鎘對河南華溪蟹卵黃磷蛋白在卵巢中表達(dá)含量的影響及ELISA法的建立
2015-11-29 02:45:58劉冬梅何永吉漢斯達(dá)姆斯
水生生物學(xué)報(bào) 2015年2期
關(guān)鍵詞:研究
楊 健 劉冬梅 何永吉 漢斯·達(dá)姆斯 王 蘭
?
鎘對河南華溪蟹卵黃磷蛋白在卵巢中表達(dá)含量的影響及ELISA法的建立
楊 健1劉冬梅2何永吉1漢斯·達(dá)姆斯3王 蘭1
(1. 山西大學(xué)生命科學(xué)學(xué)院, 太原 030006; 2. 山西醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院, 太原 030006; 3. 高雄醫(yī)學(xué)大學(xué)生命科學(xué)院, 高雄 80708)
為了探究鎘對河南華溪蟹()的卵子發(fā)生的影響, 通過凝膠過濾層析法純化了成熟卵巢中的卵黃磷蛋白(Vitellin, Vn), 采用SDS-聚丙烯酰胺凝膠電泳確定了Vn亞基數(shù)量及分子量, 以純化的Vn為抗原制備了Vn多克隆抗血清并建立了可靠的Vn含量測定的ELISA方法。采用氯化鎘體外亞慢性染毒的方法, 運(yùn)用Vn-ELISA方法檢測鎘暴露對卵巢組織中Vn含量的影響。實(shí)驗(yàn)設(shè)對照組和5.8 mg/L鎘處理組, 處理時(shí)間分別為10、15和20d。結(jié)果顯示, (1) SDS-聚丙烯酰胺凝膠電泳結(jié)果表明該蛋白由分子量為102、84和70 kD的3個(gè)亞基組成, 同時(shí)Western-blotting檢測表明, 抗血清對該蛋白具有較強(qiáng)的免疫特異性; (2) 在Vn-ELISA中, Vn抗體最佳稀釋倍數(shù)為1︰100000, 該方法的工作濃度范圍為125—2000 ng/mL, 定性檢測的靈敏度可達(dá)到15.62 ng/mL; 卵巢中Vn含量測定的結(jié)果中各批次間沒有顯著差異, 且平均變異系數(shù)分別僅為7.81%, 表示該法具有較高的穩(wěn)定性和重復(fù)性; (3) 鎘引起卵巢組織中Vn含量的顯著降低, 與鎘處理時(shí)間具有時(shí)間-效應(yīng)關(guān)系, 說明了鎘暴露可影響河南華溪蟹卵母細(xì)胞的成熟并對卵子發(fā)生產(chǎn)生抑制作用, 進(jìn)而影響卵巢的正常發(fā)育。
河南華溪蟹; 卵黃磷蛋白; SDS-聚丙烯酰胺凝膠電泳; 酶聯(lián)免疫吸附法; 鎘
鎘在工業(yè)中的大量使用導(dǎo)致其成為環(huán)境中大量存在的非必需金屬元素之一, 同時(shí)也成為水體污染中一類典型的環(huán)境污染物。本實(shí)驗(yàn)室早期研究發(fā)現(xiàn)鎘可以在河南華溪蟹()卵巢、肝胰腺、鰓以及肌肉等組織中富集, 導(dǎo)致生物體產(chǎn)生急性或慢性中毒, 造成組織器官出現(xiàn)不同程度的損害, 如鎘誘導(dǎo)的細(xì)胞壞死、細(xì)胞凋亡和脂質(zhì)過氧化反應(yīng)[1—4]。韓托等[5]研究發(fā)現(xiàn)在鎘富集的水域生物個(gè)體大量死亡, 這直接導(dǎo)致水生生物種群數(shù)量和生物多樣性顯著降低。此外, 研究發(fā)現(xiàn)鎘會(huì)對魚類、蟹類和鳥類的脂質(zhì)、糖類和蛋白質(zhì)代謝產(chǎn)生影響, 從而影響物種的遷徙和繁殖[6—8]。同時(shí)由于鎘的半衰期較長, 通過消化及呼吸系統(tǒng)進(jìn)入機(jī)體后造成的損傷是永久的, 這將會(huì)長期影響水生環(huán)境中物種的繁殖過程進(jìn)而影響生物種群及生態(tài)系統(tǒng), 因此探究可反映污染物對生物早期影響的繁殖參數(shù)是水生生態(tài)毒理學(xué)研究中急需要解決的問題。
卵子發(fā)生是甲殼動(dòng)物卵巢發(fā)育中的一個(gè)重要階段, 表現(xiàn)為卵母細(xì)胞中卵黃蛋白原(Vitellogenin, Vg)的形成和積累[9]。在卵巢中, Vg會(huì)通過進(jìn)一步的修飾和加工形成包含有Vg、脂類、碳水化合物和色素等物質(zhì)的卵黃磷蛋白(Vitellin, Vn), 并在卵母細(xì)胞中積累形成卵黃體。Vn是甲殼動(dòng)物卵子存活和胚胎發(fā)育所需營養(yǎng)物質(zhì)的主要來源, 卵巢中Vn積累的多少直接影響到其后生殖過程和幼體質(zhì)量。因此, Vn可作為研究甲殼動(dòng)物繁殖的重要生物指標(biāo)[10]。此外, 甲殼動(dòng)物的卵黃發(fā)生過程由于受內(nèi)分泌系統(tǒng)、外部環(huán)境的調(diào)節(jié)而具有較高的復(fù)雜性, 目前對其內(nèi)在的調(diào)控機(jī)制研究已成為甲殼動(dòng)物研究熱點(diǎn)和難點(diǎn)之一[11, 12]。甲殼動(dòng)物Vn已有的研究發(fā)現(xiàn), Vn含量與卵巢發(fā)育階段具有一定的相關(guān)性, 因此機(jī)體組織中Vn含量測定對于研究鎘對甲殼動(dòng)物卵巢發(fā)育和卵子發(fā)生的影響具有重要意義。
