尿路致病性大腸埃希菌FYUA基因敲除株的構建及其生物學特性
2015-08-11 08:44:11鄧小燕潘嘉韻吳曉蔓
基礎醫學與臨床 2015年4期
鄧 聰,彭 亮,鄧小燕,潘嘉韻,吳曉蔓
(廣州醫科大學 附屬第二醫院 檢驗科, 廣東 廣州 510260)
?
研究論文
尿路致病性大腸埃希菌FYUA基因敲除株的構建及其生物學特性
鄧 聰,彭 亮,鄧小燕,潘嘉韻,吳曉蔓*
(廣州醫科大學 附屬第二醫院 檢驗科, 廣東 廣州 510260)
目的構建尿路致病性大腸埃希菌FYUA基因敲除株,比較野生株和敲除株之間生物學特性的改變。方法利用λ Red同源重組系統構建FYUA基因敲除株△FYUA;測定A600繪制CFT073和△FYUA在LB液體培養基和無菌尿液中的增殖曲線;平板菌落計數法比較CFT073和△FYUA菌落形成能力;利用96孔板結晶紫染色法比較CFT073和△FYUA生物膜形成能力。結果成功構建CFT073 FYUA基因敲除株△FYUA;CFT073和△FYUA在LB液體培養基中具有相似的增殖曲線,△FYUA在無菌尿液中的增殖速率明顯低于CFT073(P<0.05);CFT073和△FYUA菌落形成能力無明顯區別;△FYUA的生物膜形成能力低于CFT073(P<0.05)。結論FYUA基因對于菌株在無菌尿液中的生長繁殖以及生物膜形成可能發揮重要作用。
FYUA基因;基因敲除;尿路感染;大腸埃希菌
尿路感染(urinary tract infection,UTIs)是目前臨床上最普遍的細菌感染之一,每年都有數百萬人患尿路感染,并且復發率很高[1- 2]。80%~90%的尿路感染由尿路致病性大腸埃希菌(uropathogenicEscherichiacoli, UPEC)所引起。
UPEC目前已知的毒力因子包括莢膜、Afa/Dr黏附素、Ⅰ型菌毛、α-溶血素、脂多糖和含鐵產氣桿菌素[3- 5]等。鐵是除乳酸桿菌和伯氏疏螺旋體外所有微生物的必要元素。但宿主體內的鐵含量很低并一般與轉鐵蛋白等螯合蛋白相結合。因此病原微生物需要適應宿主體內鐵限制的環境并且進化出一種高效的鐵轉運系統。絕大多數腸桿菌科細菌基因組中的強毒力島(high pathogenicity island,HPI)可編碼3價鐵螯合復合物以及受體。利用這些螯合物對鐵的高親和力,可以將結合于宿主鐵結合蛋白的鐵解離并將其通過受體運回細菌[6- 7]。FYUA是HPI中一個重要基因,其編碼產物FYUA定位于細菌外膜,是3價鐵螯合物的外膜受體。FYUA目前已經被發現與多種腸桿菌科細菌的毒力密切相關,且流行病學調查資料顯示其與尿路感染的發生有關[8- 10],但FYUA在UPEC中的致病機制目前尚不明確。
本研究通過構建UPEC的FYUA基因敲除株,比較其與野生株的生物學特性,初步探討FYUA基因在UPEC致病中的作用。
1 材料與方法
1.1 菌株和質粒
