不同規格圓口銅魚幼魚的化學組成和能量密度
2015-10-15 10:59:23董純唐會元潘磊楊志謝建軍喬曄
河北漁業 2015年10期
董純 唐會元 潘磊 楊志 謝建軍 喬曄


摘要:2014年11月-12月在金沙江收集體長(L)150~290 mm,體重(W)64.1~422.7 g的圓口銅魚23尾,根據體長將其分為4組,分別測定其化學組成,估算其能量密度。結果顯示:圓口銅魚幼魚含水量(WAT)、蛋白質含量(PRO)、脂肪含量(FAT)、灰分含量(ASH)和能量密度(E)占其濕重的百分比分別為6587%~69.99%,15.65%~18.17%,9.95%~14.07%,2.69%~3.80%,7.23~10.06 kJ/g。經統計分析,圓口銅魚幼魚含水量與其蛋白質含量、脂肪含量、灰分含量和能量密度均存在顯著的負線性關系。另外,圓口銅魚幼魚體長與其含水量存在顯著的負線性關系,而與蛋白質含量、脂肪含量、灰分含量和能量密度均存在顯著的正線性關系。結果表明,可以用圓口銅魚幼魚的含水量和體長估測其化學組成和能量密度,圓口銅魚高脂肪含量與其長距離繁殖洄游密切相關。
關鍵詞:體長;含水量;化學組成;能量密度;圓口銅魚
魚體化學組成是魚類生態學和生理學上的重要指標,與其攝食情況[1]、體大小[2]、棲息地[3]和生活季節[4]有關。魚類在各生長階段消耗的能量主要由蛋白質和脂肪提供,隨著魚體的生長,其化學組成會發生相應變化以適應能量需求。魚體化學組成和能量密度的測定較為復雜,工作量大,而魚體含水量與其化學組成和能量密度之間存在相關關系,因此,可以用含水量來估測魚體化學組成和能量密度[5]。
圓口銅魚(Coreius guichenoti)屬于鯉形目(Cytcriniformes)、鮈亞科(Gotioniae)、銅魚屬(Coreius),是長江中上游特有經濟魚類[6]。作為洄游性魚類,圓口銅魚幼魚在重慶江段生活3~4年,然后上游600~1 000 km到金沙江,在此完成整個生活史[7]。長距離的洄游需要消耗大量的能量[8-9],因此推測圓口銅魚洄游前儲備了大量的能量。本研究測定不同規格圓口銅魚幼魚的化學組成,探討魚體含水量和體長與其蛋白質含量、脂肪含量、灰分含量和能量密度的關系,為圓口銅魚洄游的能量生態學研究提供基礎數據。
1 材料與方法
1.1 實驗魚
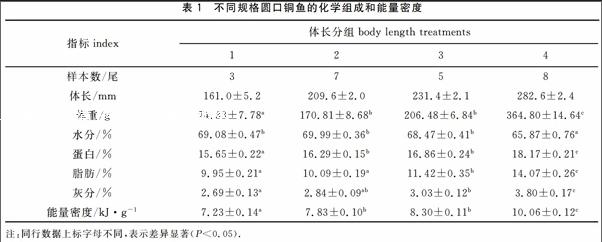
2014年11-12月在金沙江隨機采集實驗用23尾圓口銅魚幼魚,現場測定體重和體長,體重精確到0.1 g,體長精確到1 mm。實驗魚按體長分為4組,依次為150~170 mm、200~220 mm、220~240 mm、270~290 mm,各組平行樣品數分別為3、7、5、8。
1.2 樣品測定
每組樣品按照AOAC方法[10](1999)進行體成分分析。在105 ℃下烘樣品至恒重測定水分含量(WAT);采用凱氏定氮法測定樣品粗蛋白含量(PRO);采用索氏抽提法測定樣品粗脂肪含量(FAT);采用馬福爐焚燒法(550 ℃,5 h)測定樣品粗灰分含量(ASH)。碳水化合物含量占魚體比重低,約為0.5%,因此在魚體能量(E)計算中忽略不計[11],采用公式[12]:能量=蛋白×23.6+脂肪×39.5,計算所得。每個樣品測2個重復,當重復性不理想時重新測定,以保證結果的可靠性。
1.3 數據分析
使用軟件SPSS 17.0對數據進行單因素方差分析(One-way ANOVA),當方差不齊時,數據對數轉換后再進行統計分析。若差異顯著,則進行Tukey 多重比較(Tukey HSD test),顯著水平為P<0.05,并進行回歸分析。實驗所得數據表示為平均值±標準誤(mean ± S.E.)。
2 結果
2.1 體長與體重的關系
不同體長組的體長平均值依次為161.0、209.6、231.4和282.6 mm,魚體體重隨體長的增加而增加,其平均值依次為74.33、170.81、20648和364.80 g(表1)。以體長(L)為自變量,體重(W)為因變量,采用W=a Lb進行回歸運算,得W=6.73×10-5 L2.75(R2=0.96,P<005)(圖1)。
2.2 魚體化學組成和能量密度
不同體長組圓口銅魚幼魚化學組成和能量密度見表1,魚體含水量隨體長的增加而降低,而蛋白質含量、脂肪含量、灰分含量和能量密度均隨體長的增加而增加。進行統計分析得出,體長組4的含水量顯著高于其他體長組(P<0.05);體長組2和3的蛋白質含量和能量密度顯著低于體長組4,而顯著高于體長組1(P<0.05);體長組3的脂肪含量顯著低于體長組4,而顯著高于體長組1和2(P<0.05);體長組4的灰分含量顯著高于其他體長組(P<0.05)。
2.3 魚體化學組成和能量密度分別與其含水量的相關關系
圓口銅魚魚體含水量與蛋白質、脂肪、灰分含量和能量密度之間均存在顯著的負線性關系(P<0.05)。以含水量為自變量,分別以蛋白質、脂肪、灰分和能量密度作為因變量進行線性回歸分析,得到以下方程:PRO=-0.41 WAT+45.00(R2=0.56,P<0.05)(圖2);FAT=-0.72 WAT+60.53(R2=0.64,P<0.05)(圖3);ASH=-0.17 WAT+14.53(R2=0.41,P<0.05)(圖4);E=-0.39 WAT+34.86(R2=0.66,P<0.05)(圖5)。