酶聯(lián)免疫吸附法(Enzyme-linked immunoabsor-bent assay, ELISA) 已被廣泛用于甲殼動(dòng)物Vn含量的測定[13, 14]。不同的甲殼動(dòng)物在Vn亞基數(shù)目、空間結(jié)構(gòu)和分子量上通常存在一定差異, 因此需要針對不同物種制備具有特異性的Vn抗體, 并在此基礎(chǔ)上建立適合該物種Vn-ELISA方法, 為進(jìn)一步研究甲殼動(dòng)物卵黃發(fā)生及鎘對卵巢發(fā)育的影響提供重要依據(jù)。目前, 對于甲殼動(dòng)物卵子發(fā)生和鎘對卵巢發(fā)育的影響已進(jìn)行了較為廣泛的研究[13—16]。然而河南華溪蟹在這方面的研究少有報(bào)道。因此, 當(dāng)前迫切需要建立一種較為可靠的方法以測定鎘處理后河南華溪蟹卵巢中Vn含量。本研究分離并純化了河南華溪蟹的Vn, 制備了Vn多克隆抗體, 優(yōu)化了Vn測定的ELISA參數(shù), 該方法不僅可以用于研究鎘對河南華溪蟹卵巢中卵子發(fā)生的影響, 還可作為深入研究河南華溪蟹生殖調(diào)控和環(huán)境監(jiān)測的理論依據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)材料
河南華溪蟹(簡稱“溪蟹”)于2013年10月購自山西省太原市五龍口水產(chǎn)批發(fā)市場。選取卵巢發(fā)育成熟的雌蟹置于實(shí)驗(yàn)室水族缸中暫養(yǎng)三周后進(jìn)行活體解剖后將卵巢保存于–80℃冰箱中備用。
1.2 卵黃磷蛋白分離和純化
卵巢粗提液的制備溪蟹成熟卵巢中加入10倍體積預(yù)冷的勻漿緩沖液(0.02 mol/L Tris-HCl, 0.1 mol/L NaCl, 0.01% EDTA, 0.1 mmol/L PMSF)冰浴勻漿, 在4℃12000×下離心30min后取橘黃色上清液并加入等體積的飽和硫酸銨溶液, 冰浴1h后再次在4℃12000×下離心30min, 棄上清液, 向離心管中加入2 mL的PBS緩沖液(0.1 mol/L KH2PO4, 0.1 mol/L Na2HPO4·12H2O, 0.1 mol/L NaCl, 0.1 mol/L KCl, 0.01% EDTA, pH 7.7)溶解沉淀, 然后重復(fù)“沉淀-離心-復(fù)溶”的步驟5次, 最后將沉淀物溶解于1 mL PBS緩沖液中用于凝膠過濾層析分離純化蛋白。
卵黃磷蛋白提取采用凝膠柱體積為260 mL (直徑×高度=2.6 cm×100 cm, 上海廈美生物科技發(fā)展有限公司生產(chǎn)), 裝入220 mL凝膠(型號SephacrylTMS 100 HR, 瑞典Pharmacia公司生產(chǎn)), 純化前首先采用PBS緩沖液溶液平衡凝膠柱, 流速16 mL/h, 平衡1h。用注射器上樣2 mL Vn提取液, 流速為16 mL/h, 使用UVD-680-3紫外檢測器(上海金達(dá)生化儀器廠)在280 nm條件下檢測洗脫液的吸收值。當(dāng)出現(xiàn)蛋白組分時(shí)開始收集, 每2 mL收集1管, 洗脫液于–80℃保存?zhèn)溆谩J褂肧pectraMax M5多功能酶標(biāo)儀(美國Molecular Devices分子儀器公司)對提純后的卵黃蛋白溶液進(jìn)行連續(xù)光譜掃描(700—200 nm)以檢測類胡蘿卜素。
1.3 卵黃磷蛋白亞基數(shù)量及分子量分析
將純化蛋白進(jìn)行變性聚丙烯酰胺電泳(SDS-PAGE, 分離膠質(zhì)量濃度為10%, 濃縮膠質(zhì)量濃度為5%), 然后用考馬斯亮藍(lán)R-250染色后, 在Tanon-2500 (上海天能公司)凝膠成像儀拍照, 通過與蛋白標(biāo)準(zhǔn)比對并采用BandScan 5.0軟件以確定溪蟹Vn各亞基的分子量。
1.4 抗體制備和免疫印跡
選用6—8周齡雌性BALB/c小鼠作為免疫動(dòng)物, 暫養(yǎng)1周后將純化的Vn溶液與QuickAntibody等體積混合后用于免疫注射。每只小鼠于后腿小腿肌肉首次免疫注射100 μL (免疫抗原量25 μg)的抗原乳化液, 此后第21天再次注射100 μL抗原乳化液進(jìn)行加強(qiáng)免疫, 第35天小鼠摘除眼球取血, 室溫靜置1h待血液凝固后, 于4℃12000×離心10min后取上清, 加入0.02% NaN3, 無菌分裝于–80℃。
采用蛋白免疫印跡(Western-Blotting)檢驗(yàn)抗體和抗原免疫反應(yīng)的特異性。首先將純化的Vn進(jìn)行SDS-PAGE電泳, 電泳條件同前。電泳結(jié)束后的凝膠在72 V電壓下轉(zhuǎn)膜1h。將鼠多克隆抗體按 1︰5000稀釋, 加樣體積為1 mL, 室溫孵育1h; 加入1︰2000稀釋的羊抗鼠IgG-IRP(Catalog No. HSA0004, 上海麥約爾生物技術(shù)有限公司生產(chǎn))作為二抗, 室溫孵育1h后用1 mL PBST (10 mmol/L PBS, 0.05% Tween-20, pH 7.2)洗滌3次, 每次10min; 采用DAB辣根過氧化物酶顯色試劑盒(德國Roche公司)顯色3min后采用Tanon-2500凝膠成像儀拍照。
1.5 Vn抗體效價(jià)檢測和Vn-ELISA建立