大腸埃希菌CFT073菌株(臨床尿路感染分離的大腸埃希菌,由美國南加州大學洛杉磯兒童醫院黃勝和教授惠贈);質粒pKD46,為人工構建的編碼Red重組系統GAM、BET和EXO基因的重組質粒,此質粒為溫度敏感型,在37 ℃及以上溫度時停止復制。pKD4質粒,為PCR提供卡那霉素抗性基因的模板,在該抗性基因表達框的兩側各帶有一個FRT位點,其間的序列可通過定向重組(pCP20質粒表達的酵母來源的FRT重組酶)刪除。pCP20質粒,具有氨芐青霉素抗性,可編碼FRT重組酶,用于刪除替換目標基因的卡那霉素抗性基因,該質粒也是溫度敏感質粒,通過提高培養溫度可使其從細胞中消失。
1.2 試劑
TaqPlus DNA聚合酶,質粒抽提試劑盒和膠回收試劑盒(天根生物有限公司),L-阿拉伯糖(Sigma公司)。氨芐青霉素、卡那霉素和LB培養基等試劑(上海生工生物技術公司)(氨芐青霉素和卡那霉素在培養基中的工作濃度分別為50、25 mg/L)。測序由上海生工生物技術公司完成。
1.3 引物設計與合成
用于擴增FYUA基因外側同源臂的引物序列:H1-P1:上游引物5′-ACCATTGTCGGTATTTTTGGCG TTTCGCCGTCTTACAGGGACTCACAACACATATGAA TATCCTCCTTAGTTCCTATTC-3′;H2-P2:下游引物5′-TTAACTAGTGGTGTAAAAGGGATACCTTTTCGGT ATCCCTTTTACAATAAGAGCTGCTTCGAAGTTCCT-3′。下劃線序列:FYUA基因兩側的同源重組手臂序列;無下劃線序列:卡那霉素抗性基因的特異引物序列。鑒定FYUA基因敲除是否成功的引物序列:H3:5′-GTTGCGTCAACAACTGACCCGTATG-3′;H4:5′-C AGGCTAGTATAGTTTGTGGTAGCTATGAG-3′。以上引物均有上海生工生物技術公司合成,引物所在位置見圖1。
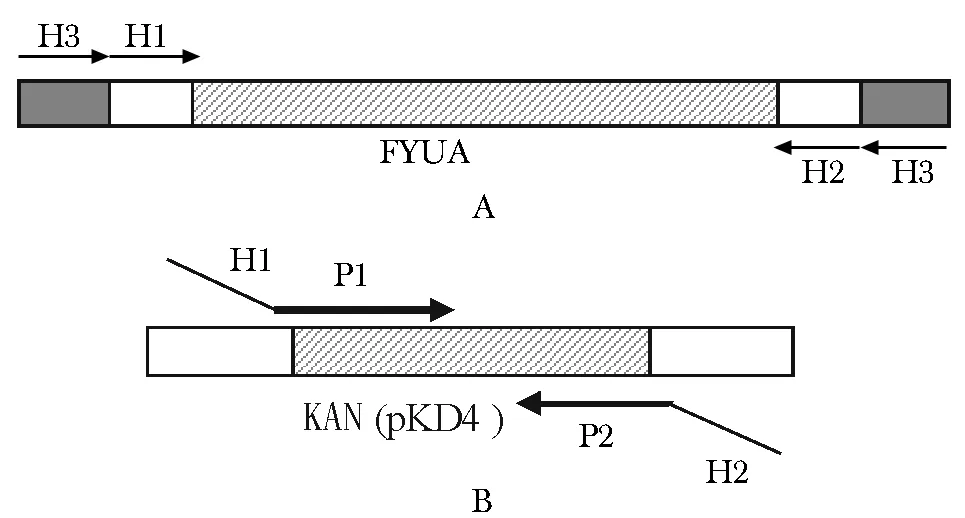
A.FYUA gene and its upstream and downstream primers location; B.KAN resistance gene in pKD4 plasmid and its upstream and downstream primers location圖1 各引物所在位置示意圖Fig 1 Positions of genome-specific primers
1.4 CFT073FYUA基因缺失株的構建
外源性打靶片段的擴增與純化:以pKD4質粒為模板,H1-P1和H2-P2作為引物,擴增含卡那霉素的靶基因。反應條件:95 ℃預變性5 min,95 ℃變性30 s, 65 ℃退火45 s,72 ℃延伸90 s,循環21次,每個循環退火溫度減0.5 ℃。95 ℃變性30 s,55 ℃退火45 s,72 ℃延伸90 s,循環15次。最后72 ℃延伸7 min。PCR產物經1.0%瓊脂糖凝膠電泳驗證條帶正確后,進行膠回收。
