新疆野杏種質資源表型性狀多樣性研究
2015-07-05 07:03:08趙世榮
西北植物學報 2015年5期
劉 娟,廖 康,劉 歡,曹 倩,孫 琪,趙世榮
(新疆農業大學 特色果樹研究中心,烏魯木齊830052)
新疆野杏(Armeniaca vulgaris Lam.)屬薔薇科(Rosaceae)杏屬植物(Armeniaca Mill.)[1],是一種珍貴的野生果樹資源,被認為是世界栽培杏的原生起源種群,對世界栽培杏的馴化起過決定性的作用[2]。伊犁地區是中國野杏的重要集中地,其分布范圍與數量僅次于野蘋果[3],1984年中國將伊犁野杏列入瀕危植物保護名錄[4],對這一種質的遺傳多樣性進行研究可為杏的新品種選育奠定基礎。表型多樣性研究已成為種質資源研究的重要內容,也是物種遺傳多樣性研究的重要組成部分[5],是遺傳多樣性與環境多樣性的綜合體現,其觀測方法簡單、直觀,所以大多數植物種質的鑒定目前仍是根據植物表型特征來進行區分、分類和命名[6]。已有通過系統的表型多樣性方法對 蘋果[7]、梅[8]、毛櫻桃[9]、野生秋子梨[10]、野生櫻桃李[5]、野生巴旦杏[6]、野生蘋果[11]、板栗[12]和山楂[13]等樹種進行了研究,并從一定程度上揭示了各樹種遺傳變異的規律。但對新疆野杏表型多樣性的研究較少,僅在何天明的研究中略有報道[2],本研究以分布于新疆伊犁地區霍城縣大西溝、新源縣博爾賽和鞏留縣伊力格代3個主要分布區的野杏種質資源為試材,對葉片、果實、果核和果仁等組織的形態特征進行觀測,分析不同居群及群體形態性狀的多樣性和變異特點,為野杏種質資源保護與利用提供依據。
1 材料和方法
1.1 試驗材料
2012~2013年,以新疆伊犁地區霍城縣、新源縣和鞏留縣的山區自然分布的野杏實生株系為試料,共計3個居群,135個單株樣本,分別進行GPS定位,各取樣地內單株間距離為100 m 以上,以避免采集到親緣關系較近的植株,各居群詳細取樣情況見表1。
1.2 觀測方法
以《杏種質資源描述規范和數據標準》[14]為參考依據,從葉片、果實、果核、種仁方面詳細調查形態特征,各指標重復觀測10次,具體觀測項目指標及方法見表2。
1.3 數據處理