采用間接包被法ELISA確定鼠多克隆Vn抗體的有效稀釋倍數(shù)并建立Vn-ELISA標(biāo)準(zhǔn)曲線。主要包括抗原包被、封閉、加抗體、顯色、反應(yīng)終止和讀數(shù)等步驟。(1)包被: 將純化的Vn用包被緩沖液(0.05 mol/L Na2CO3, 0.05 mol/L NaHCO3, pH=9. 6)溶解成1 μg/mL, 然后按每孔100 μL進(jìn)行包被, 4℃過夜后去除包被液; (2)封閉: 用200 μL的PBST重復(fù)清洗3次后, 每孔加入200 μL的封閉液(1%牛血清白蛋白)封閉2h, 然后用PBST洗滌3次; (3)加抗體: 將純化后的Vn抗血清按1︰1000、1︰10000、1︰100000和1︰20000用1%的BSA進(jìn)行稀釋, 每孔加樣100 μL, 同時(shí)以不含Vn抗體的1% BSA為陰性對照, 37℃反應(yīng)2h, 每孔用200 μL PBST洗滌3次后加100 μL稀釋的羊抗鼠IgG-HRP作為二抗, 37℃放置1h后用PBST洗滌3次; (4)顯色: 加入100 μL TMB顯色液(北京索萊寶科技有限公司), 室溫避光放置30min; (5)反應(yīng)終止: 加50 μL終止液(2 mol/L H2SO4溶液); (6)讀數(shù): 采用SpectraMax M5多功能酶標(biāo)儀測定450 nm的吸光度。
在上述實(shí)驗(yàn)的基礎(chǔ)上進(jìn)行標(biāo)準(zhǔn)曲線制備, Vn稀釋液的質(zhì)量濃度分別為7.81、15.62、31.25、62.5、125、250、500、1000、2000和4000 ng/mL, 每個(gè)質(zhì)量濃度組各重復(fù)3孔, 同時(shí)設(shè)置3個(gè)陰性對照, 在酶標(biāo)儀上讀取450。根據(jù)線性關(guān)系和變異系數(shù), 選取具有良好重復(fù)性和質(zhì)量濃度相關(guān)性的Vn濃度范圍繪制標(biāo)準(zhǔn)曲線。
1.6 驗(yàn)證性實(shí)驗(yàn)
隨機(jī)選擇5只雌性溪蟹, 分別稱重0. 1 g左右的卵巢組織, 加入10倍體積預(yù)冷的勻漿緩沖液進(jìn)行勻漿, 將勻漿液在4℃、12000×下離心15min, 取上清液0.1 mL用于ELISA測定。將勻漿液用包被緩沖液稀釋后用于ELISA測定; 該實(shí)驗(yàn)重復(fù)進(jìn)行3次, 然后分析批次間的誤差及變異系數(shù), 從而驗(yàn)證本實(shí)驗(yàn)建立ELISA方法的可靠性和穩(wěn)定性。
1.7 樣本染毒處理
選取處于卵巢發(fā)育期的成年雌性溪蟹進(jìn)行染毒處理。實(shí)驗(yàn)設(shè)對照組和5.8 mg/L鎘染毒組, 體外暴露10、15 和20d后取樣。各染毒時(shí)間組隨機(jī)選取5只溪蟹迅速解剖, 取出卵巢組織后采用Vn- ELISA法以測定其Vn含量。每個(gè)實(shí)驗(yàn)進(jìn)行3次獨(dú)立的重復(fù)。
1.8 數(shù)據(jù)分析
采用SPSS 17.0軟件(SPSS Inc., Chicago, USA)對實(shí)驗(yàn)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析, 數(shù)據(jù)采用平均值±標(biāo)準(zhǔn)差表示。分別采用Kolmogorov-Smirnov和Levene法進(jìn)行數(shù)據(jù)分布和方差齊性檢驗(yàn), 采用單因素方差分析中的Dunnett’s post-hoc 法對實(shí)驗(yàn)結(jié)果進(jìn)行方差分析。采用Test (independent samples Test)比較不同Vn抗體稀釋倍數(shù)、不同Vn質(zhì)量濃度和不同測定批次對結(jié)果是否存在顯著差異, LSD法進(jìn)行染毒組與對照組兩兩比較, 以<0.05為差異顯著標(biāo)準(zhǔn)。
2 結(jié)果
2.1 河南華溪蟹Vn的分離純化和亞基組成
溪蟹卵巢勻漿液凝膠柱層析結(jié)果顯示成熟卵巢勻漿液主要含有1個(gè)蛋白質(zhì)峰, 洗脫時(shí)間在175—190min時(shí)該蛋白組分大量被洗脫出峰。由于甲殼動(dòng)物卵黃磷蛋白含有較多類胡羅卜素, 連續(xù)光譜掃描的結(jié)果顯示卵黃磷蛋白在470 nm處有明顯的光吸收, 故該蛋白峰可能為溪蟹的Vn。SDS-聚丙烯酰胺凝膠電泳對該蛋白的進(jìn)一步分析發(fā)現(xiàn)(圖1), 該蛋白由3個(gè)亞基組成, 他們的分子量分別為102、84和70 kD。進(jìn)一步通過WesternBlotting實(shí)驗(yàn)表明, 該蛋白及3個(gè)蛋白亞基均可與溪蟹Vn的多克隆抗體產(chǎn)生特異性免疫反應(yīng)(圖1)。
1. 標(biāo)準(zhǔn)蛋白; 2. 卵黃磷蛋白0.6 mg/mL; 3. 卵黃磷蛋白 0.8 mg/mL; 4. 卵黃磷蛋白Western-blotting
1. Molecular mass markers; 2. vitellin 0.6 mg/mL; 3. vitellin 0.8 mg/mL; 4. Vn Western-blotting
2.2 河南華溪蟹Vn抗體效價(jià)檢測及標(biāo)準(zhǔn)曲線優(yōu)化
