安太堡露天礦植被恢復過程中土壤生態肥力評價
2015-12-16 08:15:58李晉川岳建英王宇宏郭春燕楊生權
水土保持研究 2015年1期
李晉川,王 翔,岳建英,王宇宏,郭春燕,盧 寧,楊生權
(山西省生物研究所,太原030006)
露天采礦對土地帶來的直接破壞是一個十分嚴重且日益受到高度重視的問題[1]。平朔露天礦區地處黃土高原東部,屬黃土丘陵強烈侵蝕生態脆弱系統[2]。該區是我國生態最脆弱、水土流失最嚴重的地區之一。在此地區進行大規模的露天開采,更加劇了水土流失。恢復和重建業已退化的生態系統,維持人類生存環境的穩定和持續發展是現代生態學研究的重要課題。礦區土地復墾與生態重建已成為解決人地矛盾、改善生態環境的一項重要措施,同時也是露天采礦企業可持續發展的保障[3]。
目前,在礦山植被恢復方面很多研究取得了較好的經濟與環境效益,如山西省生物研究所黃土高原露天煤礦礦區生態的恢復工藝技術研究、內蒙古農業大學草原露天煤礦礦區的生態恢復和植被恢復工作以及中科院沈陽生態所土壤基質改良等研究[4]。目前國內對于礦區生態恢復的研究工作主要集中在植被的恢復、土壤的 改 良 等 方 面[5-8]。 李 晉 川 等[9]的 研 究表明:在已有二十余年安太堡煤礦廢棄地復墾的植被研究中,選擇豆科牧草作為先鋒植物,以草灌喬復合種植,既可增加植被控制水蝕、風蝕等危害的作用,又同時保證了重建后生態系統的穩定,有利于礦區生產的可持續性發展。也有學者研究認為沙棘、刺槐等植物可以作為排土場的優良生態恢復植被物種,并應結合排土場的特點來選擇復墾最優方式[10]。目前對于不同植被恢復模式對土壤質量影響的研究更多局限于碳氮等理化指標,而對于露天礦區不同植被恢復模式的土壤生態恢復效果進行生物學質量的綜合評價尚無報道。土壤生態肥力是指在一定環境條件下,土壤及其生物群落(包括動物和微生物)之間長期協同進化、相互適應、相互作用而表現出的一種和諧共融特性,以及在該特性狀態下土壤保證植物生長所需物質與能量的可獲得性和可持續性的一種功能與能力[11]。而土壤肥力是土壤質量的重要組成部分,是林地生產力的基礎[12]。因此,本文以安太堡露天礦復墾地不同植被恢復模式為研究對象,對植被恢復過程中土壤碳氮等理化性質、酶活性和微生物多樣性等指標進行測定,分析植被恢復對土壤生態系統影響,為評價植被恢復效果和種植模式篩選提供依據。
1 材料與方法
1.1 研究區概況
平朔礦區地處黃土高原晉陜蒙接壤的黑三角地帶,山西省北部朔州市境內,地理坐標:東經112°11′—113°30′,北緯39°23′—39°37′[13]。平朔礦區屬于典型的溫帶半干旱大陸型季風氣候區,冬春干旱少雨、寒冷、多風,夏秋降水集中、溫涼少風。礦區年平均氣溫4.8~7.8℃,最高溫度為37.9℃,最低溫度為-37.4℃,日溫差為18~25℃。無霜期約115~130d。平朔礦區地帶性土壤為栗鈣土與栗褐土的過渡帶,礦區地帶性植被類型屬于草原,目前總體上呈農業耕作景觀[2]。