依據《杏種質資源描述規范和數據標準》[14]將非數值型性狀數值化,對非數值型性狀進行賦值(表2),利用DPS 7.05數據處理軟件計算Simpson多樣性指數和Shannon-Weaver多樣性指數;利用Excel 2007和DPS 7.05數據處理軟件,分別計算數值性狀的平均值、標準差、變異系數、最大值、最小值、極值、Simpson和Shannon-Weaver多樣性指數;利用DPS 7.05軟件將各數值進行標準化處理,進行主成分分析,并計算歐氏遺傳距離,再利用MEGA

表1 新疆野杏種質資源取樣信息Table 1 Information of wild apricot resources in Xinjiang

表2 新疆野杏種質資源形態性狀觀測項目及記載標準Table 2 Observation project and recorded standard of wild apricot resources morphological characters in Xinjiang
2 結果與分析
2.1 居群間表型多樣性分析
22個非數值型性狀的Simpson指數(表3),除了葉背絨毛、葉基形態、對稱性和仁飽滿4個性狀,其它18個性狀均為霍城居群最大,鞏留居群居中,新源居群最小;葉背絨毛的Simpson指數為新源居群最大,鞏留居群居中,霍城居群最小;葉基形態和對稱性的Simpson指數為鞏留居群最大,霍城居群居中,新源居群最小;3個居群果面茸毛的Simpson指數相等;仁飽滿的Simpson 指數為新源居群最大,霍城居群居中,鞏留居群最小。對于Shannon-Weaver指數來說,22個非數值型性狀的Shannon-Weaver指數均為霍城居群最大,鞏留居群居中,新源居群最小。從平均值來看,無論是Simpson指數還是Shannon-Weaver指數,均表現為霍城居群最高(0.979 9和4.729 9),鞏留居群居中(0.975 5和4.385 7),新源居群最小(0.944 7和3.241 9)。結果表明,野杏非數值型性狀的表型多樣性表現為霍城居群最豐富,鞏留居群次之,新源居群相對最差。
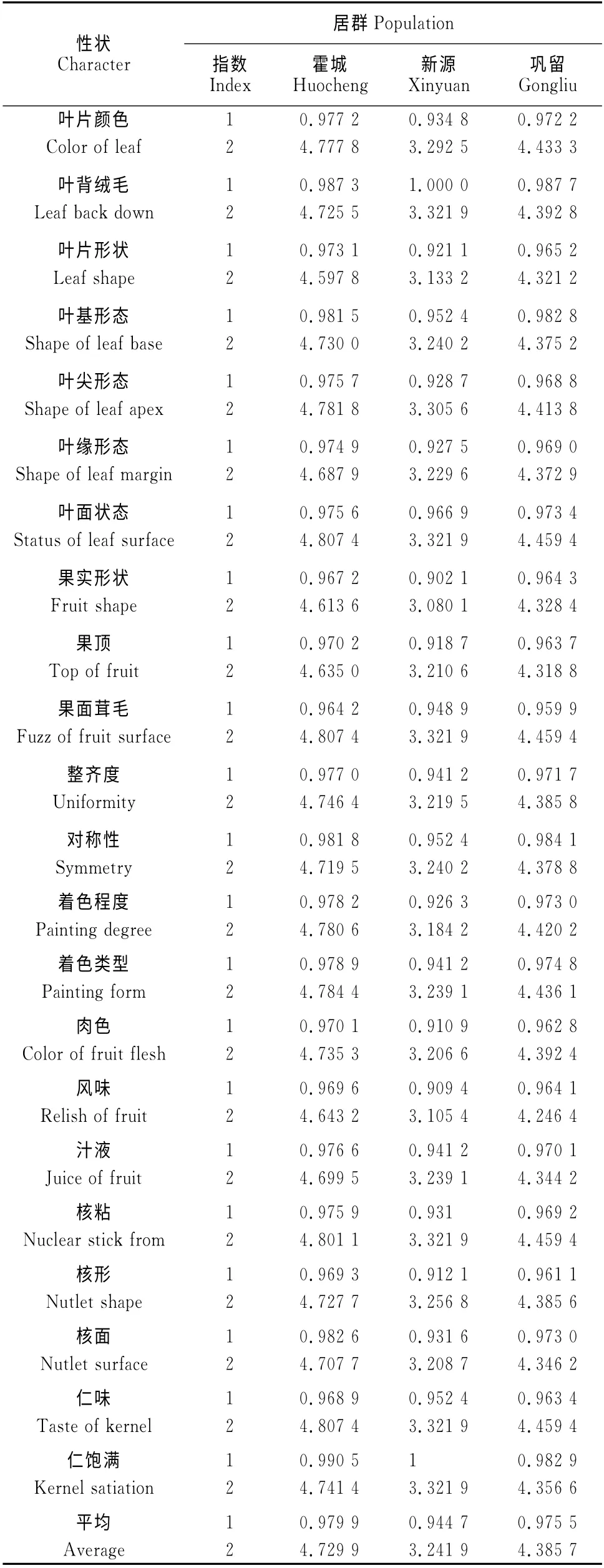
表3 新疆野杏非數值型性狀的多樣性指數分析Table 3 Diversity index analysis of non numeric characters of wild apricot in Xinjiang
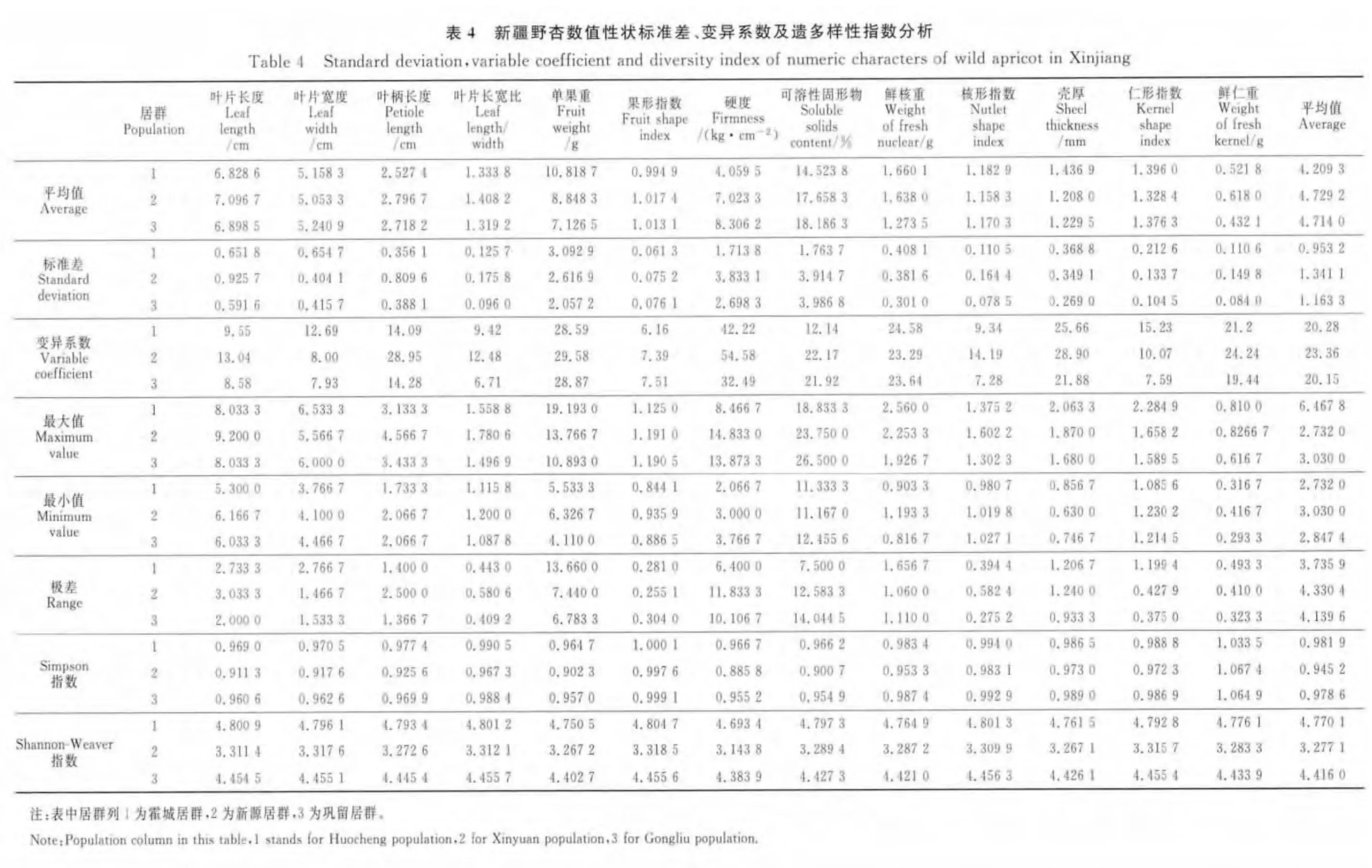
