柵藻延遲發光的初步研究*
2015-10-18 11:46:14付加雷龐靖祥聶曉艷韓金祥1
生物醫學工程研究 2015年1期
關鍵詞:生物
付加雷,龐靖祥,聶曉艷,韓金祥1,△
(1. 山東中醫藥大學,山東 濟南 250355;2.山東省中醫藥研究院,山東 濟南 250014;3. 山東省醫學科學院,山東 濟南 250062)
1 引 言
與化學發光不同,所有生命系統均存在著超弱的光輻射,他們涉及的范圍極為廣泛:動物及其器官、組織、細胞等;植物及其根莖、花、果等;各種水藻;各種微生物,如細菌、酵母菌等。這種普遍存在于生物系統中的超弱光輻射被稱為“生物光子輻射”(biophoton emission)。其典型強度為100個光子/(s·cm2),光譜分布至少在200~800 nm內是連續的。生物光子輻射作為生命新陳代謝過程的一個產物,來自生物分子從高能態向低能態的躍遷[1]。大量的實驗結果表明,生物光子輻射對生物系統內部的變化及外界環境的影響有高度的敏感性,因此,通過對生物光子輻射的探測和分析能夠獲得生物系統內部的微觀信息,了解外界環境的微弱變化[2-4]。
柵藻是淡水中常見的浮游藻類,極喜在營養豐富的靜水中繁殖,其中許多種類對有機污染物具有較強的耐性,可在水質評價中作為指示生物[5-6]。鑒于柵藻的以上特性,為了探討柵藻生物光子輻射與生物學過程的關系,本研究采用YPMS-2生物光子測量儀對其生物光子輻射行為進行了初步探測,得到了一些有意義的結論。
2 材料和方法
2.1 供試生物
柵藻(Scenedesmus sp.) 購自中國科學院武漢水生生物研究所(編號FACHB-933)。
2.2 柵藻的培養
收到藻種后,首先將試管內藻液搖勻,在無菌操作下將藻液直接轉入滅菌后的玻璃三角瓶(50 ml)內,然后將玻璃三角瓶瓶口封好,放置在光照培養箱中培養。培養溫度25±1℃,光照條件2 000 Lux,時間設置12 h晝/12 h夜。培養20 d,藻種生長狀態良好,生物量明顯增多,在無菌條件下可再次轉接,再次轉接比例1∶5(藻液:培養基),取長勢良好的柵藻轉接入盛有BG11培養基的500 ml錐形瓶中,培養一段時間后,測定藻懸液的生物光子輻射。
2.3 柵藻生長濃度的測定[7-8]
藻類的生物量測定是藻類生長、生理生化、生態等方面研究的必要手段,藻類生物量測定方法很多,通常的顯微直接計數法,光密度(OD)測量法,葉綠素測量法,Counter 電子顯微計數法等,各有優缺點。盡管到目前為止,計數細胞個體數仍然是最準確,最令人滿意的方法之一,可以獲得最基本的種群信息,但是直接計數法不但工作量大,不同種類間由此估算的生物量差異也較大。相比之下,光密度法操作簡單,需要樣品量少,能夠實現快速測定。因此,為了準確、快速的測量柵藻生物量,本實驗分別測定其細胞密度及吸光度(A),并進行直線回歸分析,得出回歸方程:C=(A×11.995+0.1502) ×106個/ml(R2=0.9766)。按照此公式,只要測定出藻液的吸光度,就能計算出藻液濃度(C)。
