定為標(biāo)樣的出峰時(shí)間。采用峰面積外標(biāo)法定量測(cè)定,以濃度Y(mg/L)為縱坐標(biāo),峰面積X(mV/s)為橫坐標(biāo),計(jì)算得到各激素的標(biāo)準(zhǔn)曲線和相關(guān)系數(shù)。準(zhǔn)確稱取一定質(zhì)量的標(biāo)樣,按1.21樣品處理方法處理,根據(jù)加入量和檢出量計(jì)算回收率,3次平行試驗(yàn)驗(yàn)證。
1.2.4 色譜條件 色譜儀為島津(Shimadzu)高效液相色譜儀,包括LC-10ATvp高壓泵,SPD-10Avp檢測(cè)器,CTO-10ASvp柱溫箱,C-R8A積分儀;色譜柱:Agilent ZORBAX SB-C18(4.6×250 mm,5 μm);流動(dòng)相:乙腈-甲醇-0.6%乙酸(50 ∶ 500 ∶ 450);流速1 mL/min;柱溫35 ℃;檢測(cè)波長(zhǎng):254 nm;進(jìn)樣量10 μL。
1.3 試驗(yàn)數(shù)據(jù)的統(tǒng)計(jì)分析
數(shù)據(jù)的處理和分析均采用Microsoft Excel2003 軟件及SPSS10.0 for Windows統(tǒng)計(jì)軟件進(jìn)行統(tǒng)計(jì)分析和Duncans(鄧肯氏)多重差異比較,p<0.05為差異具有顯著性。
2 結(jié)果與分析
2.1 標(biāo)準(zhǔn)曲線的制備及回收率測(cè)定
在選定的色譜條件下測(cè)得的赤霉素(GA3)保留時(shí)間為(3.361±0.050)s,線性回歸方程為y=58.921x-19 489(R2=0.996 0),回收率為80.4%;生長(zhǎng)素(IAA)保留時(shí)間為(5.155±0.061)s,線性回歸方程為y=852.14x-31 248(R2=0.998 8)回收率為81.3%;脫落酸(ABA)保留時(shí)間為(7.374±0.049)s,線性回歸方程為y=498.2x-38 003(R2=0.999 2)回收率為87.3%。
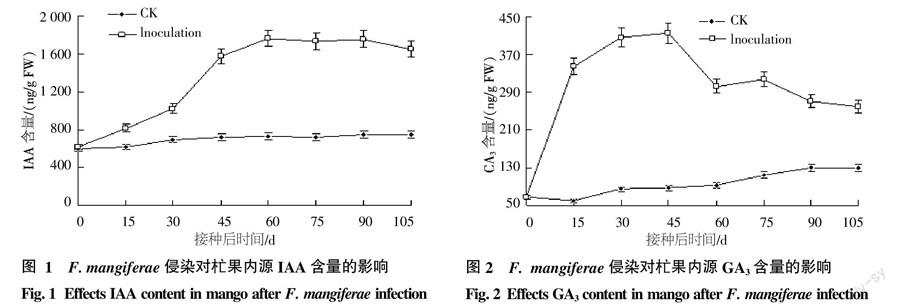
2.2 生長(zhǎng)素(IAA)含量變化規(guī)律測(cè)定
杧果健康對(duì)照組與F. mangiferae接種處理組頂芽?jī)?nèi)IAA含量的變化趨勢(shì)相同(圖1),均表現(xiàn)出逐漸上升的趨勢(shì),但F. mangiferae接種組頂芽?jī)?nèi)生長(zhǎng)素含量上升趨勢(shì)明顯高于對(duì)照組,且這種差異隨著時(shí)間的延長(zhǎng)逐漸加大。處理組頂芽?jī)?nèi)的生長(zhǎng)素在接種病原菌15 d后即顯著高于健康對(duì)照組,接種病原菌60~90 d內(nèi)含量達(dá)到最高,約為1 700 ng/g FW,是對(duì)照組(約為700 ng/g FW)的2.5倍左右,接種病原菌105 d時(shí),內(nèi)源生長(zhǎng)素含量雖略下降,但是仍顯著高于健康對(duì)照組。
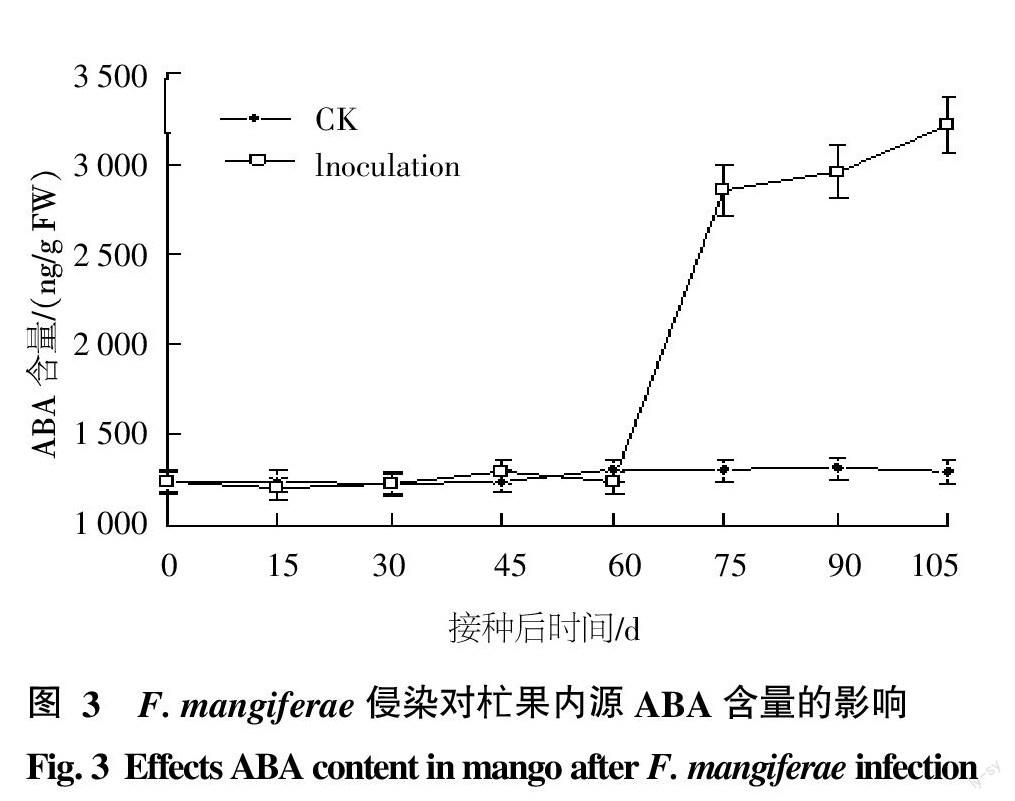
2.3 赤霉素(GA3)含量變化規(guī)律測(cè)定
