短角異劍水蚤對克氏雙鋸魚稚魚成活、生長和脂肪酸組成的影響
2015-12-15 03:35:22劉新富賈玉東張和森
海洋科學 2015年4期
孟 振, 胡 鵬, 劉新富, 賈玉東, 劉 濱, 鮑 鷹, 張和森
(1.青島市海水魚類種子工程與生物技術重點實驗室 中國水產科學研究院 黃海水產研究所, 山東 青島266071; 2.中國科學院 海洋研究所, 山東 青島 266071; 3.青島通用水產養殖有限公司, 山東 青島 266404)
海洋橈足類種類多、分布范圍廣、群體數量大,在海洋食物鏈中, 是構成海水魚類仔稚魚餌料生物的主要組成部分[1-3]。人工育苗實驗中, 無論使用野外拖網采集的包含大量橈足類的浮游動物還是使用人工培養的橈足類, 都獲得較好的生長效果和較高的成活率[4-8]。尤為重要的是, 橈足類每個種類的生活史含有12個不同規格的發育階段, 提供了較寬規格幅度的生物餌料[3-6], 而且營養豐富, 特別是不飽和脂肪酸含量高, 彌補了現行的海水魚類生物餌料體系(微藻-輪蟲-鹵蟲無節幼體)個體大小范圍有限和營養供給不足的缺陷, 可以成功應用于培育輪蟲和鹵蟲無法飼養的經濟魚類, 如小口裂的石斑魚類[9]和大口裂的金槍魚[10]等。因此, 篩選合適的橈足類品種進行人工培養, 并探討其對海水魚類早期發育階段的餌料價值和投喂效果, 對于解決許多名貴魚類的苗種培育難題、提高海水魚類人工培育苗種的質量和成活率都具有極為重要的意義。
短角異劍水蚤(Apocyclops royi)是一種廣泛分布的海洋近岸小型橈足類[11], 是臺灣培育石斑魚苗常用的橈足類品種[12], 對其生理生態[13-14]的研究, 表明該品種是適于人工培養的小型餌料橈足類之一,但國內對該品種的人工培育研究尚處于起步階段[15],也未見應用于海水魚類育苗的報道。本實驗在收集、分離和純化培養短角異劍水蚤的基礎上[16], 以重要的海水觀賞魚——克氏雙鋸魚(Amphiprion clarkii,俗稱紅雙帶小丑魚)稚魚為實驗對象, 研究了短角異劍水蚤對其生長、成活和脂肪酸組成的影響, 以驗證短角異劍水蚤的餌料價值和投喂效果。
1 材料和方法
1.1 實驗材料
短角異劍水蚤于2011年4月取自江蘇省贛榆市一輪蟲養殖池, 取回后置于150 L PVC水槽中暫養,分離、純化后以濃縮海水小球藻為餌料培養1個月。隨后轉移至 3 m3的露天水泥池中充氣培養, 每日投喂20 mL濃縮海水小球藻, 換水量0.5 m3/d。實驗時,以350目篩絹過濾收集, 計數、消毒后投喂。
克氏雙鋸魚(Amphiprion clarkii)親魚由青島絢海水族科技有限公司人工培育、光溫調控性腺成熟[17]。
褶皺臂尾輪蟲(Brachionus plicatilis)由青島通用水產養殖有限公司提供, 在室內 PVC水槽中充氣培養, 投喂仔稚魚前用濃縮小球藻強化 24 h, 過濾收集, 消毒后定量投喂。濃縮海水小球藻(Chlorellasp.)、鹵蟲(Artemiasp.)卵均購自青島越洋進出口有限公司, 鹵蟲卵經孵化、去殼、小球藻營養強化、消毒后定量投喂。
1.2 實驗方法
1.2.1 克氏雙鋸魚受精卵孵化及初孵仔魚收集