Red重組系統的誘導表達和外源線性打靶片段的電轉化:將pKD46質粒電轉化至感受態的CFT073菌株中,在含有氨芐青霉素的LB液體培養基中30 ℃振蕩培養過夜,次日按1∶100接種至50 mL LB液體培養基(不含抗生素),同時加入100 μL 1 mol/L L-阿拉伯糖,30 ℃,220 r/min培養至A600達到0.5。冰上預冷15 min,4 ℃, 4000 r/min離心15 min,棄培養基,用冰冷滅菌去離子水洗滌菌體,重復3次后,濃縮100倍成感受態細胞。 取5 μL純化的打靶片段和50 μL感受態細胞至預冷的電擊杯(0.2 cm)中,用Bio-rad電轉儀進行電轉化(電轉參數:電阻200 Ω,電容25 μF,電壓2.5 kV, 電擊時間4.6 ms)。電擊后迅速加入1 mL 37 ℃保溫的LB培養基。吹打混勻,轉移至新的無菌1.5 mL 離心管,37 ℃,200 r/min培養復蘇2 h。取500 μL轉化菌液涂布于含卡那霉素的LB平板上,37 ℃過夜培養至單菌落形成。用外側引物H3和H4進行PCR方法篩選陽性克隆, 如發生正確重組,則擴增長度為2 120 bp,未正確重組的克隆PCR長度仍為2 675 bp。陽性克隆命名為CFT073/ΔFYUA/KAN。
卡那霉素抗性基因的刪除:將pCP20質粒電轉化入CFT073/ΔFYUA/KAN感受態細胞,取500 μL轉化菌液涂布于含氨芐青霉素的LB平板上,30 ℃過夜培養至單菌落形成。用外側引物H3和H4進行PCR反應鑒定,如卡那霉素抗性基因被刪除,則擴增長度為727 bp,未發生正確刪除的克隆PCR長度仍為2 120 bp。篩選出FYUA基因和卡那霉素抗性基因均被刪除的陽性克隆。將該陽性克隆劃線接種LB平板(無抗性),42 ℃培養過夜(高溫培養促使pCP20質粒丟失)。
pCP20質粒丟失的鑒定:隨機挑選1個克隆接種入20 μL LB培養液,取10 μL劃線接種LB(含50 mg/L氨芐青霉素)平板,37 ℃培養過夜。次日觀察,無克隆生長,即表示pCP20質粒已丟失。
FYUA敲除菌株的鑒定:用外側引物H3和H4進行PCR反應鑒定,選取片段長度相符的PCR產物,經瓊脂糖電泳膠回收后,送上海生工生物技術公司測序。測序驗證正確敲除的菌株命名為△FYUA。
1.5CFT073與ΔFYUA在LB培養液中增殖曲線的測定
分別從新鮮涂布的平板上挑取CFT073和ΔFYUA單菌落接種至5mLLB液體培養基中,37 ℃,180r/min振蕩培養過夜,次日再將菌液以1∶100的比例接種于LB液體培養基中,37 ℃,180r/min培養,每隔2h取樣測定A600,繪制增殖曲線。實驗重復3次。
1.6CFT073與ΔFYUA在無菌尿液中增殖曲線的測定
征集10名志愿者(志愿者過去無尿路感染史且2個月內未使用過抗生素),每次實驗隨機選取3~4名志愿者,留取新鮮尿液,混合后經無菌過濾,存放于冰箱2~3d內待用。挑取CFT073和△FYUA單菌落,分別接種至5mLLB液體培養基中,37 ℃,180r/min振蕩培養過夜,次日再將菌液以1∶100的比例接種于無菌尿液,37 ℃,180r/min搖床培養,每隔2h取樣測定A600,繪制增殖曲線。實驗重復3次。
1.7 菌落形成能力的比較