對于13個數值型性狀的Simpson指數來說(表4),葉片長度(0.969 0)、葉片寬度(0.970 5)、葉柄長度(0.977 4)、葉 片 長 寬 比(0.990 5)、單 果 重(0.964 7)、果形指數(1.000 1)、硬度(0.966 7)、可溶性固形物(0.966 2)、核形指數(0.994 0)和仁形指數(0.988 8)10 個性狀均為霍城居群最高,鞏留居群居中,新源居群最低;鮮核重(0.987 4)和殼厚(0.989 0)兩個性狀表現為鞏留居群最高,霍城居群居中,新源居群最低;只有鮮仁重(1.067 4)表現為新源居群最高,鞏留居群居中,霍城居群最低。13個數值型性狀的Shannon-Weaver指數(表4)均表現為霍城居群最高,鞏留居群居中,新源居群最低。從平均值來看,均表現為霍城居群最高(0.981 9,4.770 1),其次為鞏留居群(0.978 6,4.416 0),新源居群最低(0.945 2,3.277 1)。結果表明,霍城居群的數值型性狀的表型多樣性最豐富,鞏留居群次之,新源居群的數值型性狀的表型多樣性最差,這與非數值型性狀的結果一致。
2.2 居群間的變異與遺傳穩定性
由表4可知,各數值型性狀在不同居群內的變異系數差異均較大,變異幅度分別為:霍城居群在6.16% ~42.22% 之 間,新 源 居 群 在7.39% ~54.58%之間,鞏留居群在6.71%~32.49%之間;3個居群變異系數的平均值表現為新源居群最大(23.36%),霍城居群居中(20.28%),鞏留居群最小(20.15%);各居群不同性狀的變異系數在6.16%~54.58%之間,每個居群只有個別性狀的變異系數低于10%,其余都在10%以上。說明各性狀在居群內變異分化較明顯,且新源居群的變異分化要大于霍城居群和鞏留居群。
3個居群內不同性狀間的平均值差異也較大,結合標準差和變異系數來看,霍城居群和新源居群均為果形指數的遺傳性較穩定,鞏留居群內葉片長寬比的遺傳性較穩定;3個居群均以硬度的變異系數最大,另有單果重、鮮核重和殼厚的變異系數也普遍較大,遺傳穩定性相對較差。
?
2.3 居群間的遺傳關系
3個居群兩兩之間的歐氏距離在19.218 6~20.445 3之間(表5),其平均值為19.792 6,說明居群間的表型親緣關系相對較遠。其中,新源居群和鞏留居群之間的歐氏距離最遠,為20.445 3;霍城居群和新源居群間的歐氏距離最小,為19.218 6。由此可以得出,霍城居群和新源居群的親緣關系最近,遺傳分化程度小;新源居群和鞏留居群的親緣關系最遠,遺傳分化程度大。