2.4 柵藻延遲發光的測量
YPMS-2生物光子測量儀,購自荷蘭米盧娜公司(Netherlands Meluna Research Company)。
將制備好的藻懸液混勻后傾入4 cm×1 cm×1 cm石英比色杯中,先用紫外分光光度計測其吸光系數,然后快速置于YPMS-2生物光子測量儀的樣品暗室內,放置一定時間(約180 s),以消除外界光源影響,使柵藻混懸液生物光子輻射達到自發狀態,再立即用白LED光源激發,激發時間為10 s,測其延遲發光,一般測量時間為600 s。測量參數為:工作電壓1250 V,測量時間600 s,間隔時間1 s。測定光子計數率—時間曲線、光子計數與柵藻濃度的關系以及培養基對柵藻延遲發光的影響。
2.5 數據處理分析
將測得的結果輸入到OriginPro 9.1 軟件中,分別對實驗數據進行作圖與非線性(雙曲線)擬合,并分析處理。
3 結果與討論
3.1 噪聲、培養基與藻液延遲發光對比分析
提前2 h打開溫濕度控制系統,室溫恒定設置為20℃,濕度恒定設置為45%。打開YPMS-2生物光子測量儀,測量暗室的本底噪聲,然后放入盛有3 ml液體培養基BG11的石英比色杯,測量培養基BG11自發與延遲發光,結果見圖1。培養基BG11為含有無機鹽NaNO3、K2HPO4、MgSO4、CaCl2、ZnSO4等的無菌水溶液。噪聲的平均光子數為11.4/s,BG11自發的平均光子數為12.05/s,培養基BG11延遲發光從光子數60多個,1s后直接衰減至自發狀態(平均光子數12.24/s)。無機鹽水溶液培養基BG11的延遲發光幾乎沒有雙曲性弛豫,呈現為指數衰減狀態。

柵藻混懸液的延遲發光與培養基BG11延遲發光對比分析,見圖2。培養基BG11延遲發光從光子數60多個,1s后直接衰減至自發狀態(平均光子數12.24/s),與本底噪聲(平均光子數11.4/s)非常接近。藻懸液的延遲發光是從光子數6×104多個向下衰減,呈現雙曲線弛豫;而培養基BG11的延遲發光從60多個光子衰減,呈現指數衰減。相對于藻懸液延遲發光而言,培養基延遲發光幾乎可以忽略不計,即可以認為藻懸液的延遲發光就是藻體本身的延遲發光。
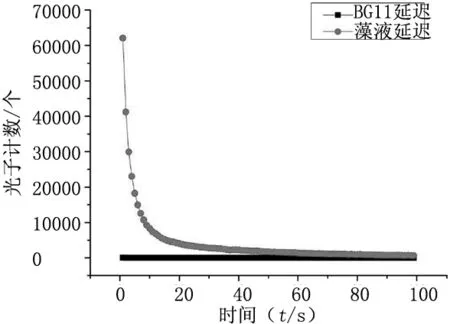
圖2 柵藻與培養基BG11延遲發光對比分析
3.2 柵藻的延遲發光光子計數率——時間曲線
將柵藻懸液混勻后傾入石英比色杯中,置于YPMS-2生物光子測量儀的樣品暗室內,放置一定時間(約180 s),待柵藻懸液生物光子輻射達到自發狀態后(自發狀態的平均光子數約30/s),立即用白LED光源激發柵藻,激發時間為10 s,測定其延遲發光,得到光子計數率——時間曲線,見圖3。

圖3 柵藻延遲發光光子計數——時間曲線
將雙曲函數公式:
I(t)=I0/(1+t/τ)β
(1)
轉換為公式:
y=a/(1+x/b)c
(2)