采用平板菌落計數法。凍存菌液接種于LB液體培養基,37 ℃,180r/min振蕩培養過夜,取100μL菌液,加入900μLLB培養基中,倍比10倍稀釋,1×10-6和1×10-7稀釋菌液涂布LB平板,靜置培養過夜后計算菌落數。實驗重復3次。
1.8 FYUA基因缺失對生物膜形成能力的影響
CFT073和△FYUA菌液接種于LB液體培養基中, 37 ℃,180r/min搖床培養過夜,次日取1mL菌液, 12 000r/min離心2min,用1mL無菌尿液重懸,每孔15μL接種于24孔板,再向每孔中加入1.5mL無菌尿液稀釋混勻。每種菌株接種5個復孔,同時設置3孔不加菌的無菌尿液作為空白對照。24孔板置于37 ℃靜止孵育12h。12h后,棄上清,無菌PBS緩沖液輕輕沖洗3次。風干后 0.1%結晶紫染色15min,無菌PBS清洗3次。80∶20的乙醇:丙酮溶液提取結晶紫。每孔各取200μL提取物至96孔板,酶標儀檢測570nm處的吸光度(A)值。每孔重復測量3次,取平均值。取5個復孔的平均值減去空白對照孔的A570平均值為其A值。實驗重復3次。
1.9 統計學分析
2 結果
2.1PCR擴增外源打靶線性片段
如圖2所示,擴增出兩端與FYUA基因上下游同源,中間為卡那霉素抗性基因的DNA線性片段,經電泳鑒定長度為1 567bp,與理論值一致(圖2)。
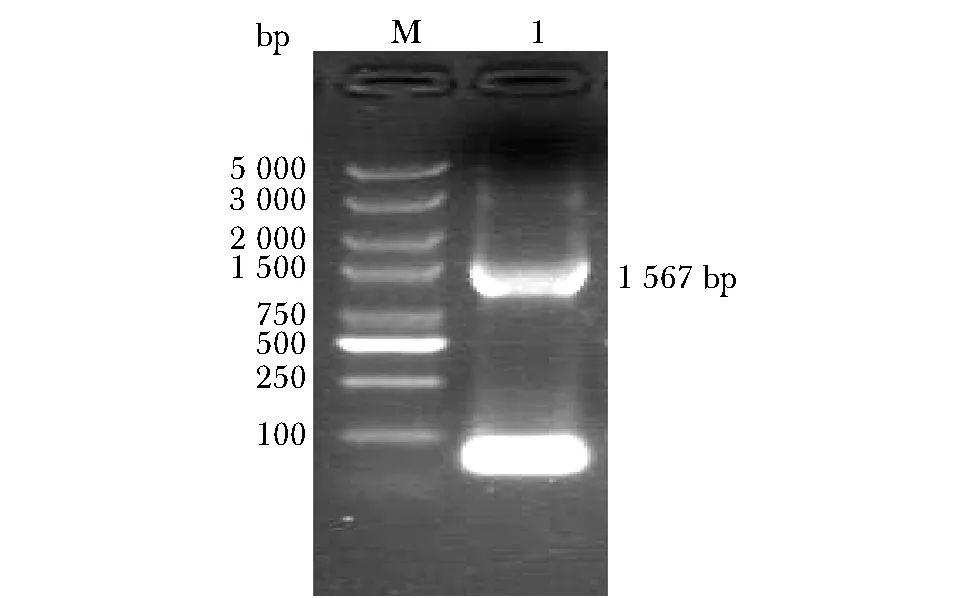
1.target linear fragment,1 567 bp; M.DNA Marker,圖2 外源打靶線性片段電泳圖Fig 2 Electrophoresis for exogenous target linear fragment
2.2 卡那抗性基因替代FYUA基因
如圖3所示,2號克隆為發生正確重組的克隆,命名為CFT073/ΔFYUA/KAN。
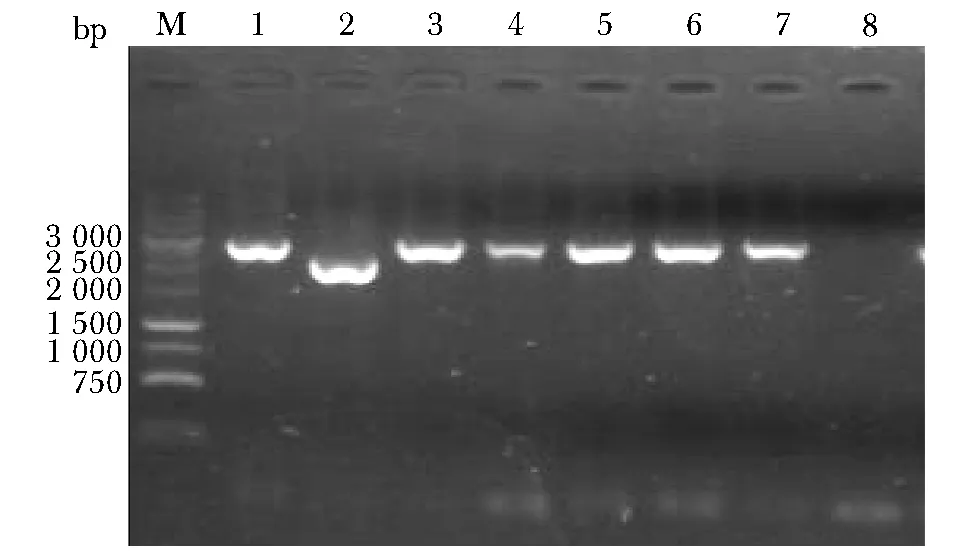
M.DNA Marker; 1~7.amplified results of clone No.1~7; 8.negative control圖3 CFT073/ΔFYUA/KAN的PCR鑒定Fig 3 Identification for CFT073/ΔFYUA/KAN by PCR
2.3 卡那霉素抗性基因的刪除
如圖4所示:第5號克隆為陽性克隆,該克隆的FYUA基因被敲除, 卡那霉素抗性基因也被刪除。

M.DNA Marker; 1~8.amplified results of clone No.1~8圖4 卡那霉素抗性基因刪除的PCR鑒定Fig 4 Identification for deletion of KAN resistant gene by PCR
2.4 測序驗證
結果證實局部序列與預期系列完全一致,測序驗證正確的菌株命名為ΔFYUA。
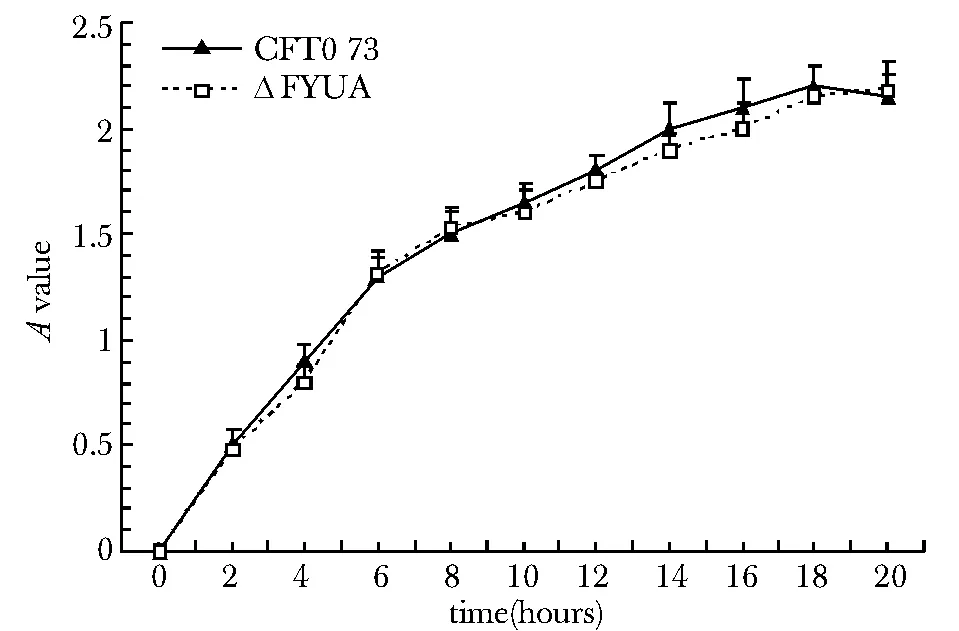
圖5 CFT073與ΔFYUA在LB液體培養基中的增殖曲線Fig 5 Proliferation curves of CFT073 strain and △FYUA strain in Luria-Bertani liquid medium