由表1可見不同稀釋倍數(shù)的溪蟹Vn抗體對ELISA讀數(shù)的影響。抗體稀釋1000—200000倍時(shí),450讀數(shù)均顯著高于陰性對照且各濃度組差異顯著。同時(shí)抗體稀釋20萬倍時(shí),450讀數(shù)與陰性對照呈顯著性差異, 這說明該抗體具體較高的效價(jià)。由于抗體稀釋10萬倍時(shí),450的差異系數(shù)僅為4.18%均低于各組, 因此本實(shí)驗(yàn)條件下的Vn抗血清最佳稀釋倍數(shù)為1︰100000。在上述優(yōu)化的Vn抗血清條件下, 將不同質(zhì)量濃度的Vn包被液按樣品直接包被法測定450值。
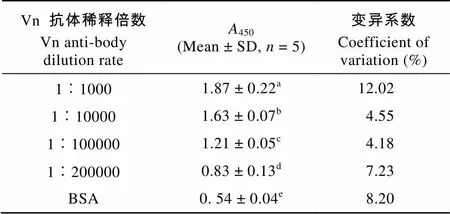
表1 河南華溪蟹Vn抗體不同稀釋倍數(shù)的ELISA反應(yīng)的A450值
注: 同列數(shù)據(jù)肩標(biāo)含有不同上標(biāo)字母為差異顯著 (< 0.05);代表樣品量, 下同
Note: Values within a column having different superscript letter are significantly different (< 0.05);means sample number. The same applies bellow
表2結(jié)果顯示在Vn濃度在7.81—4000 ng/mL時(shí),450值差異顯著。由于Vn質(zhì)量濃度為7.81 ng/mL時(shí),450值與陰性對照相比不存在顯著差異。而當(dāng)Vn質(zhì)量濃度為15. 62 ng/mL時(shí),450值顯著高于7.81 ng/mL濃度組和陰性對照, 因此該方法檢測Vn可檢出值為15. 62 ng/mL左右。
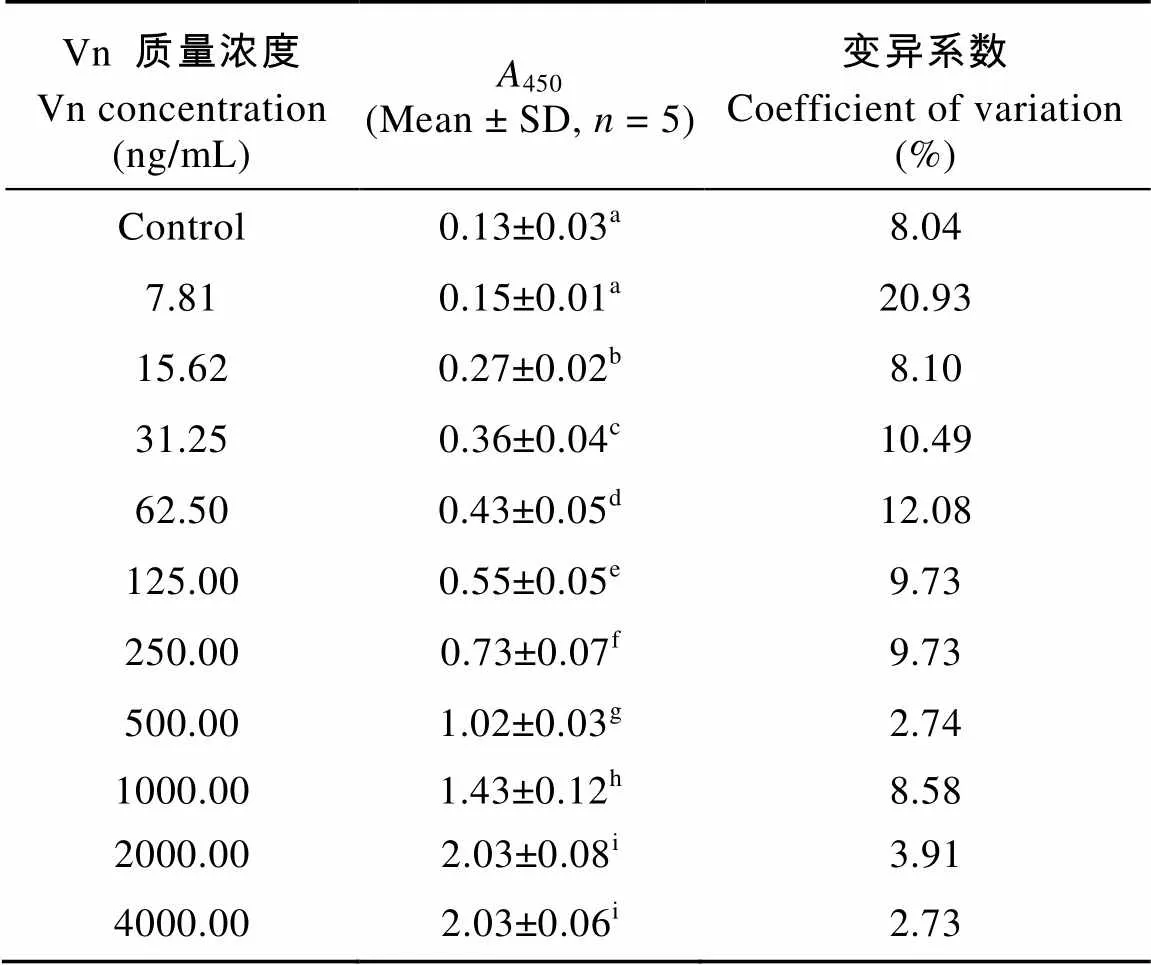
表2 不同Vn質(zhì)量濃度的A450值
Vn質(zhì)量濃度在7.81—62.5 ng/mL時(shí),450的變異系數(shù)較大不符合標(biāo)準(zhǔn)曲線的要求, 且在4000 ng/mL與2000 ng/mL Vn濃度組相比,450值不存在顯著差異, 因此選擇Vn質(zhì)量濃度為125、250、500、1000和2000 ng/mL制作標(biāo)準(zhǔn)曲線。圖2結(jié)果表明在該范圍內(nèi)Vn質(zhì)量濃度和450線性關(guān)系良好(= 0. 0008+ 0. 5565,2=0. 9763,< 0. 001,和分別代表Vn質(zhì)量濃度和450), 且各點(diǎn)的變異系數(shù)較低, 因此該標(biāo)準(zhǔn)曲線的重復(fù)性和線性相關(guān)性均符合要求, Vn工作濃度范圍為125—2000 ng/mL。
2.3 驗(yàn)證性試驗(yàn)
根據(jù)建立的Vn-ELISA標(biāo)準(zhǔn)曲線, 隨機(jī)取5只雌性溪蟹的卵巢組織進(jìn)行驗(yàn)證性試驗(yàn)并重復(fù)3次。表3為驗(yàn)證性試驗(yàn)的結(jié)果, 批次間平均變異系數(shù)僅為7. 81%。批次間測定的Vn含量無顯著差異, 因此本實(shí)驗(yàn)建立的ELISA測定方法具有較高的穩(wěn)定性和可重復(fù)性。
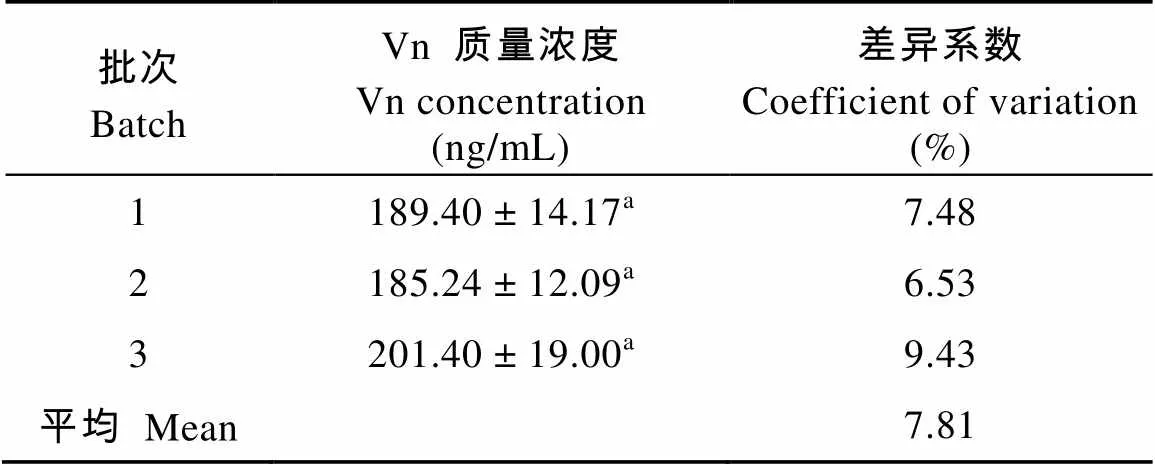
表3 河南華溪蟹卵巢中Vn的含量
2.4 鎘對河南華溪蟹卵巢中Vn含量的影響
圖3所示, 鎘染毒后, 各處理組卵巢中Vn含量與對照組比較有顯著性降低(< 0.05), 隨著處理時(shí)間的延長, 卵巢中Vn含量呈現(xiàn)逐漸降低的趨勢, 并在鎘處理20d染毒組中達(dá)到最低值(31.42 ± 7.25) mg/g。
3 討論
在大多數(shù)的甲殼動(dòng)物中僅存在一種形式的卵黃磷蛋白[17, 18], 然而Serrano-Pinto等[19]和Laino等[20]的研究發(fā)現(xiàn)了兩種存在形式的卵黃磷蛋白。在本研究中, 通過凝膠過濾層析法純化了溪蟹成熟卵巢中的Vn。在經(jīng)過SDS處理后發(fā)現(xiàn)該蛋白分成3條多肽鏈, 分子量分別為102、84和70 kD, 并且各亞基彼此差異明顯, 這與其他甲殼動(dòng)物的研究結(jié)果存在一定的差異。大量研究表明甲殼動(dòng)物卵黃磷蛋白的分子量通常在200—700 kD, 且多由2—6個(gè)亞基組成, 如中華絨螯蟹()卵黃磷蛋白的分子量為520 kD, 兩種亞基的分子量分別為97和74 kD[13]; 銹斑()Vn兩個(gè)亞基分子量分別為105和76 kD[21]; 陸蟹() 3個(gè)亞基分子量分別為115、105和85 kD[22]; 日本沼蝦() 3個(gè)亞基分子量分別為110、96和89 kD[23]; 三疣梭子蟹卵黃磷蛋白的亞基數(shù)為三個(gè), 分子量分別為100、75和66 kD[14]; 克氏原螯蝦()Vn分子量為481 kD, 有六個(gè)亞基 (198、176、132、111、92、82 kD)[24]。