1.2 采樣方法
供試土壤采自朔州市平魯區安太堡露天礦復墾地,采樣地分別為刺槐林地(1992年種植,CH)、油松—刺槐—檸條混交林(1992年種植,YCN)和油松—刺槐混交林(1992年種植,YC)。采樣地點具體信息見表1。在距植被50cm的地方按照S型多點采集土樣,取樣深度分別為0—10cm和10—20cm。各樣地內取5個重復,將土壤樣品帶回實驗室,除去石塊與植物殘體,分成兩部分。一部分于4℃下儲存,用于土壤含水量、細菌、真菌、固氮菌、硝化細菌和反硝化細菌數量測定以及細菌、真菌多樣性檢測;另一部分風干后過篩,充分混勻后儲藏,用于土壤pH、有機碳、全氮和堿解氮含量以及蔗糖酶、脲酶、過氧化氫酶和多酚氧化酶活性測定。采樣時間為2010年10月。
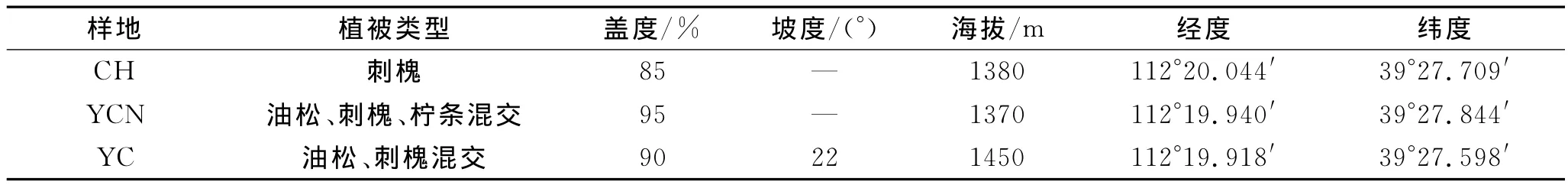
表1 樣地基本特征
1.3 測定項目與方法
土壤酸堿度的測定采用電位法,含水量的測定使用烘干法。土壤有機碳采用重鉻酸鉀氧化法、全氮采用凱氏法、堿解氮采用堿解擴散法[14]。蔗糖酶的測定采用硫代硫酸鈉滴定法,脲酶的測定采用苯酚鈉比色法,過氧化氫酶的測定采用高錳酸鉀滴定法,多酚氧化酶的測定采用鄰苯三酚比色法[15]。土壤微生物主要類群數量的測定采用稀釋涂布平板法[16],其中細菌:牛肉膏蛋白胨培養基;真菌:馬丁氏培養基;自生固氮菌:阿須貝無氮培養基;硝化細菌:硝化細菌培養基;反硝化細菌:反硝化細菌培養基。
使用美國 Mo-Bio的土壤DNA提取試劑盒(power soil DNA kit)提取土壤微生物總 DNA。提取產物用1%瓊脂糖電泳檢測。用Bio-Rad(伯樂)公司的C1000-Touch型PCR儀,采用細菌通用引物(338—GC 5′—CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGG GCCTACGGGAGGCAGCAG—3′,518R5′—ATTACCGCGGCTGGT-GG—3′)擴增土壤中總細菌。采用50μl的反應體系,其中5μl的10×Buffer,4μl的dNTP(2.5mM)、1U Taq酶、引物量各10pmol,DNA模板量10ng。細菌的16SrDNA使用Touchdown法進行擴增,程序是94℃預變性5min;94℃下1min,65℃下1min,72℃3min(退火溫度每循環降0.5℃,共20個循環,降至55℃);94℃下1min,55℃下1min,72℃3min(共10個循環);72℃終延伸10min[17]。