2.4 魚體化學組成和能量密度分別與其體長的相關關系
圓口銅魚魚體體長與其含水量、蛋白質、脂肪、灰分含量和能量密度均存在顯著的相關關系(P<0.05)。以體長為自變量,分別以含水量、蛋白質、脂肪、灰分和能量密度作為因變量進行線性回歸分析,得到以下方程:WAT=-0.04 L+7776(R2=0.55,P<0.05)(圖6);PRO=0.03 L+10.26(R2=0.88,P<0.05)(圖7);FAT=0.05 L+0.62(R2=0.89,P<0.05)(圖8);ASH=001 L+0.18(R2=0.73,P<0.05)(圖9);E=003 L+2.63(R2=0.91,P<0.05)(圖10)。
3 討論
魚體體長和體重的關系是魚類生物學研究的基本內容之一,目前魚類體長和體重的關系一般用公式W=a Lb描述[13-15]。其中,a表示魚體單位體長的體重,b值反映魚類生長的生理學特性,b值等于3表示等速生長,b值大于或小于3則表示異速生長[16]。吳斌等[17](2008)研究體長40~80 mm的圓口銅魚,發現其b值為2.61,Luo等[18](2013)研究體長46~246 mm的圓口銅魚,b值為2.93,與本研究結果b值為2.75類似,均小于3,表明圓口銅魚幼魚一般呈異速生長。而體長90~320 mm的銅魚(Coreius heterodon),其b值為3.13,大于3[19]。黃真理等[13](1999)研究發現,長江流域大多魚類的b值在2.40~3.95之間,本研究結果b值在此范圍內。
魚體含水量的測定相對于魚體其他化學組成和能量密度的測定來說,操作簡單,工作量小。并且,很多研究表明,魚體含水量與魚體化學組成和能量密度之間密切相關[20-22],建議用魚體含水量來估測其化學組成和能量密度[23-24]。本研究結果顯示,圓口銅魚的含水量與蛋白質含量、脂肪含量、灰分含量和能量密度均存在顯著的負線性關系(P<0.05),即隨著含水量的增加魚體化學組成和能量密度均降低。這與吳斌等[17](2008)的研究結果一致,而Luo等[18](2013)的研究發現,圓口銅魚含水量與灰分含量存在正線性關系,與本研究結果相反。總之,本研究結果表明,可以用圓口銅魚幼魚的含水量粗略估計其化學組成和能量密度。另外,研究結果還顯示圓口銅魚的體長與含水量存在顯著的負線性關系(P<0.05),與蛋白質含量、脂肪含量、灰分含量和能量密度均存在顯著的正線性關系(P<0.05)。因此,也可用圓口銅魚幼魚的體長粗略估計其化學組成和能量密度。
脂肪是魚類主要的供能物質,影響著魚類的繁殖力和抵御饑餓的能力[25-26]。大多數洄游性魚類含有較高的脂肪含量,如洄游性魚類高首鱘(Acipenser transmontanus)[27]和北極紅點鮭(Salvelinus alpinus)[28]脂肪含量分別為2.6%~7.5% 和9.57%~11.31%。這是因為魚類在洄游時進行有氧活動,需要消耗大量的脂肪供給能量[29-30]。圓口銅魚作為一種河流洄游性魚類,在其生活史中有長距離的繁殖洄游活動,本研究結果,圓口銅魚幼魚脂肪含量高達9.95%~1407%,為其長距離繁殖洄游儲備了必要的能量。
參考文獻:
[1]Blake, R. W., Inglis, S. D., Chan, K. H. S. Growth, carcass composition and plasma growth hormone levels in cyclically fed rainbow trout [J]. Journal of Fish Biology, 2006, 69(3): 807-817
[2] Ali, A., Al-Ogaily, S. M., Al-Asgah, N. A., et al. Effect of feeding different protein to energy (P/E) ratios on the growth performance and body composition of Oreochromis niloticus fingerlings [J]. Journal of Applied Ichthyology, 2008, 24(1): 31-37
[3] Jonsson, N., Jonsson, B. Energy allocation among developmental stages, age groups, and types of Atlantic salmon (Salmo salar) spawners [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2003, 60(5): 506-516
[4] Dempson, J. B., Schwarz, C. J., Shears, M. Comparative proximate body composition of Atlantic salmon with emphasis on parr from fluvial and lacustrine habitats [J]. Journal of Fish Biology, 2004, 64(5): 1 257-1 271