以分離純化后的溪蟹Vn為抗原, 制備了相應(yīng)的Vn抗血清。Western-blotting實(shí)驗(yàn)表明該抗血清可以特異地識別溪蟹中的Vn。這種較高的特異性確保該抗體可用于溪蟹體內(nèi)Vn的免疫學(xué)研究以建立一種Vn-ELISA法定量測定組織中的Vn的含量。當(dāng)前的研究表明本文建立的Vn-ELISA法具有較高的靈敏度, 最低可檢測到15.62 ng/mL的Vn, 其最佳的工作濃度范圍為125—2000 ng/mL。而在其他甲殼動(dòng)物中, 由Vn特異多抗血清建立的Vn-ELISA研究結(jié)果與本文的基本類似。Volz等[25]在對橈足類動(dòng)物的研究中建立Vn-ELISA工作范圍為31—1000 ng/mL, 精度為1.9 ng/mL。在中華絨螯蟹的研究中, Chen等建立了一個(gè)類似的Vn-ELISA, 其測定范圍為8—500 ng/mL[13]。牡蠣()Vn-ELISA系統(tǒng)中的工作濃度為 10—400 ng/mL[26]。驗(yàn)證性實(shí)驗(yàn)結(jié)果顯示出本實(shí)驗(yàn)所建立的河南華溪蟹Vn-ELISA法在批次內(nèi)和批次間不存在顯著性差異, 同時(shí)具有較低的差異系數(shù), 因此該法具有較高的穩(wěn)定性和可重復(fù)性。與當(dāng)前已經(jīng)發(fā)表的研究結(jié)果比較, 本實(shí)驗(yàn)所建立的Vn-ELISA工作范圍是可信和敏感的。該法所具有的較高的精確度使得可以在不同時(shí)間直接進(jìn)行樣本間卵黃磷蛋白含量的比較。為了使該法可以應(yīng)用與不同卵黃磷蛋白濃度的測定, 本文將純化好的卵黃磷蛋白稀釋為不同的濃度以建立一條標(biāo)準(zhǔn)曲線, 這樣可以定量測定在經(jīng)過鎘處理后溪蟹組織中的卵黃磷蛋白。本實(shí)驗(yàn)結(jié)果發(fā)現(xiàn), 在經(jīng)過鎘的亞慢性染毒處理之后, 溪蟹卵巢中的Vn含量顯著降低, 這表明鎘暴露可抑制溪蟹中Vn的合成和積累。本實(shí)驗(yàn)室早期研究發(fā)現(xiàn)鎘處理后溪蟹卵巢指數(shù)顯著降低, 并認(rèn)為這可能是溪蟹體內(nèi)存儲(chǔ)的大量能量物質(zhì)被分解以用于能量合成的結(jié)果[27]。Vn作為溪蟹卵巢中重要的營養(yǎng)物質(zhì), 它的減少則可能是導(dǎo)致卵巢發(fā)育異常的重要原因。Revathi等和Rodriguez等研究認(rèn)為鎘對Vn合成的抑制作用可導(dǎo)致卵巢中卵母細(xì)胞直徑減小, 這可延緩卵母細(xì)胞的成熟和卵子的發(fā)生, 進(jìn)而影響卵巢的正常發(fā)育和種群的繁殖[28, 29]。
在大部分的昆蟲中, 卵黃磷蛋白在卵巢外先合成為卵黃磷蛋白的前體蛋白, 然后通過血液循環(huán)系統(tǒng)輸送到卵巢, 然后被卵母細(xì)胞吸收。當(dāng)前關(guān)于甲殼動(dòng)物卵黃蛋白原的合成位點(diǎn)還沒有搞清楚, 一直都是研究的熱點(diǎn)。關(guān)于十足類甲殼動(dòng)物Vg合成部位的研究結(jié)果顯示, 肝胰腺和卵巢是公認(rèn)的主要的合成部位, 其次是血細(xì)胞和皮下脂肪體, 因此在甲殼動(dòng)物中Vg的合成可能是內(nèi)源的或外源的或兩者兼有。在一些種類中, Vg在卵巢中合成, 如短溝對蝦()[30]。而在另一些甲殼動(dòng)物種類中, Vg在肝胰腺中合成, 如一種溪蟹()[31]。對于不同種甲殼動(dòng)物, Vg合成部位的研究結(jié)果不盡相同。當(dāng)前關(guān)于河南華溪蟹卵黃磷蛋白的研究較少, 因此, 可以在本文研究的基礎(chǔ)上, 進(jìn)一步探討河南華溪蟹卵黃發(fā)生部位和鎘對Vn合成影響的機(jī)理。
致謝:感謝鄒恩民教授對論文的修改給予的指導(dǎo)幫助。
[1] Wang Q, Wang L, Xi Y Y,. The acute toxicity and accumulation of Cd2+in freshwater crab[J].(Natural Science Edition), 2003, 26(2): 176—178 [王茜, 王蘭, 席玉英, 等. 鎘對長江華溪蟹的急性毒性與積累. 山西大學(xué)學(xué)報(bào)(自然科學(xué)版), 2003, 26(2): 176—178]
[2] Wang L, Xu T, Lei W,. Cadmium-induced oxidative stress and apoptotic changes in the testis of freshwater crab,[J]., 2011, 6(11): e27853
[3] Liu D, Yan B, Yang J,. Mitochondrial pathway of apoptosis in the hepatopancreas of the freshwater crabexposed to cadmium [J]., 2011, 105(3-4): 394—402
[4] Jin F F, Xu T, Qin S J,. Effects of cadmium on hepatopancreas mitochondrial antioxidant enzyme activity and lipid peroxidation level in freshwater crab[J]., 2011, 35(6): 1019—1024 [金芬芬, 徐團(tuán), 秦圣娟, 等. 鎘對長江華溪蟹肝胰腺線粒體抗氧化酶活力和脂質(zhì)過氧化水平的影響. 水生生物學(xué)報(bào), 2011, 35(6): 1019—1024]
[5] Han T, Wang Q, Wang L. Ecological investigation of freshwater crab and river pollution in the basin of Qinhe River [J]., 2008, 27(5): 804—806 [韓托, 王茜, 王蘭. 沁河干流底棲動(dòng)物生態(tài)調(diào)查與重金屬積累. 四川動(dòng)物, 2008, 27(5): 804—806]