根據表5的歐氏距離,利用MEGA 3.1軟件,采用UPGMA 聚類法對野杏資源3個天然居群進行聚類分析(圖1)。以距離系數10為標準,3個居群可聚為兩類:霍城居群和新源居群聚為第1類,鞏留居群為第2類;當距離系數為9.5時,3個居群各自聚為一類。從聚類圖看出,霍城居群和新源居群親緣關系最近。
2.4 天然群體的主成分分析
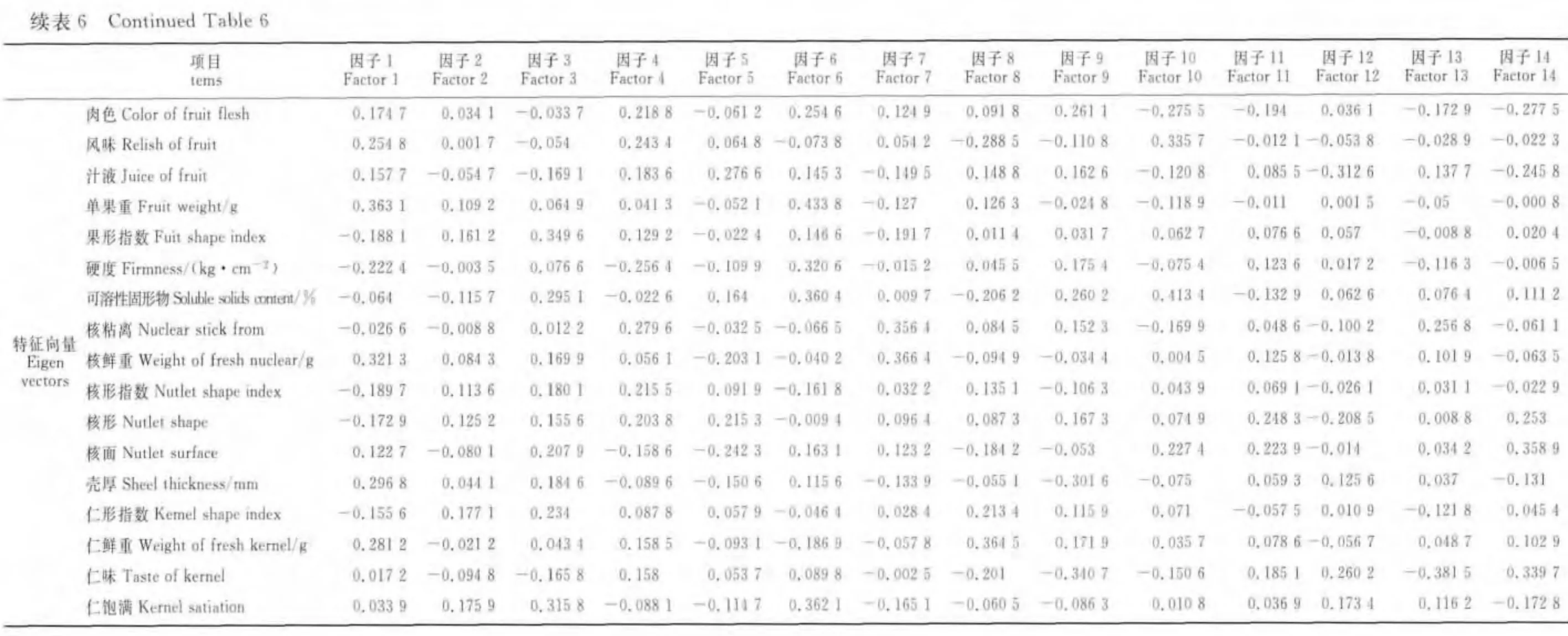
在35個主成分因子中前14個特征值的累計貢獻率為80.64%(表6)。其中第1主成分占15.37%,有較高載荷值的性狀指標依次是單果重、鮮核重、殼厚、鮮仁重、風味和硬度;第2主成分占8.93%,有較高載荷值的性狀指標依次是葉片長寬比、葉基形態、葉片形狀、葉片長度和葉片寬度,稱之為葉片性狀因子;第3主成分占8.41%,有較高載荷值的性狀指標依次是果形指數、葉片長度、仁飽滿度、可溶性固形物和果頂;第4主成分占6.85%,有較高載荷值的性狀指標依次是果形、著色類型、核粘離、硬度和風味;第5主成分占6.00%,有較高載荷值的性狀指標依次是葉片寬度、葉片長度、汁液、著色類型和葉柄長度;第6主成分占5.43%,有較高載荷值的性狀指標依次是單果重、仁飽滿度、可溶性固形物、硬度和肉色;第7主成分占5.02%,有較高載荷值的性狀指標依次是鮮核重、核粘離、果形、葉緣形態和葉尖形態;第8主成分占4.59%,有較高載荷值的性狀指標依次是葉片顏色、鮮仁重、葉背絨毛、葉尖形態和風味;第9主成分占4.08%,有較高載荷值的性狀指標依次是仁味、殼厚、葉尖形態、肉色和可溶性固形物;第10主成分占3.93%,有較高載荷值的性狀指標依次是可溶性固形物、葉片顏色、風味、葉背絨毛和肉色;第11主成分占3.43%,有較高載荷值的性狀指標依次是果面茸毛、葉背絨毛、果頂、核形和核面;第12主成分占3.24%,有較高載荷值的性狀指標依次是葉背絨毛、果面茸毛、汁液、仁味和著色類型;第13主成分占2.73%,有較高載荷值的性狀指標依次是葉緣形態、仁味、葉片顏色、著色類型和核粘離;第14主成分占2.65%,有較高載荷值的性狀指標依次是果面茸毛、核面、仁味、肉色和核形(表6)。根據特征根、貢獻率和累計貢獻率的基本情況表明,各性狀主成分累計貢獻率較分散,累計增加較緩慢。

