番茄抗黃化曲葉病毒病品種在廣西南寧的自然抗病性表現
2015-11-25 08:32:02徐鵬超李戰彪秦碧霞謝慧婷崔麗賢蔡健和
植物保護 2015年2期
徐鵬超,李戰彪,秦碧霞,謝慧婷,崔麗賢,蔡健和*
(1. 廣西大學農學院,南寧 530004;2. 廣西作物病蟲害生物學重點實驗室,廣西農業科學院植物保護研究所,南寧 530007)
?
番茄抗黃化曲葉病毒病品種在廣西南寧的自然抗病性表現
徐鵬超1,2#,李戰彪2#,秦碧霞2,謝慧婷2,崔麗賢2,蔡健和2*
(1. 廣西大學農學院,南寧 530004;2. 廣西作物病蟲害生物學重點實驗室,廣西農業科學院植物保護研究所,南寧 530007)
近年來,廣西番茄黃化曲葉病毒病(TYLCD)發生較為嚴重,病原也比較復雜(發現4 種病毒且存在復合侵染現象),對廣西的番茄生產造成了很大威脅,種植抗病品種是防治該病的關鍵措施。本研究對中國農科院蔬菜花卉所、上海市農科院園藝所、浙江省農科院蔬菜所、江蘇省農科院蔬菜所、北京市農林科學院蔬菜研究中心和廣西大學提供的43 份抗TYLCD番茄品種在廣西南寧的自然抗病性進行了評價。采用自然誘發的方法,病圃的番茄混合感染中國番茄曲葉病毒與中國番木瓜曲葉病毒。試驗結果表明,表現高抗的品種有‘浙紅3號’、‘浙紅4號’、‘K22’、‘K64’等8個番茄品種,表現抗病和中抗的品種有‘紅貝貝’、‘春展56’等25 個品種,表現感病的品種有‘櫻紅1號’、‘K6’等5 個品種,表現高感的品種有‘金陵佳玉’、‘蘇粉11號’等5 個品種。本試驗結果為抗病番茄品種在廣西乃至全國的推廣和布局提供參考。
番茄黃化曲葉病毒病; 抗病品種; 自然抗病性
番茄黃化曲葉病(Tomato yellow leaf curl disease,TYLCD)是一類嚴重危害番茄生產的病害, 引起該病的病原在分類上都屬于雙生病毒科(Geminiviridae)菜豆金色花葉病毒屬(Begomovirus)。在自然條件下,該屬病毒主要通過煙粉虱(Bemisiatabaci)以持久方式傳播蔓延,因此也稱為粉虱傳雙生病毒(whitefly-transmitted geminivirus,WTGs)[1]。目前,這類病毒已在亞洲、非洲、地中海地區、中北美洲及加勒比海等地的眾多國家和地區造成嚴重危害,我國的廣西、廣東、云南、浙江、江蘇、山東、上海、北京等10 多個省份均有發生,對生產已造成嚴重危害[2-9]。