受精卵在原水槽中孵化, 孵化水溫 26℃±1℃,pH值 7.8, 鹽度 30, 微充氣, 溶解氧高于 5.6 mg/L,光照強度1 000 lx, 光照周期L∶D=14 h∶10 h。待全部仔魚孵出2 h內, 根據初孵仔魚的趨光性, 光誘導聚集, 采收后將初孵仔魚平均分到 2個 50 L的PVC水槽中進行投喂實驗。采用不同批次受精卵重復實驗一次。
1.2.2 克氏雙鋸魚仔稚魚投喂實驗
2批次仔稚魚培育水質條件同上, 每天吸底并換水2次, 10 d前每次換水量25%, 10~20 d換水量50%,同時統計各實驗組死魚數目。從孵化后1~7 d投喂褶皺臂尾輪蟲, 日投喂2次, 投喂量10個/mL, 同時添加海水小球藻以調控水質顏色。從5 d至實驗結束(20 d)分別投喂鹵蟲無節幼體(A1/A2組)和橈足類(C1/C2組)。A1/A2、C1/C2組鹵蟲無節幼體和橈足類投喂密度均為5個/mL, 日投喂2次。
1.2.3 鹵蟲無節幼體、橈足類和克氏雙鋸魚稚魚脂肪酸的測定
1.2.3.1 樣品采集
鹵蟲無節幼體、橈足類: 鹵蟲無節幼體孵化8 h后, 投喂濃縮海水小球藻營養強化10 h, 饑餓12 h,分離卵殼, 經200目篩絹用蒸餾水沖洗過濾。橈足類以200目篩網收集轉入新鮮海水中, 饑餓12 h清空消化道內的單胞藻, 篩濾掉排出的廢物, 用蒸餾水沖洗。樣品冷凍干燥24 h。
克氏雙鋸魚稚魚: 取20 d稚魚20條, 轉入新鮮海水, 饑餓24 h清空消化道內餌料, 用蒸餾水沖洗,樣品冷凍干燥24 h。
1.2.3.2 樣品處理
樣品處理方法參考劉光興等[18], 取凍干樣品30 mg,加入1 mol/L KOH-甲醇3 mL, 在80℃水浴鍋中皂化10 min, 加入2 mol/L HCl-甲醇溶液3 mL, 在80℃水浴鍋中酯化反應10 min; 加入正己烷1 mL, 振蕩萃取, 靜止分層, 吸取上清液約 1 mL放入離心管中,低速離心5 min, 取1 μL進樣測定。
1.2.3.3 脂肪酸測定
使用美國安捷倫公司7890A氣相色譜儀分析測定脂肪酸的含量, 用面積歸一法計算各種脂肪酸占總脂肪酸的百分含量。
1.3 數據處理

其中Lt為實驗結束時稚魚體長(mm),L0為實驗開始時初孵仔魚體長(mm),t為實驗時間(d)。
采用SPSS17.0進行數據處理和統計檢驗。
2 結果
2.1 短角異劍水蚤和鹵蟲無節幼體脂肪酸組成分析
短角異劍水蚤與鹵蟲無節幼體脂肪酸組成的分析見表1, 幾種對海水魚類仔稚魚具有重要營養價值的必需脂肪酸(EFA, 即n-3、n-6系列高不飽和脂肪酸 HUFA)中, 濃縮海水小球藻營養強化的鹵蟲無節幼體未檢出 DHA(22: 6n-3), 其 EPA(20: 5n-3)、ARA(20: 4n-6)和EFA也分別僅占其總脂肪酸含量的1.408%、1.090%和3.801%, 而短角異劍水蚤DHA、EPA和ARA含量分別達21.185%、11.088%和3.250%,EFA含量為37.417%, 極顯著高于鹵蟲無節幼體。橈足類DHA/EPA和EPA/ARA分別為1.911和3.412,鹵蟲無節幼體EPA/ARA為1.292。
2.2 短角異劍水蚤和鹵蟲無節幼體投喂對克氏雙鋸魚稚魚成活和生長的影響
5~20日齡, 投喂短角異劍水蚤的克氏雙鋸魚稚魚成活率(80.71%±8.23%)稍高于鹵蟲無節幼體組(76.30%±7.00%)(表2), 但兩者差異不顯著。短角異劍水蚤投喂組克氏雙鋸魚稚魚生長速度顯著高于鹵蟲無節幼體組, 實驗結束時體長極顯著高于后者(P<0.01),體重和特定生長率均顯著高于后者(P<0.05)。