從圖2中可以看出,杧果受F. mangiferae感染后,其頂芽?jī)?nèi)源赤霉素GA3含量呈現(xiàn)出先上升后下降的趨勢(shì),接種病原菌45 d達(dá)到最大值,約為415.65 ng/g FW,是空白對(duì)照組(86.4 ng/g FW)的4.8倍,45 d后其含量略下降,但仍顯著高于健康對(duì)照組。
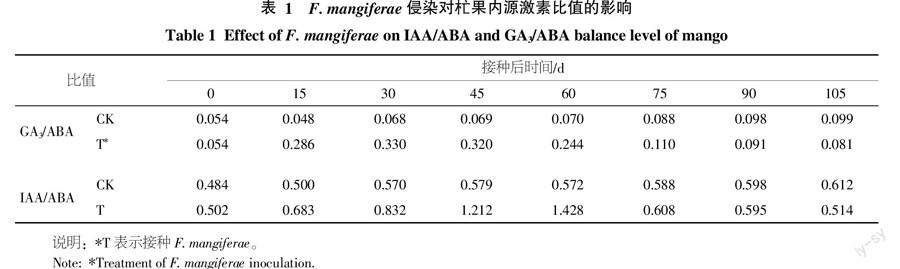
2.4 脫落酸(ABA)含量變化規(guī)律測(cè)定
杧果頂芽?jī)?nèi)源脫落酸的含量在F. mangiferae侵染后變化趨勢(shì)與生長(zhǎng)素和赤霉素有所不同,接種病原菌60 d內(nèi)無(wú)顯著變化,隨之迅速提高,接種病原菌105 d達(dá)到最大,為3 214.4 ng/g FW,是接種初期和對(duì)照組的2.6倍左右(圖3)。
2.5 病原菌侵染后杧果內(nèi)源激素比值的變化
植物激素間的生理效應(yīng)既相互促進(jìn)又相互拮抗,任何一類植物激素可影響到生長(zhǎng)發(fā)育的多個(gè)過(guò)程,同樣,植物生長(zhǎng)發(fā)育是多種激素相互作用的結(jié)果。IAA和GA3能促進(jìn)植物生長(zhǎng),延緩衰老,而ABA則促進(jìn)植物衰老,其比值通常用來(lái)表示植物內(nèi)源激素對(duì)生長(zhǎng)的作用是促進(jìn)還是抑制。從表1可以看出,在整個(gè)試驗(yàn)階段,健康杧果頂芽?jī)?nèi)IAA/ABA和GA3/ABA基本呈現(xiàn)上升趨勢(shì),而處理組IAA/ABA和GA3/ABA出現(xiàn)先上升后下降的趨勢(shì),且前期上升幅度遠(yuǎn)大于健康對(duì)照組,后期又低于對(duì)照處理。由此推斷,F(xiàn). mangiferae侵染能夠改變杧果頂芽?jī)?nèi)IAA/ABA和GA3/ABA的平衡,致使杧果的生長(zhǎng)發(fā)育受到影響,進(jìn)而引發(fā)畸形病的產(chǎn)生。
3 討論與結(jié)論
脫落酸、赤霉素和生長(zhǎng)素是調(diào)節(jié)植物細(xì)胞生長(zhǎng)代謝的重要內(nèi)源激素,在植物生長(zhǎng)發(fā)育中充當(dāng)著重要而又復(fù)雜的角色,其含量及變化決定著細(xì)胞與組織的生長(zhǎng)狀況。在植物體正常生長(zhǎng)情況下,內(nèi)源激素處于某種動(dòng)態(tài)平衡狀態(tài),調(diào)節(jié)植物的生長(zhǎng)發(fā)育。當(dāng)病原物入侵植物體后,內(nèi)源激素原有的動(dòng)態(tài)平衡被打破,從而使植株生長(zhǎng)發(fā)育受到影響,進(jìn)而產(chǎn)生病害。高增貴等[12]采用酶聯(lián)免疫吸附測(cè)定法,測(cè)定玉米苗期接種絲黑穗病菌后植株體內(nèi)生長(zhǎng)素、赤霉素、玉米素核苷和脫落酸4種內(nèi)源激素的含量變化,從寄主與病菌互作過(guò)程中植物內(nèi)源激素的變化研究玉米對(duì)絲黑穗病的抗性機(jī)制。