表5 新疆野杏3個居群間基于35個表型性狀的歐氏距離Table 5 Euclidean distance of 3populations of wild apricot resources in Xinjiang for 35phenotypic traits
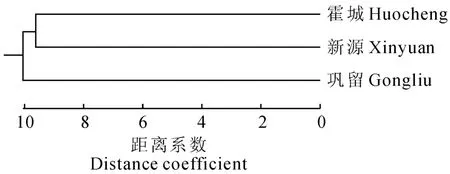
圖1 新疆野杏3個居群的UPGMA 聚類圖Fig.1 Dendrogram of UPGMA method cluster analysis of wild apricot in Xinjiang
3 討 論
3.1 居群的表型多樣性及親緣關系
Kostina等將栽培杏的700個品種劃分為4個地理生態群[15],各生態群的杏品種之間在樹體和果實等方面由于長期的生態適應發生了明顯的性狀差異[16],新疆野杏作為世界杏的起源種,其遺傳多樣性極為豐富。從研究結果可以看出,新疆野杏種質資源的不同性狀在不同的居群之間表現出了不同程度的多樣性。非數值性狀的Simpson指數和Shannon-Weaver指數表現為霍城居群最高,鞏留居群居中,新源居群最小;數值性狀的Simpson 指數和Shannon-Weaver指數均表現為霍城居群最高,其次為鞏留居群,新源居群最低。說明霍城居群的表型多樣性最豐富,鞏留居群居中,新源居群的表型多樣性最差,所以在進行資源保護時要優先保護霍城居群。
歐氏距離反映居群間的遺傳分化程度和親緣關系的遠近,是衡量居群間物種種源變異水平的重要指標。3個居群間表型性狀的歐氏距離在19.218 6~20.445 3,新源居群和鞏留居群的親緣關系最遠,遺傳分化程度大,二者間的歐氏距離為20.445 3;霍城居群和新源居群的親緣關系最近,遺傳分化程度小,二者間的歐氏距離為19.218 6。

?
?
對于野杏3個居群的遺傳多樣性和親緣關系所表現的這種關系,可能與樣本量的大小也有關系,樣本量大所涵蓋的表型表現較多,從而多樣性就豐富,反之,如果樣本量小,有可能所搜集的表型表現會不夠全面,從而導致多樣性相對較差,這就需要在今后的研究中對其進一步進行研究驗證。同時,由于表型性狀受環境條件及主觀因素影響很大,所以通過形態來分析果樹表型多樣性具有一定的局限性,最好結合同工酶標記、分子標記等不受環境及主觀因素影響的方法對表型多樣性進行鑒定。
3.2 居群的遺傳及變異