采用真菌18SrDNA通用引物(NS15′—GTAGTCATATGCTTGTCTC —3′GC-Fang 5′—CGCCCGCCGCGCCCCGCGCCCGGCCCGCCGCCC CCGCCCCATTCCCCGTTACCCGTTG—3′)擴增土壤中總真菌。采用50μl的反應體系,其中5μl的10×Buffer,4μl的dNTP(2.5mM)、1U的Taq酶、引物量各10pmol,DNA模板量10ng。真菌18SrDNA的擴增程序是94℃預變性3min;94℃下1min,50℃下1 min,72℃3min(共40個循環);72℃終延伸10min[18]。
變性梯度凝膠電泳使用Bio-Rad基因突變儀,使用10%聚丙烯酰胺凝膠,制膠和電泳緩沖液為1×TAE,濃度梯度為30%~60%(7mol/L尿素和40%去離子甲酰胺為100%變性),電泳電壓120V,時間8h,用SYB greenⅡ(1∶10 000,體積比)染色1h。Bio-Rad公司Versa DOC儀器系統下成像,Quantaty one 4.6.2軟件對DGGE圖譜進行分析。使用Shannon-Weiner指數和Simpson′s diversity指數來表示土壤微生物群落多樣性,計算公式為:
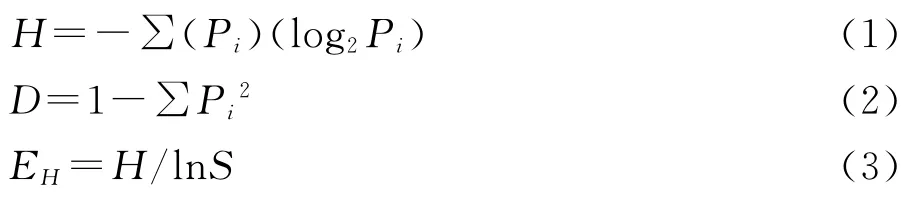
式中:H——香農—威納指數;D——辛普森多樣性指數;EH——均勻度指數;Pi——樣品的多度值;S——每個樣品中的電泳條帶數目。
1.4 土壤生物學肥力的評價方法
應用多元統計分析方法,以各主成分特征值貢獻率為權重,加權計算各樣地土壤生物學肥力指標值,以評價各樣地土壤的生態肥力特征[12,19-20]。
使有機碳、全氮、堿解氮、蔗糖酶、脲酶、過氧化氫酶、多酚氧化酶、細菌、真菌、固氮菌、硝化細菌、反硝化細菌依次為X1—X12,將數據進行標準化處理后,經過主成分分析,得到變量,計算得分系數,進而依據IFI公式得出不同樣地的IFI值。
1.5 數據處理
采用 Excel 2003與SPSS 19.0對數據進行統計分析和計算。
2 結果與分析
2.1 不同植被恢復模式對土壤理化性質的影響
在不同的植被恢復模式下土壤有機碳、全氮、C/N,堿解氮的含量變化見表2,表層土壤中有機碳含量YC最高,且顯著高于CH和YCN。下層土壤中三種模式間差異顯著,其中YC含量最高。而土壤全氮含量和C/N在三種樣地間也表現出相同變化趨勢。與此同時,土壤堿解氮含量變化表現為表層土壤YC和YCN顯著高于CH,但前兩者之間無顯著差異;下層土壤也表現出相同變化趨勢。