研究結(jié)果表明,玉米絲黑穗病菌侵染玉米植株后,植株體內(nèi)單一激素ABA含量提高;IAA、GA3和ZR的含量降低;IAA/ABA、ZR/ABA、GA3/ABA及(IAA+GA3+ZR)/ABA的比值均明顯低于對(duì)照。趙錦等[13]用高效液相色譜法分別對(duì)健株、感病棗樹(shù)內(nèi)源激素的含量進(jìn)行了測(cè)定,結(jié)果表明植原體侵染棗樹(shù)植株后致使其內(nèi)源激素失衡,主要是細(xì)胞分裂素含量的增加,最終導(dǎo)致了棗瘋病癥狀表現(xiàn)。
將病原菌回接到杧果嫩梢上結(jié)果顯示,45 d之后接種頂芽處形成大量凸起小芽包,75 d后凸起的芽包大量萌發(fā),后期150 d萌發(fā)大量新芽,約是空白對(duì)照的7~9倍,且新葉細(xì)小生長(zhǎng)緩慢,與對(duì)照相比節(jié)間偏短,與典型的枝葉畸形相同。本研究結(jié)果表明,F(xiàn).mangiferae侵染后杧果頂芽?jī)?nèi)IAA、GA3和ABA的含量均有不同程度的提高,但提高幅度和病害的發(fā)展程度存在一定的相關(guān)性。病害發(fā)生初期,IAA和GA3含量迅速提高,刺激了杧果頂芽的迅速生長(zhǎng),形成大量新芽,然后由于頂芽的過(guò)度生長(zhǎng),植株?duì)I養(yǎng)狀況的供給不足,因此形成的芽簇生葉片變細(xì)變脆。病害發(fā)生后期,ABA的含量迅速上升,頂芽長(zhǎng)勢(shì)病變,是組織衰老死亡的征兆。從內(nèi)源激素比例中也可以看出,在供試試驗(yàn)階段,健康對(duì)照組內(nèi)IAA/ABA、GA3/ABA的比例呈逐漸上升的趨勢(shì),但是總體上升幅度不大。而接種處理組中,接種病原菌45 d時(shí),GA3/ABA的比值高達(dá)0.320,約是健康處理組的5倍左右,接種病原菌60 d時(shí),IAA/ABA的比值為1.428,健康處理組僅為0.572,GA3/ABA和IAA/ABA比值的增高促進(jìn)了杧果頂芽的徒長(zhǎng)。當(dāng)接種病原菌105 d,接種病原菌的杧果頂芽?jī)?nèi)GA3/ABA和IAA/ABA比值均比健康處理組低,顯示了杧果組織開(kāi)始處于衰老和死亡的狀態(tài),然而杧果頂芽?jī)?nèi)的內(nèi)源激素這種變化究竟是由病原菌侵染直接造成的,還是病原菌誘導(dǎo)了細(xì)胞損傷或影響了杧果頂芽同化作用后所造成的,還有待深入研究。
參考文獻(xiàn)
[1] 徐榮旗, 石磊巖. 棉花黃萎病菌致害棉株葉片內(nèi)源激素的動(dòng)態(tài)變化[J]. 棉花學(xué)報(bào), 2000, 12(6):310-312.
[2] Clarke S F, Burritt D J, Jameson P E, et al. Effects of plant hormones on white clover mosaic potexvirus double-stranded RNA[J]. Plant Pathology, 2000, 49(4): 428-434.