將公式(2)編輯入OriginPro 9.1 軟件,將柵藻的混懸液延遲發光測得的數據,與理論表達式(2)做雙曲線最佳擬合,其回歸方程為:
y=586714/(1+t/0.4)^1.25,R2=0.9986
將測定的柵藻混懸液延遲發光數據,與雙曲線函數(2)進行擬合,擬合關聯度高達99.86%,擬合的三個參數a(I0)=586714,b(τ)=0.4,c(β)=1.25。雙曲線衰減規律說明生物系統內的各個激發態分子之間是相互偶聯的,它們可能通過生物系統內存在的電磁場互相聯系,而這正是相干場的重要特征[9-10]。電磁輻射相干性:一部分自發的和光誘導的生物超弱發光的光子(量子),起源于生物系統內一個高度相干的電磁場,這種相干電磁場很可能是活組織內通訊聯絡的基礎[11-13],而恰恰可能是生物光子扮演著生物體內一種新型通訊信使的角色[14]。用雙曲線規律擬合柵藻的延遲發光數據曲線,擬合關聯度高達99%以上,這一結果支持了Popp 的相干性理論,也符合延遲發光弛豫動力學過程不能用指數函數描述的特點[15]。
雙曲函數公式(1)所含的三個參數I0、τ、β具有一定的物理意義:I0代表初始強度,它依賴于被測樣品的性質,同時與光照條件有關;τ 是一個特征時間,只與樣品自身的性質有關;β是一個指數因子,強烈控制弛豫的速率。這三個參數值定量地刻畫了被測樣品(在延遲發光意義上)的性質[4]。
3.3 柵藻藻液濃度對其延遲發光的影響
分別取生長旺盛的柵藻3 ml至5個14 ml離心管中,然后用液體培養基BG11分別稀釋至4、5、6、7、8 ml,取其中3 ml入石英比色杯中,測量其吸光度以及延遲發光。根據測得的藻液吸光度A,依據公式C=(A×11.995+0.1502) ×106(個/ml),計算每個稀釋比例藻懸液的濃度(個/mL),見表1。把不同濃度的藻懸液測得的延遲發光數據,分別與理論表達式(2)做雙曲線最佳擬合,得出不同濃度藻懸液延遲發光曲線擬合參數,見表2。
由表1 可知,隨著稀釋比例的降低,不同濃度的藻懸液的吸光度也降低,其相應的藻懸液濃度也明顯降低。
將盛有不同濃度藻懸液的石英比色杯,放入YPMS-2生物光子測量儀,測定其延遲發光。測得的不同濃度藻懸液數據利用OriginPro 9.1軟件進行畫圖處理,由圖4可知,單純從延遲發光曲線來看,不同濃度藻懸液之間幾乎沒有明顯的區別。將不同稀釋比例的藻懸液(不同濃度的藻懸液)延遲發光數據,分別與雙曲線函數(2)進行最佳擬合,擬合關聯度都高達99.60%以上,其擬合后的參數,見表2。實測初始強度,即盛有藻懸液的石英比色杯,放入YPMS-2生物光子測量儀的樣品暗室內,被白LED燈光源激發后,暗室快門打開,生物光子儀器初始記錄的光子數。
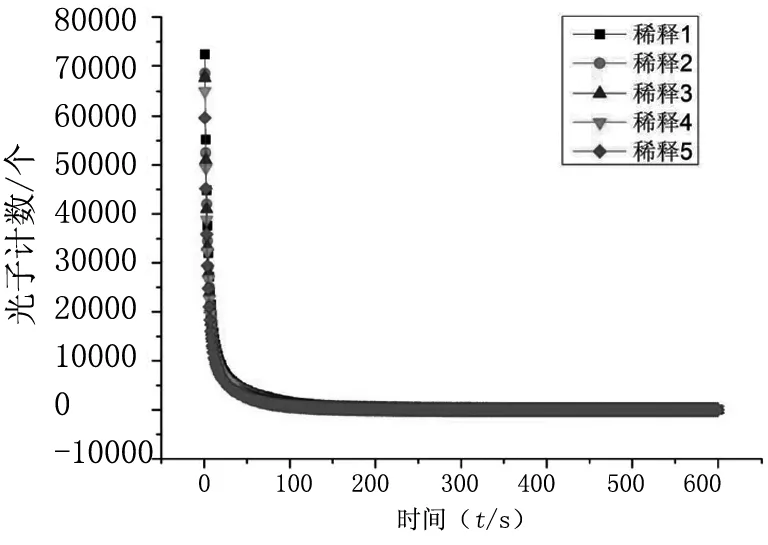
圖4 不同稀釋比例的藻液延遲發光曲線對比

表2 不同濃度的藻液延遲發光曲線擬合參數對比