2.2 不同植被恢復模式對土壤酶活性的影響
不同的植被恢復模式土壤酶的活性不同(表3)。從表層土壤來看三種恢復模式之間蔗糖酶活性無顯著差異,但在下層土壤中YC的酶活性顯著高于其它模式。脲酶的變化趨勢與蔗糖酶不同,表層土壤中YC的酶活性顯著高于CH和YCN,后二者間無顯著差異;下層土壤表現出相同趨勢。過氧化氫酶活性與土壤微生物的活動有關,在上層土壤中三種恢復模式間差異顯著,其中YC最高,YCN最低;但在下層土壤中三種模式間無顯著差異。多酚氧化酶通常被認為與土壤的腐殖化程度有關,其中在表層土壤YCN顯著低于YC和CH,后二者間無顯著差異;下層土壤也呈相同規律。
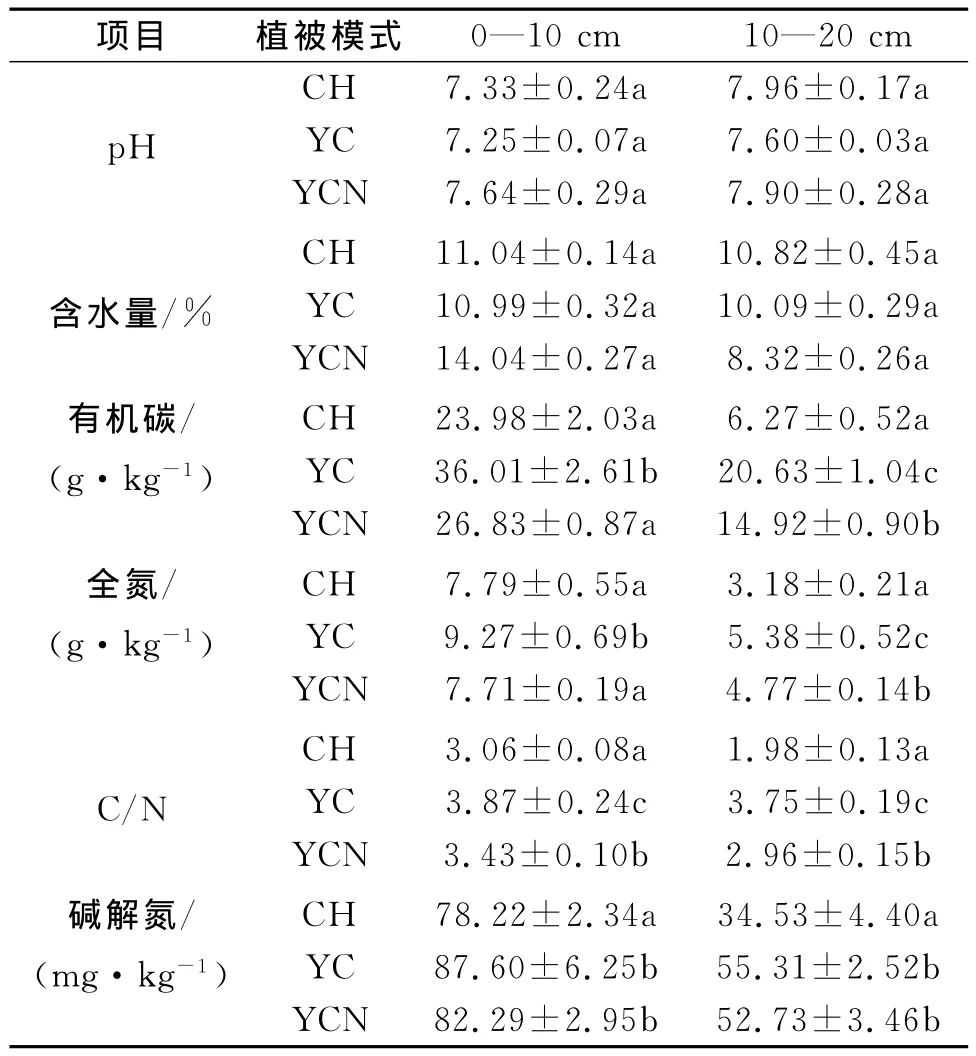
表2 不同樣地的土壤理化性質
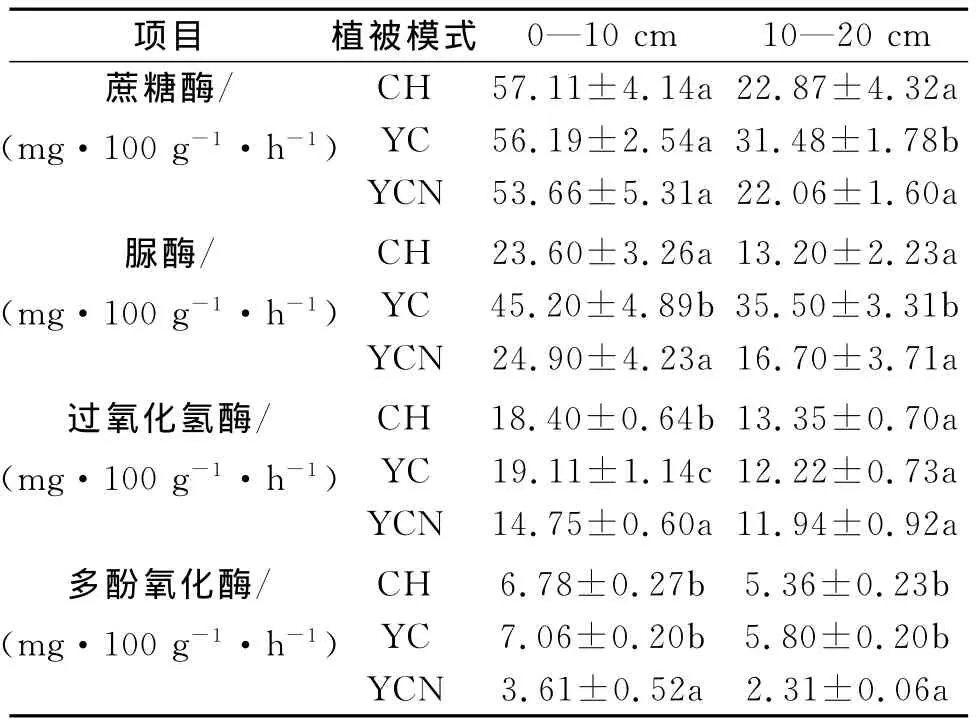
表3 不同樣地的土壤酶活性
2.3 不同植被恢復模式對土壤微生物數量的影響
對安太堡露天礦不同植被恢復模式土壤微生物數量分析表明,表層土壤中YC的細菌數量顯著高于CH和YCN,后二者間無顯著差異;在下層土壤中三種模式差異顯著,其中YC最高,CH最低。真菌數量表現出不同的變化規律,上層土壤YCN顯著高于其它模式,下層土壤不同樣地間均無顯著差異。固氮菌和反硝化細菌在上層土壤中變化規律相同,不同植被模式間均無顯著差異;在下層土壤中固氮菌數量表現為CH顯著低于YC和YCN,反硝化細菌數量則是YC顯著高于CH和YCN。同時,CH和YCN的硝化細菌數量顯著高于YC且二者間無顯著差異,下層土壤則均無顯著差異(表4)。

表4 不同樣地的土壤微生物數量
2.4 不同植被恢復模式對土壤微生物群落多樣性的影響
16SrDNA擴增的DGGE條帶的圖像分析表明(圖1A),在3個樣地中共有5個優勢細菌DNA條帶,條帶的數量在5~7之間變化,說明不同的植被恢復模式影響細菌的多樣性和結構。在CH中條帶數最少,只有5條,分別是A、B、C、D、E,并且這五個條帶在所有樣地中均有出現,說明這些細菌為該類型土壤中常見類型,并且廣泛分布。可以看到YCN的條帶數增加到了6條,即具有新的條帶F。YC的條帶數為7條,即增加了G、H,可見細菌的群落多樣性顯著不同。
18SrDNA擴增的DGGE條帶圖像分析表明(圖2B),在3個樣地中共有6個優勢DNA條帶,條帶數量在7~9之間變化,反映出不同的植被恢復模式影響真菌的群落多樣性。在CH中,條帶數最少,只有7條,分別是A、B、C、D、E、F、K,并且除K之外,其余條帶在所有樣地均有出現,說明這些真菌為該類型土壤中常見種類。還可以看到YC條帶數增加到了9條,即具有新的條帶G、L、M、YCN的條帶數為9條,增加了新的條帶H、I,可見植被恢復對真菌群落多樣性影響不同。