表型等性狀既具有變異性又具有穩定性,受其本身的遺傳組成和生態環境兩方面的影響,是生物適應其生態環境的表現形式[17]。表型變異必然蘊藏著遺傳變異,表型變異越大,可能存在的遺傳變異越大[18]。根據標準差和變異系數的數值大小可以判斷各個居群各數值型表型性狀的變異幅度[19]。若居群間表型性狀的平均值相差較多,單看標準差并不能準確地說明居群某一表型性狀的變幅大小,為了更客觀準確地說明各性狀的變異程度,還要結合變異系數來分析,變異系數是品種間表型性狀變異程度的反映,越大說明該表型性狀在品種間的差異越大,遺傳多樣性越豐富,越有可能利用該性狀來鑒別品種,變異系數小說明遺傳穩定性較好,不易受到環境等因素的影響[20-21]。本研究中13個數值型性狀的標準差、變異系數和極差的研究結果可以看出:同一性狀在居群間均有較大的差異,且同一居群內的各個性狀也有較大差異,變異系數的變異幅度在6.16%~54.58%之間,只有個別性狀的變異系數低于10%,其余都在10%以上,各性狀在居群間變異分化較明顯。霍城居群和新源居群均為果形指數的遺傳性較穩定,鞏留居群內葉片長寬比的遺傳性較穩定;3個居群均以硬度的變異系數最大,另有單果重、鮮核重和殼厚的變異系數也普遍較大,遺傳穩定性相對較差,與前人研究基本一致。馮濤等[22]認為新疆野蘋果不同居群均以果實縱徑、橫徑及果形指數等形態性狀的變異相對穩定;周龍等[5]也發現野生櫻桃李各天然群體間各個性狀中果形指數的變異系數最小,最為穩定。
3.3 表型性狀的主成分分析
主成分分析是指將原來較多的指標簡化為少數幾個新的綜合指標的多元統計方法[23],其主要作用是將多個變量通過線性組合構成為數不多的新變量,而且每個新變量盡可能包含原始變量的信息。本研究在構成主成分的信息中篩選出了14個性狀主分量,包含的總遺傳信息為80.64%,表明葉片長度、葉片寬度、葉片長寬比、葉基形態、葉尖形態、著色類型、單果重、鮮核重、鮮仁重、果實形狀、可溶性固形物等性狀是造成野杏種質表型差異的主要因素。以往的一些研究,周龍等[5]從14個主成分因子中提出前6個特征值,其累計貢獻率為88.76%,說明主成分因子足以代表原始因子所代表的大部分信息,而本研究中根據各自特征根、貢獻率和累計貢獻率的基本情況表明,提取出的14個主成分的累計貢獻率較分散,累計增加較緩慢,主成分分析得出的主分量反應總體遺傳信息的能力相對較弱,在材料選擇時不能把主分量作為唯一的評價標準。
[1] 俞德俊.落葉果樹分類學[M].上海:上海科學技術出版社,1984:67.