2.3 克氏雙鋸魚稚魚脂肪酸組成的分析
實驗結束時(20 d), 投喂短角異劍水蚤的克氏雙鋸魚稚魚DHA、EPA、ARA和EFA含量均高于鹵蟲無節幼體投喂組(表1), 且2個實驗組的不飽和脂肪酸組成基本與餌料生物相對應。
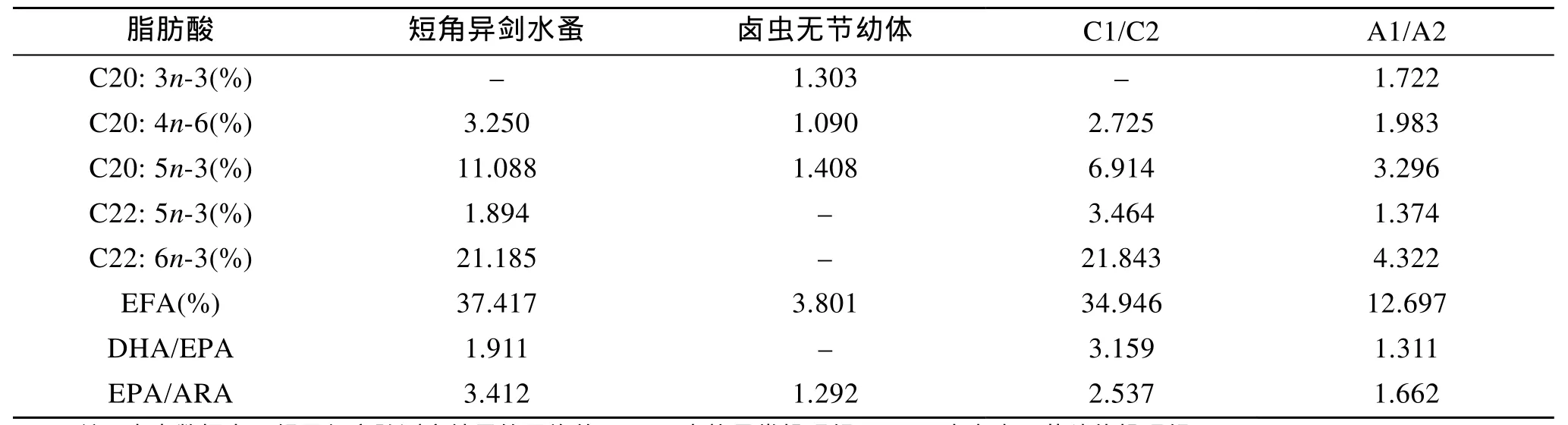
表1 短角異劍水蚤、鹵蟲無節幼體及實驗克氏雙鋸魚稚魚脂肪酸含量的測定Tab.1 Fatty acids composition of Apocyclops royi, Artemia nauplii and juveniles of Amphiprion clarkii

表2 短角異劍水蚤和鹵蟲無節幼體對克氏雙鋸魚稚魚成活和生長的影響Tab.2 The survival and growth rate of Amphiprion clarkii juveniles fed with Apocyclops royi and Artemia nauplii respectively
3 討論
生物餌料的開發和利用是海水魚類苗種人工培育的關鍵技術。長期以來, 海水魚類苗種培育主要依賴于輪蟲和鹵蟲無節幼體作為生物餌料, 在多種魚類苗種人工培育中取得了成效, 但由于二者個體大小范圍有限、營養成分不穩定、缺乏高度不飽和脂肪酸等原因, 并非對所有經濟魚類都適用。橈足類作為海水魚類仔稚魚天然的餌料生物, 將其開發為生物餌料一直是海水魚類苗種培育的研究熱點之一。已有研究證實, 在使用輪蟲作為海水魚類仔魚開口餌料的階段結束之后, 使用橈足類替代鹵蟲無節幼體[2,19], 或是兩者同時投喂都取得了較為滿意的效果[2,20]。本實驗中, 以輪蟲作為克氏雙鋸魚仔魚的開口餌料, 稚魚期單獨投喂短角異劍水蚤的實驗組稚魚的成活率稍高于投喂鹵蟲無節幼體的實驗組, 生長狀態則顯著優于后者, 與上述學者的研究結果相一致, 初步表明短角異劍水蚤作為海水魚類仔稚魚的生物餌料優于鹵蟲無節幼體。本實驗中 2個實驗組克氏雙鋸魚稚魚成活率均高于 75%, 與克氏雙鋸魚苗種培育階段死亡高峰出現在仔魚期(孵化后 2~3日齡)有關[17]。
評價生物餌料優劣的標準除適口性(餌料大小)和運動能力外, 主要考慮其營養價值(如脂肪酸、蛋白質和自由氨基酸), 其中尤以多不飽和脂肪酸(PUFA)含量和組成為主要參數, 特別是 DHA、EPA和 ARA等海水仔稚魚必需的高不飽和脂肪酸(HUFA)[21]。餌料中HUFA的數量和種類, 會直接影響海水仔稚魚的生長速度、成活率、應激能力和體內相關成分的組成[22]。輪蟲和鹵蟲作為傳統生物餌料, 其體內缺乏海水魚類仔稚魚必需的HUFA, 需要富集餌料中 HUFA以滿足海水魚類仔稚魚的營養需求。本實驗中, 經海水小球藻強化的鹵蟲無節幼體仍未檢出 DHA, 小球藻強化對鹵蟲無節幼體必需脂肪酸的組成和含量沒有顯著效果, 與黃旭雄等[23]所得結論一致。而僅使用海水小球藻強化的短角異劍水蚤DHA、EPA、ARA和n-3HUFA含量均較高, 暗示短角異劍水蚤可能具有自身合成 HUFA的能力, 這需要進一步實驗驗證, 但就必需脂肪酸種類和含量而言, 短角異劍水蚤營養價值高于鹵蟲無節幼體。