同時,不同植被恢復模式土壤中細菌、真菌的群落多樣性通過多樣性指數反映出來,由表5可見,細菌的群落多樣性從香農指數和辛普森指數來看均表現為YC最高,真菌的群落多樣性從香農指數和辛普森指數來看均表現為YCN最高。此外,三種樣地的細菌、真菌群落多樣性均勻度指數較為接近。
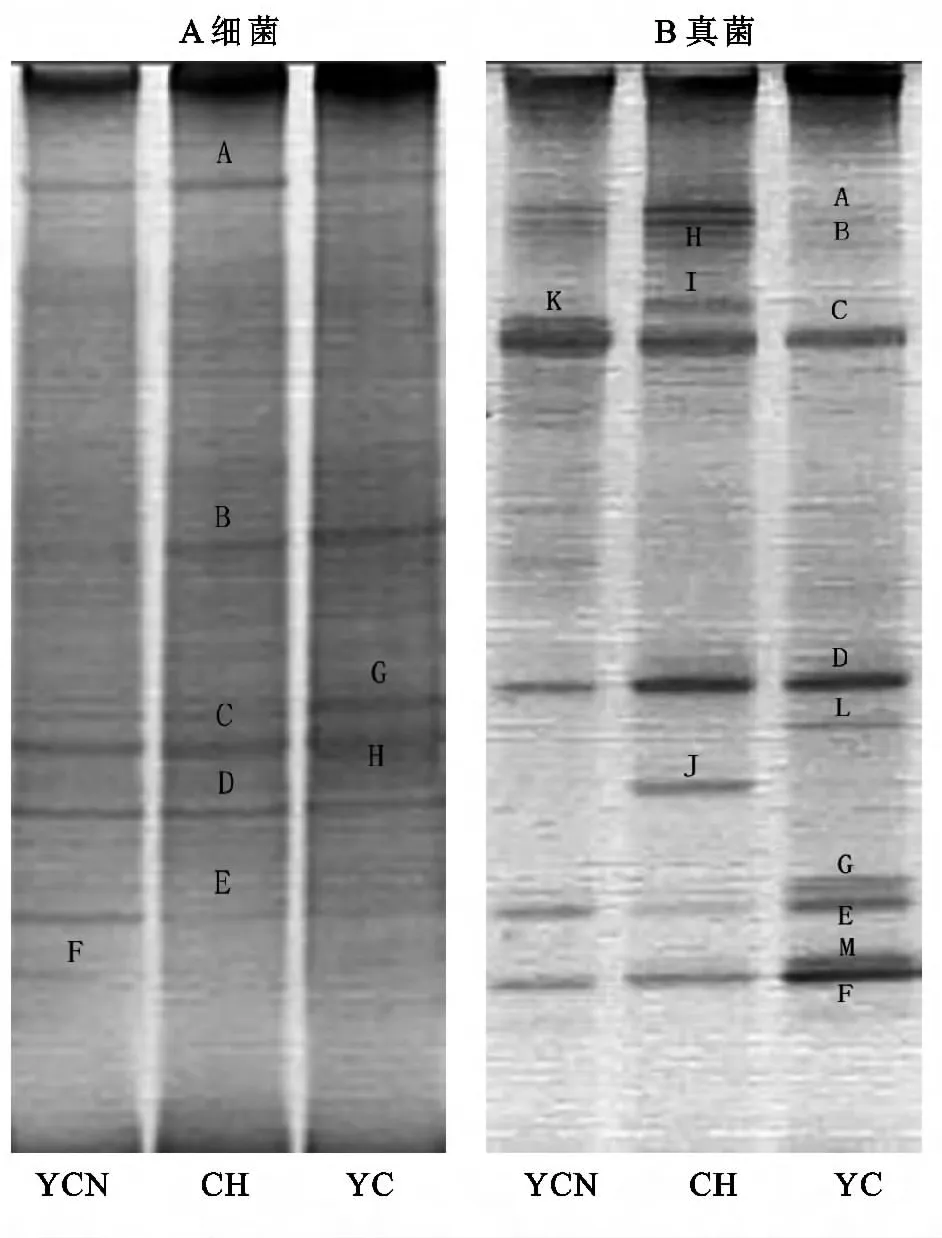
圖1 不同樣地土壤細菌、真菌的DGGE圖譜
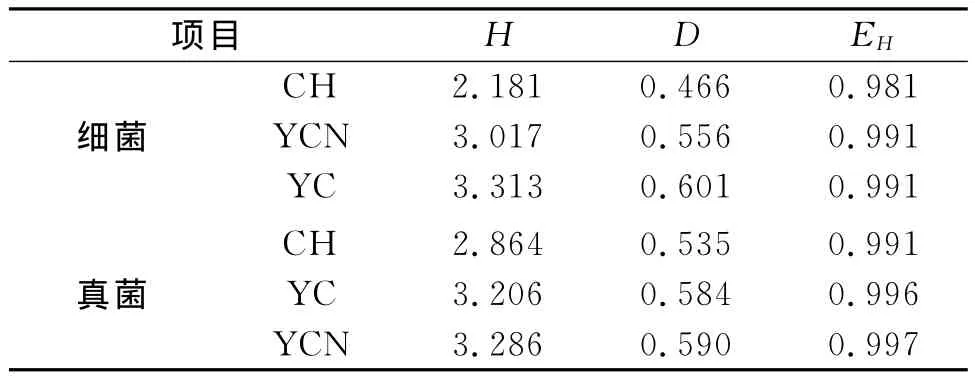
表5 不同樣地土壤細菌、真菌的多樣性指數
2.5 不同植被恢復模式土壤生物學肥力評價
為了全面反映土壤的生物學活性,需要進一步分析土壤各個生物學指標與土壤生物學肥力的關系,在前人研究方法基礎上,使用主成分的分析方法評價各植被模式下的土壤生物學肥力高低。使用X1,X2…X12分別代表有機碳、全氮、堿解氮、蔗糖酶、脲酶、過氧化氫酶、多酚氧化酶、細菌、真菌、固氮菌、硝化細菌、反硝化細菌。
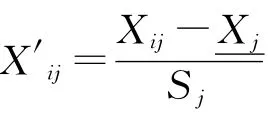
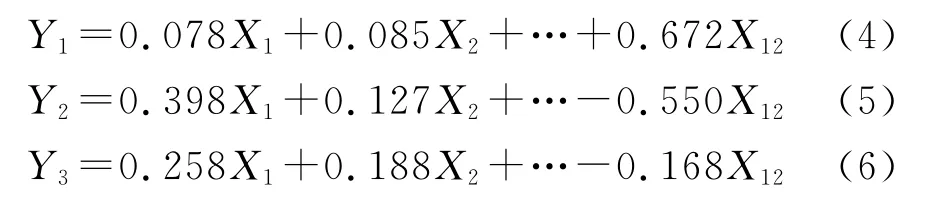
各主成分的因子得分見表6。可以看出,第一主成分Y1主要反映了有機碳、全氮、堿解氮、蔗糖酶、細菌、真菌、固氮菌、硝化細菌、反硝化細菌等指標的綜合變量,對土壤生物學肥力評價十分重要。第二主成分Y2主要反映了蔗糖酶、脲酶、過氧化氫酶、多酚氧化酶等指標的綜合變量,對土壤生物學肥力評價具有一定作用。第三主成分Y3主要反映了固氮菌、堿解氮、硝化細菌等指標的綜合變量,對土壤生物學肥力評價具有一定作用,但影響較小。
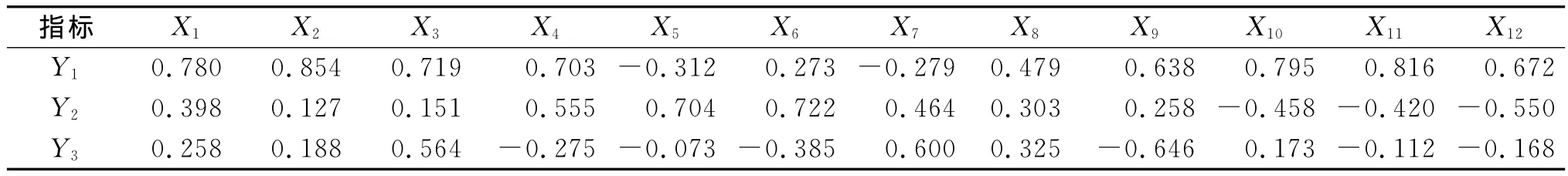
表6 土壤生物學指標主成分因子得分系數矩陣
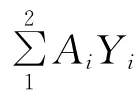
由表7可見,不同植被恢復模式土壤生物學指標綜合評價得分值(IFI)不同。依據其得分多少進行排序,可知表層土壤YC>CH>YCN;下層土壤YC>YCN>CH。可以看到,不同植被恢復模式的土壤生物學性狀具有明顯差異,并且表現出表層土壤高于下層土壤的趨勢。其中YC土壤生物學性狀最優。
2.6 土壤理化性質、土壤酶活性與土壤微生物數量的相關性