WTGs侵染番茄后,癥狀主要表現為植株生長緩慢或停滯,嚴重矮縮,頂部葉片黃化變小,葉片邊緣向上卷曲,無法正常開花結果;后期感染,染病植株僅上部葉片和新芽表現癥狀,結果量減少,果實變小。該病具有發病率高、危害大、防治難、傳播效率高等特點。目前,生產上主要的防治措施有:培育無病壯苗、黃板誘殺煙粉虱、化學農藥防治、換茬輪作,培育抗病品種等,其中培育抗病品種是防治該病的根本。前人已對番茄品種的抗病性進行了較多報道,田兆豐等[10]綜合分析了20 多個番茄品種對番茄黃化曲葉病毒的抗病性,篩選出了3 份抗病品種;矯曉陽等[11]利用5種雙生病毒的侵染性克隆對‘浙雜502’、‘浙粉701’、‘浙粉702’等3個品種進行了抗病性鑒定。結果發現,這3個品種對這5種雙生病毒均具有較好的抗性。孔令娟等[12]對6 個抗黃化曲葉病毒病的番茄品種抗性進行了評價,發現番茄品種種植地域不同,其抗病性表現會有所差異。目前,市售的番茄種子有較多的品種均標注為抗病品種,然而,由于地域及抗病基因和病毒種類的不同造成番茄品種的抗性表現有較大差異,筆者對中國農科院蔬菜花卉所等單位提供的抗TYLCD番茄品種在廣西南寧進行了煙粉虱自然接種抗病性評價。
1 材料與方法
1.1 材料
1.1.1 番茄品種來源
供試的番茄品種共43個,其中‘K6’、‘K22’、‘K36’、‘K54’、‘K61’、‘K64’、‘K72’ 由上海農科院園藝所提供;‘浙紅1號’、‘浙紅2號’、‘浙紅3號’、‘浙紅4號’、‘浙櫻粉1號’、‘浙櫻粉2號’等由浙江農科院蔬菜所提供;‘13H-Z2’、‘13H-Z4’、‘13H-Z5’、‘13H-Z11’、‘13H-Z12’、‘3156’、‘1361’、‘1362’、‘1363’、‘1364’ 由中國農科院蔬菜花卉所提供;‘金陵甜玉’、‘金陵秀玉’、‘金陵寶玉’、‘金陵佳玉’、‘金陵黛玉’、‘蘇粉11號’、‘蘇粉12號’、‘蘇粉13號’、‘JSTY13S41’、‘JSTY13S42’由江蘇農科院蔬菜所提供;‘紅貝貝’、‘佳紅8號’、‘春展56’、‘秋光140’、‘秋光277’由北京市農林科學院蔬菜研究中心提供;‘西大1號’、‘西大2號’、‘櫻黃1號’、‘櫻黃2號’、‘櫻紅1號’由廣西大學提供。感病對照為‘超級大明星’(市售),抗病對照為‘T11-7’,由南寧市立新公司提供。