必需脂肪酸的比例也是衡量生物餌料脂肪酸品質的重要指標, 由于 PUFA在代謝方面存在競爭作用, 主要是DHA和EPA與ARA的競爭, 這3種重要的 HUFA的比例會直接影響海水仔稚魚視網膜和腦的發育[24]。張其永等[25]研究鲆鰈類有眼側色素沉積與 DHA、EPA和 ARA 的比例失調有關, 導致視網膜和腦不能正常發育, 嚴重影響視力; Reitan等[26]用DHA/EPA為2/1的餌料培育大菱鲆仔稚魚, 有效地防止了大菱鲆白化的產生; McEvoy等[27]報道, 對大菱鲆和大西洋庸鰈而言, 餌料中DHA含量占總脂肪酸的6.5%~12.5%, 用EPA/ARA為4.2/1和20/1的乳化油強化活餌料, 仔稚魚表現出正常的生長、成活率、色素和變態, 然而用EPA/ARA為1.5/1和0.5/1的乳化油強化活餌料, 仔稚魚的色素和變態均不正常。此外, 多數海水魚魚卵和仔稚魚體內, DHA/EPA的含量均為2/1左右, 而不同魚類對ARA需求不同,歐洲鱸的 EPA/ARA的適宜值為 1/1, 大菱鲆和大西洋庸鰈的EPA/ARA的適宜比例為10∶1以上。本實驗中短角異劍水蚤DHA/EPA為1.911, EPA/ARA為3.159, 均優于小球藻強化的鹵蟲無節幼體, 從必需脂肪酸比例分析短角異劍水蚤也更適于作為海水魚類仔稚魚的生物餌料加以開發利用。
本文通過分析短角異劍水蚤和鹵蟲無節幼體的脂肪酸組成及2種生物餌料對克氏雙鋸魚稚魚存活、生長和脂肪酸組成的影響表明, 短角異劍水蚤HUFA、DHA、EPA、ARA 含量高, DHA/EPA、EPA/ARA比值與海水魚類仔稚魚必需脂肪酸比例需求類似, 橈足類投喂組表現出較高的生長優勢和成活率, 優于傳統的生物餌料。綜上所述, 短角異劍水蚤適于作為海水魚類仔稚魚的生物餌料, 在苗種培育產業中具有廣闊的應用前景。
[1]Lee C, O'Bryen P J, Marcus N H.Copepods in Aquaculture[M].Ames, Iowa, USA: Blackwell Publishing, 2005.
[2]劉光興, 徐東暉, 邱旭春, 等.火腿許水蚤對牙鲆仔稚魚成活、生長及脂肪酸組成的影響[J].中國海洋大學學報(自然科學版), 2007, 37(2): 259-265.
[3]鄭重, 李少菁, 連光山.海洋橈足類生物學[M].廈門: 廈門大學出版社, 1992.
[4]van der Meeren T.Selective feeding and prediction of food consumption in turbot larvae (Scophthalmus maximusL.) reared on the rotiferBrachionus plicatilisand natural zooplankton[J].Aquaculture, 1991, 93(1):35-55.
[5]Naess T, Germain-Henry M, Naas K E.First feeding of Atlantic halibut (Hippoglossus hippoglossus) using different combinations ofArtemiaand wild zooplankton[J].Aquaculture, 1995, 130(2-3): 235-250.