通過對復墾地土壤理化性質、土壤酶活性和土壤微生物數量相關性分析表明,三類指標均存在相關關系,但相關程度有所不同(表8)。pH與蔗糖酶顯著負相關;有機碳與全氮、C/N、堿解氮極顯著正相關,與蔗糖酶顯著正相關,與脲酶、多酚氧化酶極顯著負相關;全氮與C/N、堿解氮、脲酶、真菌極顯著正相關,與蔗糖酶、自生固氮菌顯著正相關,與多酚氧化酶極顯著負相關;C/N與堿解氮極顯著正相關,與蔗糖酶顯著正相關,與脲酶、多酚氧化酶極顯著負相關;堿解氮與多酚氧化酶極顯著負相關,與脲酶、過氧化氫酶、反硝化細菌顯著負相關;蔗糖酶與細菌、硝化細菌極顯著正相關,與真菌、自生固氮菌顯著正相關,與多酚氧化酶極顯著負相關,與脲酶顯著負相關;脲酶與多酚氧化酶極顯著正相關;過氧化氫酶與反硝化細菌極顯著正相關,與真菌顯著正相關;多酚氧化酶與細菌極顯著負相關,與真菌、自生固氮菌顯著負相關;細菌與真菌、自生固氮菌、硝化細菌、反硝化細菌極顯著正相關;真菌與自生固氮菌、硝化細菌、反硝化細菌極顯著正相關;自生固氮菌與硝化細菌、反硝化細菌極顯著正相關;硝化細菌與反硝化細菌極顯著正相關。土壤理化性質與酶活性相關性較強,可能是土壤酶參與促進土壤物質代謝與轉化,而土壤酶與微生物相關密切,可能與土壤酶大部分來源于微生物有關。
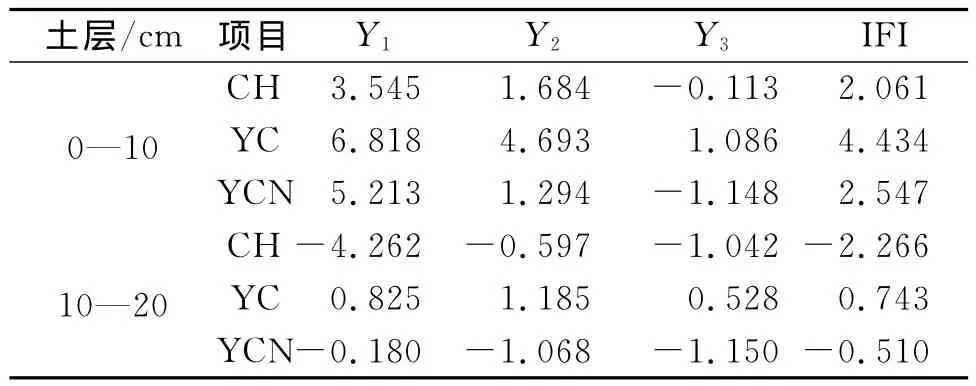
表7 各樣地土壤肥力綜合指標值
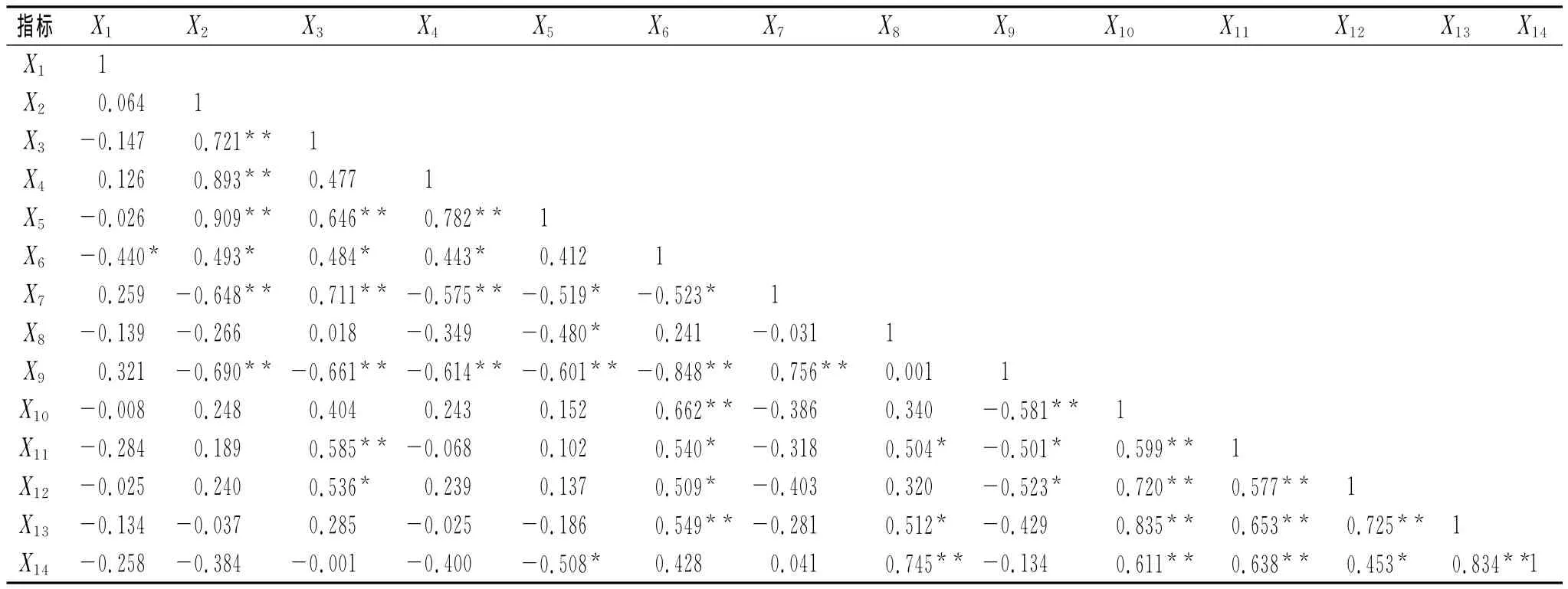
表8 土壤理化性質、酶活性與微生物數量的相關性
3 討 論
(1)安太堡露天礦復墾區完全是由人工建成、土壤結構破壞極其劇烈、植被恢復從零開始的生態修復區,如何對復墾區植被恢復模式進行科學、合理的篩選搭配是在安太堡露天礦進行植被恢復所面臨的主要問題之一。本研究結果表明,對不同植被模式生態肥力的比較呈現YC>CH>YCN,說明油松—刺槐混交林的恢復效果較好,這也與本課題組進行生態調查時所觀測到的結果一致,也說明了檢測指標的選取與不同植被實際生長情況具有一致性。
(2)通過主成分分析,得到對安太堡露天礦區復墾地土壤生態肥力綜合評價起主要作用的若干指標,如有機碳、全氮、堿解氮、蔗糖酶、細菌數量、真菌數量、固氮菌數量等。通過比較這些指標的優劣,可為安太堡露天礦區植被恢復效果評價及植被模式篩選提供科學依據。
(3)在安太堡露天礦復墾地三種植被模式中均表現出碳氮含量隨土層加深而遞減,表現出植被恢復下土壤有機質和氮素具有一定表聚性。植被產生的凋落物為土壤提供了有機物質原料,促進了土壤物質循環,提高了土壤肥力,進而促進了植物生長發育,形成良性循環,逐步提高了土壤質量[21]。
(4)同時,土壤酶活性也表現出相同的變化趨勢。安韶山等[22]認為大多數植被恢復模式對土壤的脲酶活性有著促進作用。安太堡露天礦進行植被恢復后,土壤蔗糖酶和脲酶活性的增強,表明土壤中碳素和氮素循環轉化強度有較大的提高。此外,本研究的結果也說明土壤酶活性與土壤生物學肥力有關并且可以作為衡量土壤生物學性狀高低的指標之一[23]。