[2] 何天明.中國普通杏(Prunus armeniaca)種質資源遺傳多樣性及紫杏(P.dasycarpa)起源的研究[D].山東泰安:山東農業大學,2006:6.
[3] 廖 康.新疆野生果樹資源研究[M].烏魯木齊:新疆人民出版總社,2013:133.
[4] LIU W,CHEN X S,LIANG Q,et al.Interspecific hybridization of Prunus persica with P.armeniacaand P.salicinausing embryo rescue[J].Plant Cell Tissue and Organ Culture,2007,88(3):289-299.
[5] ZHOU L(周 龍),HU J F(胡建芳),XU ZH(許 正),et al.Analysis on fruit phenotypic diversity of natural populations in Prunus divaricata[J].Journal of Jilin Agricultural University(吉林農業大學學報),2011,33(6):637-642(in Chinese).
[6] ZENG B(曾斌),LUO SH P(羅淑萍),LI J(李 疆),et al.Study on phenotypic diversity of morphologic characteristics of Amygdalus ledebouriana Schlecht.leafs in Xinjiang[J].Xinjiang Agricultural Sciences(新疆農業科學),2008,45(2):221-224(in Chinese).
[7] XING L M邢麗敏HUAI X T槐心體ZHANG X ZH張新忠et al.Genetic diversity in morphologystress tolerance and graftcompatibility among species,ecotypes and their segregation within populations of apple rootstock resources[J].Journal of Fruit Science(果樹學報),2013,30(4):516-525(in Chinese).