1.1.2 試劑及PCR引物
2×TaqMaster Mix 購自北京康為世紀生物科技有限公司,SanPrep柱式DNA膠回收試劑盒購自生工生物工程(上海)股份有限公司,PGEM-T Easy Vector 試劑盒購自Promega,NaOH為國產分析純試劑。
PCR引物參考AV494/CoPR[13-14]委托生工生物工程(上海)股份有限公司合成。
1.2 方法
1.2.1 番茄培育及病毒接種
試驗地點為廣西農科院植保所網室內,網室常年種植有番茄黃化曲葉病毒病病株,且煙粉虱蟲量較大。所有的番茄試驗材料于2013年9月4日采用穴盤法進行育苗,待幼苗長至5~6 葉期于10月1日移栽至盆中,每個品種移栽20 株,移栽后,將盆栽苗放置于網室內進行自然接種,定期調查植株發病情況,10月19日進行第1 次調查,10月29日進行第2 次調查。
1.2.2 PCR檢測及序列分析
采集網室內發病的‘超級大明星’番茄樣品8份,用NaOH 粗提法進行DNA的提取[15],將提取液用0.1 mol/L Tris-HCl稀釋50 倍進行PCR擴增。所得PCR產物克隆至PGEM-T Easy Vector,進行序列測定,測序工作由北京諾賽基因組研究中心有限公司完成。
病情調查完成后,采集各品種未表現癥狀的番茄葉片各1份(如果一個品種中有多株未發病植株則作為混合樣采集),用NaOH粗提法提取各樣品的總DNA,將提取液用0.1 mol/L Tris-HCl稀釋50 倍進行PCR檢測。
1.2.3 病情調查與分析
每個番茄品種調查20 株,采用Lapidot 和Friedmann[16]的病情分級標準記錄病情和計算病情指數;按發病的嚴重程度劃分為0、1、3、5、7、9共6個等級,病情分級如下:
0級:無癥狀;
1級:頂端葉片的葉緣輕微發黃;
3級:羽狀復葉的小葉末端黃化和輕微卷曲;
5級:多數葉片黃化,卷曲,邊緣上翹;
7級:葉片顯著黃化,上翹,卷曲,植株矮化;
9級:葉片極度黃化,卷曲,植株嚴重矮化,甚至停止生長。