[6] De Smet H, Blust R. Stress responses and changes in protein metabolism in carpduring cadmium exposure [J]., 2001, 48(3): 255—262
[7] Pierron F, Baudrimont M, Bossy A,. Impairment of lipid storage by cadmium in the European eel () [J]., 2007, 81(3): 304—311
[8] Lucia M, André J M, Gonzalez P,. Effect of dietary cadmium on lipid metabolism and storage of aquatic bird[J]., 2010, 19(1): 163—170
[9] Wu X G, Yao G G, Yang X Z,. A study on the ovarian development ofin East China Sea during the first reproductive cycle [J]., 2007, 29(4): 120—127 [吳旭干, 姚桂桂, 楊筱珍, 等. 東海三疣梭子蟹第一次卵巢發(fā)育規(guī)律的研究. 海洋學(xué)報(bào), 2007, 29(4): 120—127]
[10] Ghanawi J, Patrick Saoud I. Molting, reproductive biology, and hatchery management of redclaw crayfish[J]., 2012, 358-359: 183—195
[11] Kim D H, Kim S K, Choi J H,. The effects of manipulating water temperature, photoperiod and eyestalk ablation on gonad maturation of the swimming crab,[J]., 2010, 83(2): 129—141
[12] Tinikul Y, Poljaroen J, Tinikul R. Effects of gonadotropin-releasing hormones and dopamine on ovarian maturation in the Pacific white shrimp,, and their presence in the ovary during ovarian development [J]., 2014, 420-421: 79—88
[13] Chen L, Jiang H, Zhou Z,. Purification of vitellin from the ovary of Chinese mitten-handed crab () and development of an antivitellin ELISA [J].,, 2004, 138(3): 305—311
[14] Zhang Y, Wu X G, Yang F,. Purification of vitellin and ELISA determination of vitellin of swinming crab () [J]., 2011, 35(8): 1146—1157 [張艷, 吳旭干, 楊帆, 等. 三疣梭子蟹卵黃磷蛋白純化及其ELISA測定方法. 水產(chǎn)學(xué)報(bào), 2011, 35(8): 1146—1157]
[15] Allner B, von der G?nna S, Griebeler E M,Reproductive functions of wild fish as bioindicators of reproductive toxicants in the aquatic environment [J]., 2010, 17(2): 505—518
[16] Jones I, Kille P, Sweeney G. Cadmium delays growth hormone expression during rainbow trout development [J]., 2001, 59(4): 1015—1022
[17] Zheng H P, Zhang Q, Liu H L,. Cloning and expression of vitellogenin (Vg) gene and its correlations with total carotenoids content and total antioxidant capacity in noble scallop(Bivalve: Pectinidae) [J]., 2012, 366—367: 46—53
[18] Jia X, Chen Y, Zou Z,. Characterization and expression profile of vitellogenin gene from[J]., 2013, 520(2): 119—130