(5)土壤微生物數量因植被模式和土壤層次不同而存在顯著差異,這可能是由于不同植被模式植物根系生長和互作不同所致,Bosalo等[24]根據擴增的16SrDNA的DGGE指紋圖譜分析,發現微生物群落結構的不同與不同的植被模式有關。植被生長使得土壤微生物種類趨于豐富,群落多樣性更高[25],而本研究結果也證實了這一點。不同的恢復模式形成不同的微生態環境,為土壤微生物生長提供不同的食物來源與生長條件,導致了土壤微生物群落結構的多樣化。隨著植被生長演替和種植時間增加,植被對土壤的改造也趨于增強[26],并促進土壤理化性質良性發展,提高了土壤酶活性和微生物數量,影響了土壤微生物群落多樣性,改善了土壤的生物學性狀。
(6)本研究結果說明,土壤理化性質與土壤的酶活性和微生物數量具有不同程度的相關,說明土壤理化性質、酶活性和微生物數量之間相互影響。不過三類指標的相關性會因為植被、土壤、大氣、地理、環境綜合影響而出現不同狀況。
4 結 論
植被恢復對于安太堡露天礦區復墾地土壤質量改善具有重要影響,不同的植被模式影響效果不同。通過比較不同樣地土壤理化性質指標,可見油松—刺槐混交林的相關數值較好;同時,對土壤酶活性進行綜合比較,可知油松—刺槐混交林的相關指標較好;此外,不同的土壤微生物類群也表現出這一趨勢,土壤細菌、真菌群落多樣性也呈現相同變化規律。從土壤生物學指標綜合評價得分值來看,油松—刺槐混交林恢復效果最好,土壤有機碳、全氮、堿解氮、蔗糖酶、細菌數量、真菌數量、固氮菌數量等可作為植被模式篩選與土壤生態肥力評價的科學指標。
[1] 楊修,高林.德興銅礦礦山廢棄地植被恢復與重建研究[J].生態學報,2001,21(11):1932-1940.
[2] 曹銀貴,白中科,劉澤民,等.安太堡露天礦區土地類型變化研究[J].西北林學院學報,2007,22(2):44-48.
[3] 畢如田,白中科,李華,等.基于RS和GIS技術的露天礦區土地利用變化分析[J].農業工程學報,2008,24(12):201-204.
[4] 黃丹勇.礦區土地復墾與生態環境恢復綜述[J].湖南有色金屬,2011,27(6):45-48.
[5] 臺培東,李培軍,賈宏宇,等.排土場邊坡人工沙棘灌叢對風滾植物的固留作用及生態效應[J].應用生態學報,2001,12(6):833-836.
[6] 王改玲,王小利,李東方,等.安太堡露天煤礦復墾地土壤養分相關研究[J].煤礦環境保護,2001,15(5):25-27.
[7] 魏忠義,馬銳,白中科,等.露天礦大型排土場水蝕特征及其植被控制效果研究:以安太堡露天煤礦南排土場為例[J].水土保持學報,2004,18(1):164-167.