2.5 CFT073與ΔFYUA增殖曲線的測定
在LB液體培養基中,ΔFYUA和CFT073菌株具有相似的增殖曲線 (圖5)。而在無菌尿液中,ΔFYUA的增殖速率明顯低于CFT073菌株(P<0.05),并且無論是ΔFYUA還是CFT073菌株,其在無菌尿液中的增殖總數都遠低于LB液體培養基(圖6);12 h后,無菌尿液中生長的ΔFYUA和CFT073菌株都進入衰亡期,菌株數目迅速減低。而LB液體培養基中,一直到20 h,菌株還維持較高的增殖速率。
2.7 生物膜形成能力比較
CFT073 570 nm處的吸光度A值為0.43±0.04,顯著高于ΔFYUA的0.33±0.05(P<0.05)。
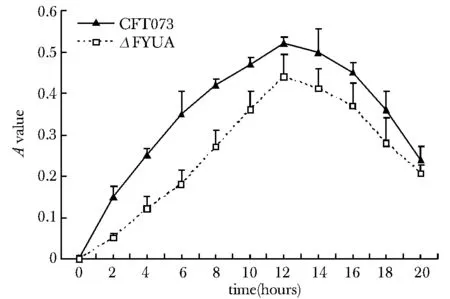
圖6 CFT073與ΔFYUA在無菌尿液中的增殖曲線Fig 6 Proliferation curves of CFT073 strain and △FYUA strain in sterile urine
2.6 落形成能力比較
ΔFYUA和CFT073菌株在平板培養14 h后菌落形成能力無顯著差異(表1)。

表1 CFT073和ΔFYUA菌落形成能力比較Table 1 Comparison on colony formation abilities between CFT073 strain and △FYUA strain
3 討論
基因敲除是研究某一具體基因功能最直接的方法。本研究使用的基因敲除的方法為λ Red同源重組。該方法只需要較短的同源序列,而不需要特定的限制性酶切位點即可完成重組過程,大大簡化了基因敲除的操作步驟,并且具有很高的重組陽性率。但傳統的λ Red同源重組發生之后,會在靶標分子上留下篩選標記基因,以本研究為例,會在目的基因位置留下卡那霉素的篩選標記基因。本研究通過引入pCP20質粒來實現卡那霉素抗性基因的刪除,同時通過提高培養溫度使pCP20質粒從細胞中消失。通過刪除篩選標記基因和去除外源性質粒,可以最大程度地減低基因操作對宿主菌的影響,從而更能保證實驗結果的客觀準確。
本研究通過構建FYUA基因敲除株,研究其在菌株增殖速率、菌落形成能力以及生物膜形成能力的改變。從結果可見,ΔFYUA和CFT073菌株在LB液體培養基中具有相似的增殖曲線,說明FYUA基因的缺失在營養豐富的LB培養基中對細菌的增殖速率未產生明顯影響。而在營養物質缺乏且鐵含量很低的無菌尿液培養基中,FYUA基因的缺失明顯降低了菌株的增殖速率。在尿液中快速生長繁殖是UPEC致病必需的重要一步,FYUA基因的缺失降低了菌株在尿液中的生長速率,提示FYUA基因可能通過促進細菌在尿液中的生長繁殖而在UPEC致病中發揮重要作用,其具體機制還需要進一步深入研究。