發病率(%)=(發病株數/調查總株數)×100;
病情指數(DI)=∑(級別代表值×本級病株數)/ (調查總株數×最高級代表值)×100;
抗病評價標準(參考葉青靜等[17]):
高抗(HR):0
2 結果與分析
2.1 病情調查及抗病性評價
番茄植株接種后,每周觀察番茄植株的發病情況,到第3周進行第1次調查,第5周進行第2次調查,計算各品種的病情指數并進行分析,參考抗病性評價標準進行抗病性鑒定(結果見表1)。
接種3 周后,除抗病對照未表現癥狀外,其余各番茄品種均表現出番茄黃化曲葉病毒病的典型癥狀;接種5 周后(表1),各番茄品種均有發病,發病最為嚴重的為‘金陵佳玉’與‘蘇粉11號’,病情指數分別達到100與81.94;其余各供試品種病情指數在3.47~79.17。其中感病對照‘超級大明星’發病率達100 %,病情指數達到73.61,抗病對照‘T11-7’發病率為5 %,病情指數為0.69。
從表1可以看出,攜帶Ty-3a基因的番茄品種抗性最好,大部分表現高抗,但也有例外,‘浙紅1號’等4個品種表現抗病和中抗,‘K6’和‘蘇粉11號’表現感病和高感;攜帶Ty-1/Ty-3 和Ty-3基因的番茄品種抗性表現中等;而攜帶Ty-2 基因的番茄品種抗性表現較差,表現抗、中抗、感病甚至高感。
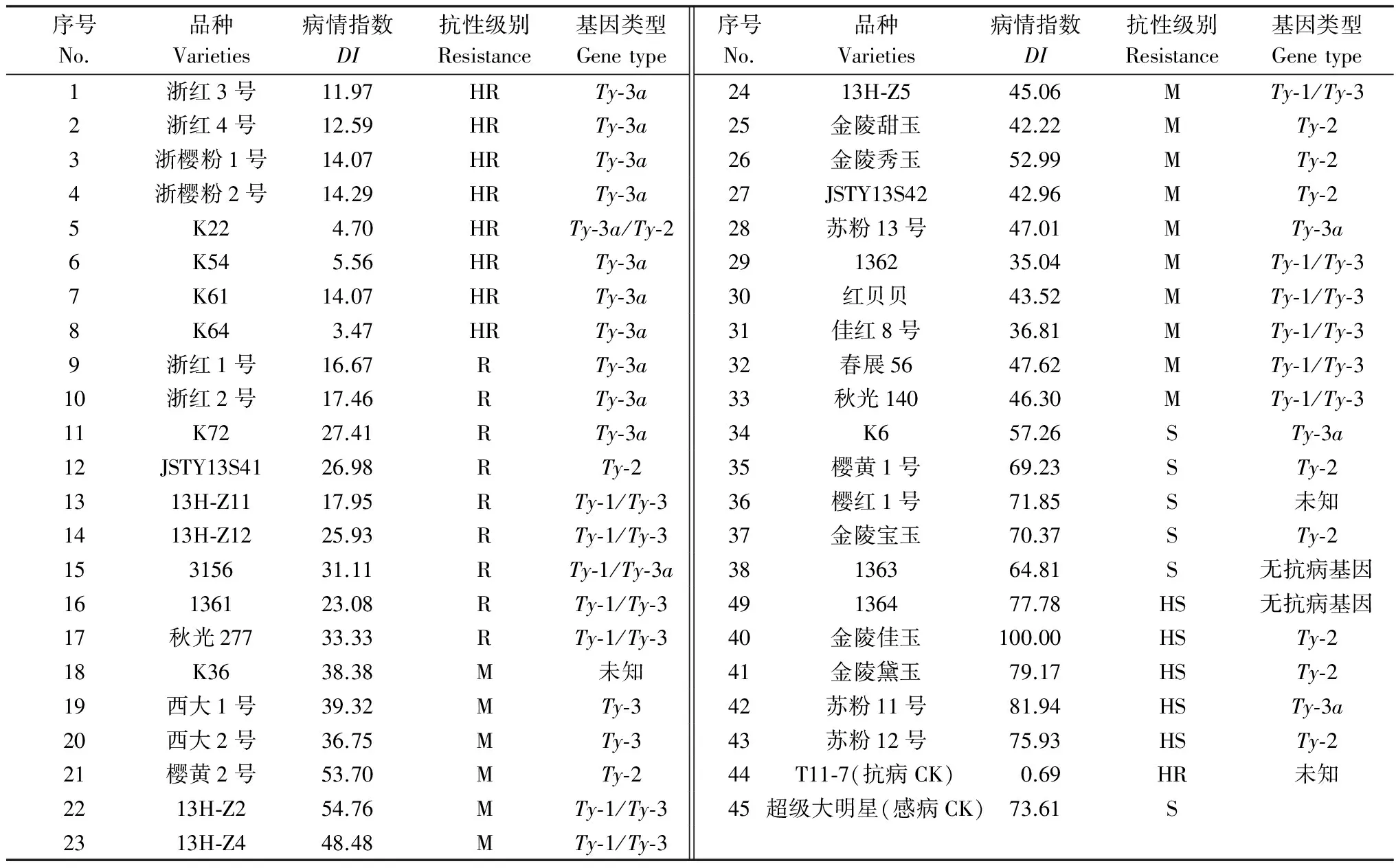
表1 番茄品種對番茄黃化曲葉病抗性評價結果
2.2 病毒種類分子鑒定
在自然接種區共采集8份感病的‘超級大明星’番茄植株葉片樣品,提取總DNA后,用AV494/CoPR引物進行擴增,PCR產物經純化后連接至載體進行序列測定,測序結果經GenBank,BLASTN分析表明,感病植株均為中國番茄曲葉病毒(TomatoleafcurlChinavirus,ToLCCNV)與中國番木瓜曲葉病毒(PapayaleafcurlChinavirus, PaLCuCNV)復合侵染。
2.3 未表現癥狀番茄植株的PCR檢測
各番茄品種中部分未表現癥狀的16 份番茄樣品PCR檢測結果表明,‘浙紅1號’、‘櫻粉1號’等未顯癥的番茄品種也攜帶病毒,僅有‘金陵甜玉’等幾個品種未檢測出病毒(圖1)。
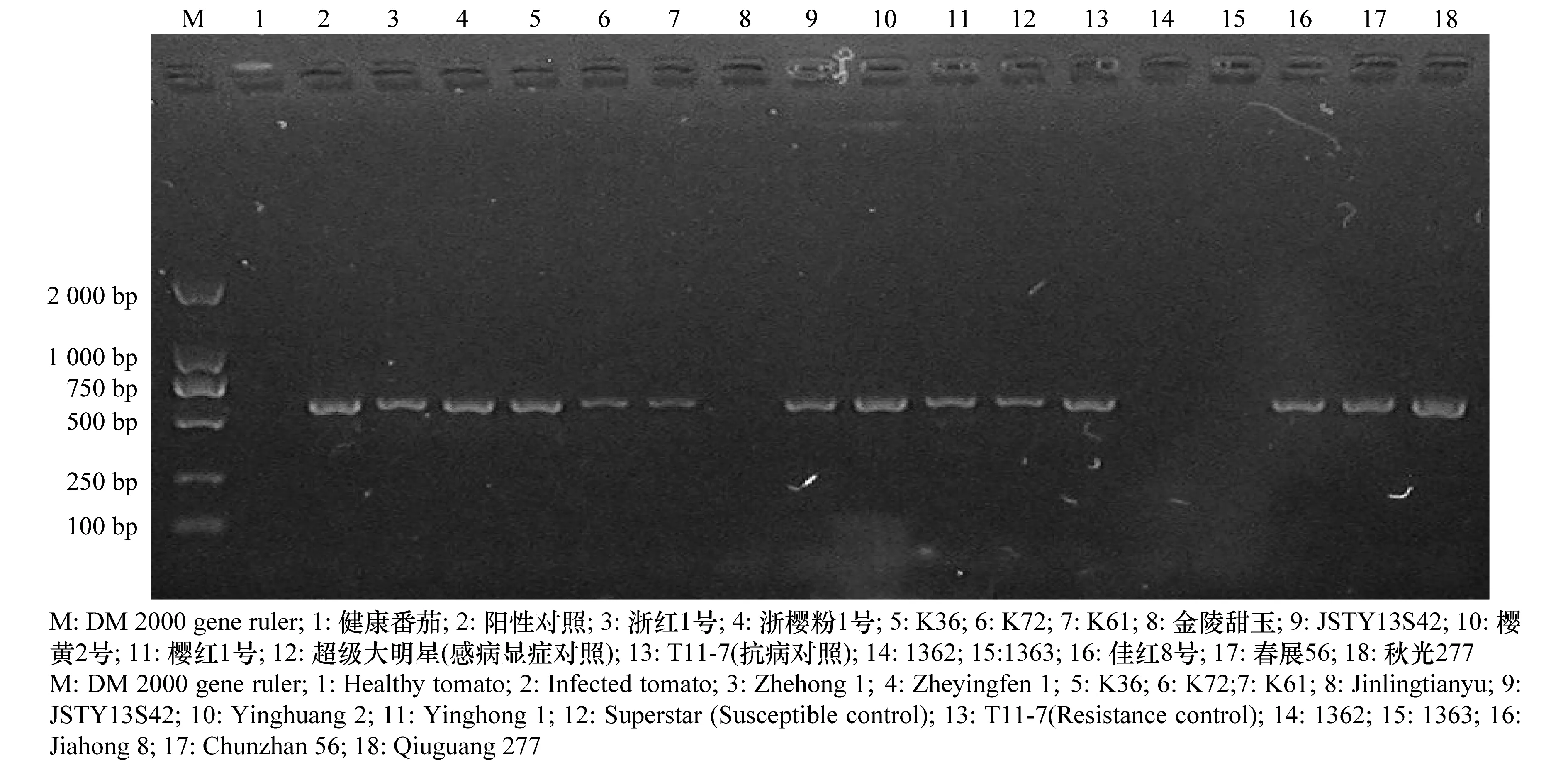