[8] SHEN Y Y(沈玉英),DING X J(丁夏君),GAO ZH H(高志紅),et al.Analysis of genetic diversity in Prunus mume based on mathematic classification of morphologic characters[J].Journal of Fruit Science(果樹學報),2011,28(5):802-807(in Chinese).
[9] YANG X(楊 鑫),YANG J Q(楊家全),CHEN H(陳 紅),et al.Genetic diversity analysis on the characteristics of fruit cores of wild peaches in Guizhou[J].Acta Bot.Boreal.-Occident.Sin.(西北植物學報),2013,33(11):2 225-2 231(in Chinese).
[10] AN M M(安萌萌),WANG Y T(王艷廷),SONG Y(宋 楊),et al.Genetic diversity of fruit phenotypic traits of wild Pyrus ussuriensis Maxim[J].Scientia Agricultura Sinica(中國農業科學),2014,47(15):3 034-3 043(in Chinese).
[11] LIU J(劉 靜),ZHOU Q H(周慶和),SUN H W(孫海偉),et al.Study on the phenotype biodiversity of Xinjiang wild apples(Malus sieversii)[J].Journal of Fruit Science(果樹學報),2004,21(4):285-288(in Chinese).
[12] MA Y M(馬玉敏),CHEN X S(陳學森),HE T M(何天明),et al.Genetic diversity of morphological traits in wild populations of Castanea mollissima Blume[J].Acta Horticulturae Sinica(園藝學報),2008,35(12):1 717-1 726(in Chinese).
[13] JIANG Y L(姜英林),DONG W X(董文軒).Diversity analysis of morphological characters in hawthorn germplasm resources[J].Northern Fruits(北方果樹),2009,(1):8-10(in Chinese).
[14] 劉 寧,劉威生.杏種質資源描述規范和數據標準[M].北京:中國農業出版社,2006:12-24.
[15] KOSTINA K F.Application of phytogeographical method to apricot classification[J].Trud.Nikit Bot.Sad.,1964:37.
[16] 孫家正.南疆栽培杏部分表型性狀遺傳多樣性研究[D].山東泰安:山東農業大學,2010:6.
[17] JIANG X B(江錫兵),GONG B CH(龔榜初),LIU Q ZH(劉慶忠),et al.Phenotypic diversity of important agronomic traits of local cultivars of Chinese chestnut[J].Acta Horticulturae Sinica(園藝學報),2014,41(4):641-652(in Chinese).
[18] MENG CH(孟 超),ZHENG X(鄭 昕),JI ZH F(姬志峰),et al.Phenotypic diversity of natural population of Acer grosser in Shanxi[J].Acta Bot.Boreal.-Occident.Sin.(西北植物學報),2013,33(11):2 232-2 240(in Chinese).
[19] GUO CH Y(郭傳友),HUANG J Q(黃堅欽),WANG ZH(王 正),et al.Phenotypic diversity of fruit characters in Carya dabieshanensis[J].Nonwood Forest Research(經濟林研究),2007,25(3):15-18(in Chinese).
[20] THEIMER T C.Intraspecific variation in seed sise affects scatterhoarding behavior of an Australian tropical rain-forest rodent[J].Journal of Tropical Ecology,2003,19(1):95-98.
[21] WANG L R(王力榮),ZHU G R(朱更瑞),FANG W CH(方偉超).The evaluating criteria of some botanical quantitative characters of peach genetic resources[J].Scientia Agricultura Sinica(中國農業科學),2005,38(4):770-776(in Chinese).
[22] FENG T(馮 濤),ZHANG H(張 紅),CHEN X S(陳學森),et al.Genetic diversity of fruit morphological traits and content of mineral element in Malus sieversii(Ldb.)Roem.and its elite seedlings[J].Journal of Plant Genetic Resources(植 物 遺 傳 資 源 學 報),2006,7(3):270-276(in Chinese).
[23] 紀海波.西瓜種質資源表型多樣性研究[D].蘭州:甘肅農業大學,2013:6.