細菌生物膜是指細菌黏附于生物或非生物的表面時,形成的含有大量細菌的膜樣復合物。細菌感染形成生物膜后,可以發揮其協同作用,形成復雜有序的立體結構,從而可以逃避宿主免疫系統的攻擊,抵抗吞噬,具有較高程度的耐藥性等,并且可以在生物膜解離后播散導致其他系統的感染。本研究評估了FYUA基因缺失對菌株生物膜形成能力的影響。結果顯示,FYUA基因敲除株與野生株相比,其生物膜形成能力明顯減低,提示FYUA基因在生物膜形成方面具有一定作用。
尿路感染由于其復發率高以及UPEC對多種抗生素的耐藥性,目前仍是臨床治療的重點和難點問題。本研究初步探討了FYUA基因在UPEC致病中的作用,為UPEC致病機制的進一步明確奠定了基礎。
[1] Schmiemann G, Kniehl E, Gebhardt K,etal. The diagnosis of urinary tract infection: a systematic review[J]. Dtsch Arztebl Int,2010,107: 361- 367.
[2] Larcombe J. Urinary tract infection in children[J]. Am Fam Physician, 2010,82:1252- 1256.
[3] Agarwal J, Srivastava S, Singh M. Pathogenomics of uropathogenic Escherichia coli[J]. Indian J Med Microbiol,2012,30:141- 149.
[5] Hannan TJ, Totsika M,Mansfield KJ,etal. Host-pathogen checkpoints and population bottlenecks in persistent and intracellular uropathogenic Escherichia coli bladder infection[J]. FEMS Microbiol Rev,2012,36:616- 648.
[6] Hancock V, Ferrières L, Klemm P. The ferric yersiniabactin uptake receptor FYUA is required for efficient biofilm formation by urinary tract infectious Escherichia coli in human urine[J]. Microbiology,2008,154: 167- 175.
[7] Schubert S, Rakin A, Heesemann J. The Yersinia highpathogenicity island (HPI): evolutionary and functional aspects[J]. Int J Med Microbiol, 2004, 294: 83- 94.
[9] Rachel R, Spurbeck, Paul C,etal. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract[J]. InfectImmun, 2012, 80:4115- 4122.