圖1 部分未顯癥番茄植株病毒檢測結果Fig.1 Electrophoretic analysis of PCR products from partial tomato plants
3 結論與討論
煙粉虱傳雙生病毒(WTGs)在自然界由于基因重組突變容易導致病毒變異,從而引起品種抗性喪失,導致病害暴發流行[18]。據報道,世界上引起番茄黃化曲葉病毒病的病原超過57種,侵染我國番茄的雙生病毒至少有10種[19]。江蘇、北京等地的番茄黃化曲葉病毒病主要病原為番茄黃化曲葉病毒(Tomatoyellowleafcurlvirus,TYLCV)[6,9],浙江省主要病原為TYLCV和臺灣番茄曲葉病毒(TomatoleafcurlTaiwanvirus,ToLCTwV)[7],廣西番茄雙生病毒發生頻率最高的是PaLCuCNV和ToLCCNV兩種。試驗結果表明,各番茄品種在廣西南寧的抗性表現與其在品種提供單位所在地的表現差異較大,上海農科院與浙江農科院提供的‘K22’、‘浙紅3號’等8個番茄品種表現為高抗,‘紅貝貝’、‘春展56’等25 個番茄品種僅表現抗病和中抗, ‘櫻紅1號’、‘K6’ ‘金陵佳玉’、‘蘇粉11號’等表現感病甚至高感。
番茄品種所攜帶的抗病基因與番茄品種的抗病性密切相關,含有Ty-1、Ty-2、Ty-3、Ty-4及Ty-5其中1 個抗性基因的番茄品系,對番茄黃化曲葉病毒病都有一定抗性,但抗性水平不等[16]。本研究中上海農科院與浙江農科院的番茄品種普遍攜帶Ty-3a抗性基因,江蘇農科院的番茄品種除‘蘇粉11號’與‘蘇粉13號’攜帶Ty-3a外,其余品種材料均攜帶Ty-2基因,從試驗結果可以看出,攜帶Ty-3a基因的番茄品種普遍對PaLCuCNV和ToLCCNV有較好的抗性,攜帶Ty-3 基因和攜帶Ty-1/Ty-3 基因的番茄品種對PaLCuCNV和ToLCCNV也表現出了較好的抗性,攜帶Ty-3 基因的番茄品種對PaLCuCNV有較好抗性;另外,攜帶Ty-2基因的番茄品種部分表現高感,部分表現中等及以上抗性,該研究結果與矯曉陽等[11]研究結果部分吻合,但含有Ty-3a的‘蘇粉11號’和‘K6’抗性表現較差,原因還有待查明。