[19] Serrano-Pinto V, Vazquez-Boucard C, Villarreal-Colmenares H. Yolk proteins during ovary and egg development of mature female freshwater crayfish () [J]., 2003, 134(1): 33—43
[20] Laino A, Cunningham M L, Heras H,. Isolation and characterization of two vitellins from eggs of the spider(Araneae: Sparassidae).,, 2011, 158(2): 142—148
[21] Mak A S C, Choi C L, Tiu S H K,. Vitellogenesis in the red crab: Hepatopancreas speciflic expression and farnesoic acid stimulation of vitellogenin gene expression [J]., 2005, 70(3): 288—300
[22] Pateraki L E, Stratakis E. Synthesis and organization of vitellogenin and vitellin molecules form the land crab[J].,, 2000, 125(1): 53—61
[23] Mu S M. Yolk proteins ofand effects on their accumulation by unilateral eyestalk ablation [D]. Thesis for Master of Science. Hebei University, 2004 [穆淑梅. 日本沼蝦()卵黃蛋白及切除眼柄對其積累影響的研究. 碩士學(xué)位論文, 河北大學(xué), 2004]
[24] Xie S, An J H, Liang X H,. Partial characteriza-tion of vitellin of crayfish,[J].(Natural Science Edition), 2006, 26(2): 199—202 [謝松, 安建宏, 梁曉輝, 等. 克氏原螯蝦卵黃蛋白的部分生化性質(zhì). 河北大學(xué)學(xué)報(bào)(自然科學(xué)版), 2006, 26(2): 199—202]
[25] Volz D C, Chandler G T. An enzyme-linked immunosorbent assay for lipovitellin quantification in copepods: a screening tool for endocrine toxicity [J]., 2004, 23(2): 298—305
[26] Arcos F G, Ibarra A M, Rodríguez-Jaramillo M C,. Quantification of vitellin/vitellogenin-like proteins in the oyster(Hertlein 1951) as a tool to predict the degree of gonad maturity [J]., 2009, 40(6): 644—655
[27] Yang J, Liu D, Jing W,. Effects of Cadmium on lipid storage and metabolism in the freshwater crab[J]., 2013, 8(10): e7756
[28] Revathi P, Vasanthi LA, Munuswamy N. Effect of cadmium on the ovarian development in the freshwater prawn(De Man) [J]., 2011, 74(4): 623—629
[29] Rodríguez E M, Lopez Grego L S, Fingerman M. Inhibition of ovarian growth by cadmium, in the fiddler crab(Decapoda, Ocypodidae) [J]., 2000, 46(2): 202—206
[30] Avarre J C, Michelis R, Tietz A,. Relationship between vitellogenin and vitellin in a marine shrimp () and molecular characterization of vitellogenin complementary DNAs [J]., 2003, 69(1): 355—364
[31] Girish B P, Swetha C H, Reddy P S. Hepatopancreas but not ovary is the site of vitellogenin synthesis in female fresh water crab,[J]., 2014, 447(2): 323—327
ESTABLISHED OF ELISA METHOD OF VITELLIN FROM FRESHWATER CRAB () AND EFFECT OF CADMIUM ON VITELLIN ACCUMULATION IN OVARY