由表2可知,隨著藻懸液濃度的降低,實測初始強度明顯降低;a(I0)值也明顯降低;c(β)值有升高趨勢;b(τ)先降低后又微弱升高。(1)a(I0)應該代表理論初始強度,它依賴于被測樣品的性質,同時與光照條件有關。在相同的白LED光源、相同時間激發下,不同濃度的藻懸液測得的延遲發光數據,經雙曲線函數(2)進行擬合后,得出的理論上的初始強度a(I0)(即擬合出的雙曲線的理論最高點),一般比實際測得的初始強度高出許多。實測初始強度和理論初始強度都與藻懸液濃度呈正相關,即隨著藻懸液濃度的降低,實測與理論初始強度也隨之降低。(2) c(β)是一個指數因子,它強烈控制弛豫的速率。隨著藻懸液濃度的降低,指數因子逐漸增大,即雙曲線的弛豫速率升高。換而言之,隨著藻懸液濃度的降低,其相應的延遲發光衰減變快。這一點從圖4也可以看出,即隨著藻懸液濃度越低,其延遲發光數據雙曲線拐點越接近中心坐標軸原點。(3)b(τ)是一個特征時間,只與樣品自身的性質有關,實驗結果顯示隨著柵藻濃度的降低,延遲發光的特征時間有升高的趨勢。生命系統相干性的另一表現是生物光子輻射的合作性[4],其基本特征為:輻射壽命與原子數成反比,輻射強度與原子數的平方成正比,這就是所謂的“超輻射”。復雜的生命系統是由無數的小的子系統構成,整個復雜系統的輻射壽命與子系統的數量成反比,輻射強度與子系統的數量的平方成正比,我們的結果初步驗證了該理論。
綜上所述,單純從延遲發光曲線來看,不同濃度藻懸液之間幾乎沒有明顯的區別;但通過雙曲函數擬合出來的三個參數(I0、τ、β),就能很明顯的把它們區別開,這三個參數攜帶著不同濃度柵藻和液體培養基的信息,即生物光子輻射攜帶著自身狀態和外界狀況的信息。這些實驗數據的獲得,為柵藻可作為液體環境的生物指示劑,奠定了一定的理論基礎。
4 結論
(1)柵藻與培養基延遲發光對比以及柵藻在液體培養基中延遲發光分析,表明柵藻具有較強生物光子輻射能力。
(2)柵藻的光子計數率—時間曲線回歸說明,用雙曲線函數擬合,其擬合關聯度高達99%以上,雙曲線規律為生物光子輻射的相干理論提供了證據。
(3) 柵藻延遲發光的初始強度I0,隨藻密度的增加而增強,兩個計量之間呈顯著正相關;隨著藻液濃度的降低,指數因子β逐漸增大,即雙曲線的弛豫速率升高,衰減變快;τ 是一個特征時間,與樣品自身的性質有關。隨著柵藻濃度的降低,延遲發光的特征時間有升高的趨勢,這一結果再次驗證了生物光子輻射相干性理論。
柵藻有較強的生物光子輻射能力,其延遲發光的雙曲線規律為生物光子輻射的相干性理論提供了證據。雙曲函數擬合出來的三個參數(I0、τ、β)有明顯差別,其中τ 是一個特征時間,與樣品自身的性質有關。隨著柵藻濃度的降低,延遲發光的特征時間有升高的趨勢,這一結果再次驗證了生物光子輻射相干性理論。這三個參數攜帶著不同濃度柵藻和液體培養基的信息,即生物光子輻射攜帶著自身狀態和外界狀況的信息。這些實驗數據的的獲得,為柵藻可作為液體環境的生物指示劑,奠定了一定的理論基礎。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