本研究對部分未顯現癥狀的番茄植株進行了PCR檢測,檢測結果表明,這些番茄植株雖然未顯現癥狀,但是同樣受到病毒侵染,未發現免疫的番茄品種。
由于參試番茄株數偏少,接種壓力偏大,也未進行測產,試驗結果不一定完全反映各品種在田間的自然抗耐病性,僅供有關育種部門參考。
[1] Harrison B D, Robinson D J. Natural genomic and antigenic variation in whitefly-transmitted geminiviruses (Begomoviruses) [J]. Annual Review of Phytopathology,1999,37:369-398.
[2] 蔡健和,秦碧霞,朱桂寧,等.番茄黃化曲葉病毒病在廣西暴發的原因和防治策略[J]. 中國蔬菜, 2006(7): 47-48.
[3] 何自福,虞皓,毛明杰,等.中國臺灣番茄曲葉病毒侵染引起廣東番茄黃化曲葉病[J].農業生物技術學報,2007,15(1):119-123.
[4] 王玉,丁銘,楊莉,等. 侵染番茄的中國番木瓜曲葉病毒基因組結構特征[J]. 西南農業學報, 2010, 23(6):1917-1922.
[5] 褚棟,侯麗霞,劉國霞,等. 山東省局部地區番茄黃化曲葉病毒的分子鑒定[J]. 山東農業科學, 2010, 42(2):13-15.
[6] 趙統敏,余文貴,周益軍,等. 江蘇省番茄黃化曲葉病毒病 (TYLCD)的發生與診斷初報[J]. 江蘇農業學報, 2007, 23 (6):654-655.
[7] 袁偉,萬紅建,王榮青,等. 浙江省番茄黃化曲葉病毒的分子鑒定及序列分析[J].分子植物育種,2013,11(2):185-192.
[8] 宋晰,師迎春,張世晨,等. 北京地區番茄黃化曲葉病病毒分離物測定及株系的初步鑒定[J]. 植物病理學報, 2013,43 (2):113-119.
[9] 田守波,張輝,于力,等. 上海地區番茄黃化曲葉病癥狀多樣性與分子鑒定[J]. 上海農業學報, 2012, 28(3):1-5.
[10]田兆豐,劉偉成,謝歡,等. 不同番茄品種對番茄黃化曲葉病毒的抗病性鑒定[J]. 植物保護學報, 2013, 40(1):56-60.