[5] Robards, M. D., Anthony, J. A., Rose, G. A., et al. Changes in proximate composition and somatic energy content for Pacific sand lance (Ammodytes hexapterus) from Kachemak Bay, Alaska relative to maturity and season [J]. Journal of Experimental Marine Biology and Ecology, 1999, 242(2): 245-258
[6] Hartman, K. J., Margraf, F. J. Common relationships among proximate composition components in fishes [J]. Journal of Fish Biology, 2008, 73(10): 2 352-2 360
[7] Yang, S. R., Gao, X., Li, M. Z., et al. Interannual variations of the fish assemblage in the transitional zone of the Three Gorges Reservoir: persistence and stability [J]. Environmental Biology of Fishes, 2012, 93(2): 295-304
[8] Liu, L. H., Wu, G. X., Wang, Z. L. Reproduction ecology of Coreius heterodon (Bleeker) and Coreius guichenoti (Sauvage et Dabry) in the mainstream of the Changjiang River after the construction of Gezhouba Dam [J]. Acta Hydrobiological Sinica, 1990, 14(3): 205-215
[9] Kiessling, A., Lindahl-Kiessling, K., Kiessling, K. Energy utilization and metabolism in spawning migrating early start sockeye salmon (Oncorhynchus nerka): the migratory paradox [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2004, 61(3): 452-465
[10] Caudill, C. C., Daigle, W. R., Keefer, M. L., et al. Slow dam passage in adult Columbia River salmonids associated with unsuccessful migration: delayed negative effects of passage obstacles or condition-dependent mortality [J]? Canadian Journal of Fisheries and Aquatic Sciences, 2007, 64(7): 979-995
[11] AOAC (Association of Official Analytical Chemists). 1999. Official Methods of Analysis, 16th edition. AOAC, Washington, DC, USA
[12] Dawson, A. S., Grimm, A. S. Quantitative changes in the protein, lipid and energy content of the carcass ovaries and liver of adult female plaice (Pleuronects platessa L.) [J]. Journal of Fish Biology, 1980, 16(5): 493-504
[13] Brett, J. R., Groves, T. D. D. Physiological energetics in “Fish physiology”[M]. New York: Academic Press, 1979(8): 279-352
[14] Von Bertalanffy, L. Aquantitative theory of organic growth [J]. Hum Biol, 1938, 10: 181-213
[15] 彭姜嵐, 曹振東, 付世建. 鲇魚形態特征參數與體長關系及變異分析[J]. 重慶師范大學學報(自然科學版), 2007, 24(1): 69-71
[16] 黃真理, 常劍波. 魚類體長與體重關系中的分形特征[J]. 水生生物學報, 1999, 23(4): 330-335.