Yang Jian1, Liu Dong-mei2, He Yong-ji1, Hans-Uwe Dahms3and Wang Lan1
(1. School of Life Science, Shanxi University, Taiyuan 030006, China; 2. School of Basic Medical,Shanxi Medical University, Taiyuan 030006, China; 3. School of Life Science, Kaohsiung Medical University, Kaohsiung 80708,Taiwan, China)
To explore the effect of cadmium (Cd) on oogenesis of freshwater crab (), vitellin (Vn) and the method of Vn-ELISA was studied and established. Vn was purified from mature ovaries ofby gel filtration chromatography. Using SDS- polyacrylamide gel electrophoresis (SDS-PAGE), the molecular weight and the quantity of Vn subunit were determined. Based on the purified Vn and Vn anti-serum, the reaction parameters for an enzyme-linked immunosorbent assay (ELISA) was developed. To explore the effect of cadmium on the Vn in the ovaries, freshwater crab.weretreated with 5.8 mg/L Cd for 10, 15, and 20d. The results showed that Vn was composed of three subunits (116, 66 and 45 kD). Western-blotting confirmed the polypeptides had a specific reactivity with mouse polyclonal Vn anti-serum. The optimal dilution rates of Vn anti-serum were shown to be 1︰100000. According to these optimal parameters, the standard linear equation was established for the determination of Vn concentration with a valid range of 125—2000 ng/mL, and qualitative detection can be 15.62 ng/mL. In verification experiments, Vn content in the ovaries showed no significant differences, and the mean coefficients of variation of inter-assay were 7.81%, suggesting that the developed ELISA of this study was precise, stable and repeatable. Moreover, Cd caused a time-dependend down-regulation of Vn level, and showed significant effects of Cd on vitellogenesis, which suggest that Cd slows down oocyte maturation and vitellogenesis in
; Vitellin; SDS-polyacrylamide gel electrophoresis; Enzyme-linked immunosorbent assay; Cadmium
10.7541/2015.38
X174
A
1000-3207(2015)02-0287-07
2014-09-04;
2014-12-07
國家自然科學(xué)基金(No. 30870267, No. 30970361); 山西省回國留學(xué)人員科研項(xiàng)目(No. 2010015); 山西省普通高校特色重點(diǎn)學(xué)科建設(shè)項(xiàng)目(No. 2011-SXDX-SWX-003)資助
楊健(1988—), 男, 山西長治人; 博士; 主要從事環(huán)境重金屬污染研究方向。E-mail: yangjian333001@126.com
王蘭(1960—), 女, 山西太原人; 教授; 主要從事典型重金屬污染的細(xì)胞與分子機(jī)制研究方向。E-mail: lanwang@sxu.edu.cn
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