[11]矯曉陽,周雪平,楊悅儉,等. 不同番茄品種對5種植物雙生病毒的抗病性鑒定[J]. 植物病理學報, 2013, 43(6):655-658.
[12]孔令娟,潘紅,陳紅輝,等. 番茄抗黃化曲葉病毒病品種篩選試驗[J]. 北方園藝, 2011(22):13-15.
[13]Wyatt S D, Brown J K. Detection of subgroup Ⅲ geminivirus isolates in leaf extracts by degenerate primers and polymerase chain reaction [J]. Phytopathology, 1996, 86:1288-1293.
[14]何自福,虞皓,羅方芳. 番茄煙粉虱傳雙生病毒PCR檢測[J]. 中國病毒學,2004,19(1):67-69.
[15]李正和,李桂新,謝艷,等. 云南番茄曲葉病是由煙草曲莖病毒引起的[J]. 病毒學報,2002,18(4):355-361.
[16]Lapidot M, Friedmann M. Breeding for resistance to whitefly-transmitted geminiviruses [J]. Annals of Applied Biology, 2002,140:109-127.
[17]葉青靜,周國治,王榮青,等. 番茄黃化曲葉病毒病抗性鑒定技術研究[J]. 分子植物育種, 2011, 9(2):210-217.
[18]Zhou X P, Liu Y L, Calvert L,et al. Evidence that DNA-A of a geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination [J]. Journal of General Virology,1997, 78:2101-2111.
[19]張輝.侵染我國番茄雙生病毒種類鑒定及致病性分析[D]. 杭州:浙江大學, 2009.
Natural resistance of TYLCD-resistant tomato varieties in Nanning,Guangxi
Xu Pengchao1,2, Li Zhanbiao2, Qin Bixia2, Xie Huiting2, Cui Lixian2,Cai Jianhe2
(1. College of Agriculture, Guangxi University, Nanning 530004,China; 2. Key Laboratory of Biology for Crop Diseases and Insect Pests,Institute of Plant Protection, Guangxi Academy of Agricultural Sciences, Nanning 530007, China)
Tomato yellow leaf curl disease (TYLCD) occurred seriously and cause severe economic losses in Guangxi during recent years. Application of resistant variety was the best way to control this disease. In this study, the natural resistances of 43 TYLCD-resistant tomato varieties collected from Chinese Academy of Agricultural Sciences, Shanghai Academy of Agricultural Sciences, Zhejiang Academy of Agricultural Sciences, Beijing Academy of Agricultural Sciences, Beijing Academy of Agriculture and Forestry Sciences and Guangxi University were determined by the methods ofBemisiatabaciinoculation naturally in Nanning City, Guangxi Zhuang Autonomous Region. The diseased tomato were mix-infected byTomatoleafcurlChinavirus(ToLCCNV)andPapayaleafcurlChinavirus(PaLCuCNV) in diseased nursery.The result showed that 8 varieties (including ‘Zhehong 3’, ‘Zhehong 4’,‘ K22’, ‘K64’ etc.) were highly resistant to TYLCD ; 25 varieties were resistant and moderately resistant, 5 varieties were susceptible, and 5 varieties were highly susceptible. The results could provide a theoretical reference for the extension of these TYLCD-resistant tomato varieties in Guangxi and other regions of China.
tomato yellow leaf curl disease; TYLCD-resistant tomato varieties; natural resistance
2014-03-21
2014-06-23
公益性行業(農業)科研專項(201003065);廣西農業科學院科技發展基金(桂農科2013JQ19);國家國際科技合作項目(2012DFA31220);廣西農業科學院基本科研業務專項(桂農科2011YZ15);廣西作物病蟲害生物學重點實驗室基金(13-051-47-ST-4);廣西特聘專家專項
S 432.21
B
10.3969/j.issn.0529-1542.2015.02.037
* 通信作者 E-mail: caijianhe@gxaas.net
# 對本文有同等貢獻,并列為第一作者