[17] Ricker, W. E. Computation and interpretation of biological statistics of fish populaions [J]. Bull Fish Res Boad Can, Bulletin of the Fisheries Research Board of Canada, 1975, 191: 1-382
[18] 吳斌, 羅毅平, 謝小軍. 圓口銅魚幼魚魚體的化學組成及能量密度[J]. 西南大學學報(自然科學版), 2008, 30(10): 62-67
[19] Luo, Y. P., Huang, Q. D., Zhang, Y. R., et al. Comparison of the body proximate compositions of juvenile bronze gudgeon (Coreius heterodon) and largemouth bronze gudgeon (C. guichenoti) in the upstream region of the Yangtze River [J]. SpringerPlus, 2013, 2(1): 75-80
[20] 王倩倩, 羅毅平. 銅魚魚體的化學組成及能量密度[J]. 重慶師范大學學報(自然科學版), 2010, 27(4): 21-25
[21] 王軍輝, 謝小軍. 瓦氏黃顙魚不同季節魚體的化學組成及能量密度預測模型[J]. 生態學報, 2003, 23(1): 122-129
[22] 雷思佳, 葉世洲, 李德尚, 等. 臺灣紅羅非魚幼魚水分含量與脂肪、蛋白質含量及比能值之間關系的研究[J]. 華中農業大學學報, 1999,18(4): 367-370
[23] Jonsson, N., Jonsson, B. Body composition and energy allocation in life-history stages of brown trout [J]. Journal of Fish Biology, 1998,53(6): 1306-1316
[24] Pangle, K. L., Sutton, T. M. Temporal changes in the relationship between condition indices and proximate composition of juvenile Coregonus artedi [J]. Journal of Fish Biology, 2005,66(4): 1 060-1 072
[25] Hutchings, J. A., Pickle, C. R., Mcgregor-Shaw, C. R., et al. Influence of sex, body size, and reproduction on over winter lipid depletion in brook trout [J]. Journal of Fish Biology, 1999, 55(5): 1 020-1 028
[26] Simpkins, D. G., Huber, W. A., Martinez, D. R., et al. Constraints of body size and swimming velocity on the ability of juvenile rainbow trout to endure periods without food [J]. Journal of Fish Biology, 2004,65(2): 530-544
[27] Cui, Y., Hung, S. S. O., Zhu, X. Effect of ration and body size on the energy budget of juvenile white sturgeon [J]. Journal of Fish Biology, 1996,49(5): 863-876
[28] Adams, C. E., Huntingford, F. A. Growth, maturation and reproductive investment in Arctic charr [J]. 1997,51(4): 750-759
[29] Hinch, S. G., Standen, E. M., Haeley, M. C., et al. Swimming patterns and behavior of upriver-migrating adult pink (Oncorhynchus gorbuscha) and sockege (O. nerka) salmon as assessed by EMG telemetry in the Fraser River British Columbia, Canada [J]. Hydrobiologia, 2002,483(1-3):147-160
[30] Bradford, R. G. Differential utilization of storage lipids and storage proteins by Northwest Atlantic herring (Clupea harengus Harengus) [J]. Journal of Fish Biology, 1993,43(6): 811-824
Abstract:To investigate the changes trend of chemical composition with the increasing body size in juvenile Coreius guichenoti, 23 specimens whose body length (L) and body weight (W) ranged 150~290 mm and 64.1~422.7 g respectively were collected from Jinsha River in November~December, 2014. The fish were divided into 4 groups according to the body length. Their chemical compositions were measured and energy density was estimated. The results showed that the contents of water (WAT), protein (PRO), lipid (FAT) and ash (ASH) of fish accounted for the wet weight 65.87%~69.99%, 15.65%~18.17%, 9.95%~14.07% and 2.69%~3.80%, respectively. The energy density (E) ranged 7.23~10.06 KJ/g. The statistical analysis displayed that PRO, FAT, ASH and E were in a significant negative linear correlation with WAT. In addition, WAT was in a significant negative linear correlation with the body length. However, PRO, FAT, ASH and E were in a significant positive linear correlation with the body length. The results suggested that the chemical composition and the content of energy density of Coreius guichenoti can be estimated by the water content and the body length. The fish in a high lipid is supposed to be closely correlated with its long distance migratory breeding.
Key words:Body length; Water content; Chemical composition; Energy density; Coreius guichenoti