[3] Zhu S, Gao F, Cao X, et al. The rice dwarf virus P2 protein interacts with ent-kaurene oxidases in vivo, leading to reduced biosynthesis of gibberellins and rice dwarf symptoms[J]. Plant Physiology, 2005, 139(4): 1 935-1 945.
[4] Padmanabhan M S, Goregaoker S P, Golem S, et al. Interaction of the tobacco mosaic virus replicase protein with the Aux/IAA protein PAP1/IAA26 is associated with disease development[J]. Journal of Virology, 2005, 79(4): 2 549-2 558.
[5] Robert-Seilaniantz A, Navarro L, Bari R, et al. Pathological hormone imbalances[J]. Current Opinion in Plant Biology, 2007, 10(4): 372-379.
[6] 丁新倫, 張孟倩, 謝荔巖, 等. 實(shí)時(shí)熒光定量PCR檢測(cè)RSV脅迫下抗病、 感病水稻中與脫落酸相關(guān)基因的差異表達(dá)[J]. 激光生物學(xué)報(bào), 2008, 17(4): 464-469.
[7] 楊金廣, 王文婷, 丁新倫, 等. 水稻條紋病毒與水稻互作中的生長(zhǎng)素調(diào)控[J]. 農(nóng)業(yè)生物技術(shù)學(xué)報(bào), 2008, 16(4): 628-634.
[8] 柳 鳳, 盧乃會(huì), 詹儒林,等. 芒果畸形病研究進(jìn)展[J]. 熱帶作物學(xué)報(bào), 2012, 33(11): 2 104-2 109.
[9] Zhan R L, Yang S J, Liu F, et al. First report of Fusarium mangiferae in China causing mango malformation[J]. Plant Disease, 2012, 96(5): 762.
[10] 楊順錦, 詹儒林, 柳 鳳, 等. 金沙江干熱河谷地區(qū)芒果畸形病的病原菌[J]. 菌物學(xué)報(bào), 2011, 30(1): 27-31.
[11] 楊順錦, 詹儒林, 趙艷龍, 等. 果畸形病莖部解剖結(jié)構(gòu)及內(nèi)源激素含量變化[J]. 果樹(shù)學(xué)報(bào), 2011, 28(4): 708-711.
[12] 高增貴, 張寶艷, 張小飛, 等. 玉米絲黑穗病菌對(duì)植株體內(nèi)內(nèi)源激素水平改變的影響[J]. 玉米科學(xué), 2011, 19(2): 80-83.
[13] 趙 錦, 劉孟軍, 代 麗, 等. 棗瘋病病樹(shù)中內(nèi)源激素的變化研究[J]. 中國(guó)農(nóng)業(yè)科學(xué), 2006, 39(11): 2 255-2 260.