[6]A review on the status and progress in rearing copepods for marine larviculture: Advantages and disadvantages among calanoid, harpacticoid and cyclopoids copepods[C]//Avances en nutrición acuicola (Vol.Ⅷ).Mazatlán,Sinaloa, México: Memorías del Octavo Simposium Internacional de Nutrición Acuícola, 2006.
[7]St?ttrup J G, Richardson K, Kirkegaard E, et al.The cultivation ofAcartia tonsaDana for use as a live food source for marine fish larvae[J].Aquaculture, 1986,52(2): 87-96.
[8]St?ttrup J G, Norsker N H.Production and use of copepods in marine fish larviculture[J].Aquaculture,1997, 155(1-4): 231-247.
[9]Toledo J D, Golez M S, Ohno A.Use of Copepod Nauplii During Early Feeding Stage of GrouperEpinephelus coioides[J].Fisheries Science, 1999, 65(3):390-397.
[10]Deveney M R, Bayly T J, Johnson C J, et al.A parasite survey of farmed Southern bluetin tuna,Thunnus maccoyii(Castelnau)[J].Journal of Fish Diseases, 2005,28(5): 279-284.
[11]劉俊鵬, 趙文, 王珊, 等.短角異劍水蚤胚胎發育和幼體發育的研究[J].大連水產學院學報, 2007, 22(4):239-245.
[12]Lee K, Dahms H, Park H G, et al.Population growth and productivity of the cyclopoid copepodsParacyclopina nana,Apocyclops royiand the harpacticoid copepodTigriopus japonicusin mono and polyculture conditions: a laboratory study[J].Aquaculture Research, 2012: 1-5.
[13]張文炳.橈腳類短角異劍水蚤之生理生態學研究[D].中國臺北: 臺灣大學, 1992.
[14]許家興.食物及溫度對短角異劍水蚤生長及生殖的影響[D].中國臺灣高雄: 中山大學, 1988.
[15]劉敏, 陳世杰.短角異劍水蚤的培養試驗[J].福建水產, 1995(9): 19-22.
[16]孟振, 王國棟, 劉新富, 等.環境因素對短角異劍水蚤攝食的影響[J].海洋科學, 2013, 37(01): 81-86.
[17]鮑鷹, 張鵬, 祝承勇, 等.紅小丑人工繁殖和育苗的初步研究[J].海洋科學, 2009, 33(2): 5-10.
[18]劉光興, 崔建麗, 黃瑛.幾種單胞藻對火腿許水蚤脂肪酸組成的影響[J].廈門大學學報: 自然科學版,2006, 45(A02): 250-255.
[19]Kraul S, Nelson A, Brittain K, et al.Evaluation of Live Feeds for Larval and Postlarval MahimahiCoryphaena hippurus[J].Journal of the World Aquaculture Society,1992, 4(23): 299-306.
[20]Leu M, Chou Y.Induced spawning and larval rearing of captive yellowfin porgy,Acanthopagrus latus(Houttuyn)[J].Aquaculture, 1996, 143(2): 155-166.
[21]湯保貴, 陳剛, 吳灶和.海水仔魚開口生物餌料的研究進展[J].飼料研究, 2006(4): 43-44.
[22]許友卿, 莊麗, 丁兆坤.多不飽和脂肪酸對海水仔稚魚生長發育的影響及機理[J].飼料工業, 2010, 31(14):13-18.
[23]黃旭雄, 王瑞, 呂耀平, 等.不同強化餌料對鹵蟲必需脂肪酸組成的影響[J].水產科學, 2005, 24(10):1-4.
[24]許友卿, 張海柱, 丁兆坤.二十二碳六烯酸和二十碳五烯酸的代謝研究[J].水產科學, 2007, 26(10):580-583.
[25]張其永, 洪萬樹.海洋養殖魚類仔稚魚攝食和營養研究的進展[J].臺灣海峽, 2001, 20(z1): 1-10.
[26]Reitan K I, Rainuzzo J R, Olsen Y.Influence of lipid composition of live feed on growth, survival and pigmentation of turbot larvae[J].Aquaculture International, 1994, 2(1): 33-48.
[27]McEvoy L A, Estevez A, Bell J G, et al.Influence of dietary levels of eicosapentaenoic and arachidonic acid on the pigmentation success of turbot (Scophtalmus maximus) and halibut (Hippoglossus hippoglossus)[J].Bulletin of the Aquaculture Association of Canada,1998, 98: 17-20.