[10] Snyder JA, Haugen BJ, Buckles EL,etal. Transcriptome of uropathogenic Escherichia coli during urinary tract infection[J]. Infect Immun. 2004,72:6373- 6381.
新聞點擊
肥胖增加偏頭痛概率
據美國WebMD醫學新聞網(2013-09-09)報道,肥胖者陣發性偏頭痛的風險較高,這是一種常見的偏頭痛,每個月發作少于14 d;慢性偏頭痛是每個月至少發作15 d。
美國神經學會指出,偏頭痛是指頭的某部位有節奏地強烈陣痛,癥狀包括惡心、嘔吐、以及對聲光敏感,有10%以上的人罹患此病。
這篇研究針對超過3 800位高BMI的成人,他們有陣發性偏頭痛的概率比低BMI的成人81%,尤其是女性及未滿50歲患者。
這種橫斷面的研究無法證明肥胖會造成陣發性偏頭痛,但醫生們應該建議患者注意飲食與運動。
但到底是先肥胖還是先有偏頭痛?Toledo大學的Gretchen Tietjen博士認為,這有很多種可能性,有可能事先有偏頭痛,開始吃amitriptyline或是valproic acid等藥物,而這些藥物會造成體質量增加。
肥胖與偏頭痛有關的理論:脂肪組織(adipose)中的炎性物質釋放,而年輕的婦女以及肥胖者有較多adipose組織,這或許可以解釋為何這些人比較常有偏頭痛。
減重是否代表偏頭痛的頻率會減少呢?專家們認為,雖然會鼓勵肥胖者減肥,但這不等于會舒緩偏頭痛。
這篇研究刊登在2013年9月11日的神經學(Neurology)期刊中。
魚翅恐傷神經影響生育
據英國《BBC新聞》(BBC NEWS)2013年9月19日報道,香港政府于2013年9月13日正式公告,今后魚翅不能在香港國宴上出現,而美國紐約也將在2014年7月1日,讓魚翅從餐桌徹底消失。
美國海洋藥物期刊指出,研究員發現魚翅含有144~1 838 μg/g β-N-甲氨基-L-丙氨酸(BMAA)神經毒素,而這與阿爾茨海默病患者、葛雷克氏病患者腦中的神經毒素相當。
全球頂級的公衛雜志EHP在2013年11月5日就刊登了,魚翅含有神經毒素BMAA,它與阿爾茨海默病有關,而且對孕婦和幼童傷害更大。
ConstructionofuropathogenicEscherichia colistrainwithFYUAgenedeletionanditsbiologicalproperties
DENG Cong, PENG Liang, DENG Xiao-yan,PAN Jia-yun, WU Xiao-man*
(Dept. of Clinical Laboratory, the Second Affiliated Hospital of Guangzhou Medical University, Guangzhou 510260,China)
Objective To construct aFYUAgene deletion mutant of uropathogenicEscherichiacolistrain CFT073;To compare the different biological properties between wide-type and mutant.Methods λ Red homologous recombination technology was used to construct the FYUA gene deletion strain △FYUA.TheproliferationcurveofCFT073and△FYUAinLuria-Bertaniliquidmediumandsterileurineweredrawedbymeasuringtheabsorbanceat600nm.ColonyformationabilitiesofCFT073and△FYUAwereevalutatdbycountingthenumberofcoloniesintheplates.CrystalvioletstainingmethodwasusedtocomparethebiofilmformationabilitiesofCFT073and△FYUA.ResultsAFYUAgenedeletionstrain△FYUAwassuccessfullyconstructed.CFT073and△FYUA had the similar proliferation curves in Luria-Bertani liquid medium.The proliferation rate of △FYUAinsterileurinewasobviouslyslowerthanCFT073(P<0.05).ThereisnodifferenceincolonyformationbetweenCFT073and△FYUA.Biofilmformationabilityof△FYUAwasweakenedascomparedwithCFT073(P<0.05).ConclusionsFYUAgenemayplaysanimportantroleintheCFT073proliferationandbiofilmformationinsterileurine.
FYUA;geneknockout;urinarytractinfection; Escherichia coli
2014- 09- 15
2014- 12- 01
廣州醫科大學青年基金(2013A08)
1001-6325(2015)04-0502-06
R378.2+
A
*通信作者(corresponding author):wxm622@21cn.com