[8] 鐘爽,劉志斌.生態學在露天煤礦排土場生態恢復中的應用[J].露天采礦技術,2006(2):51-53,55.
[9] 李晉川,王文英.安太堡露天煤礦新墾土地植被恢復的探討[J].河南科學,1999(增刊):92-95.
[10] 臺培東,孫鐵珩,賈宏宇,等.草原地區露天礦排土場土地復墾技術研究[J].水土保持學報,2002,16(3):90-93.
[11] 章家恩,廖宗文.試論土壤的生態肥力及其培育[J].土壤與環境,2000,9(3):253-256.
[12] 魏媛,喻理飛,張金池,等.退化喀斯特植被恢復過程中土壤生態肥力質量評價:以貴州花江喀斯特峽谷地區為例[J].中國巖溶,2009,28(1):61-67.
[13] 薛建春,蔡松.生態脆弱礦區土地利用動態變化研究:以平朔礦區為例[J].水土保持研究,2011,18(6):204-207.
[14] Kelly J J,Tate R L.Effects of heavy metal contamination and remediation on soil microbial communities in the vicinity of azincsmelter[J].Journal of Environmental Quality,1998,27(3):609-617.
[15] 關松萌.土壤酶及其研究法[M].北京:農業出版社,1986.274-339.
[16] 中國科學院南京土壤研究所.土壤微生物試驗法[M].北京:科學出版社,1983.
[17] 王奇贊,徐秋芳,姜培坤,等.天目山毛竹入侵闊葉林后土壤細菌群落16SrDNA V3區片段PCR的DGGE分析[J].土壤學報,2009,46(4):662-669.
[18] Lisa A M,Brenda S,Michael G S.Comparative denaturing gradient gel electrophoresis analysis of fungal communities associated with whole plant corn silage[J].Journal of Microbial,2001,47(9):829-841.
[19] 張慶費,宋永昌,由文輝.浙江天童植物群落次生演替與土壤肥力的關系[J].生態學報,1999,19(2):174-178.
[20] 駱伯勝,鐘繼洪,陳俊堅.土壤肥力數值化綜合評價研究[J].土壤,2004,36(1):104-106.
[21] 黃耀,沈雨,周密,等.木質素和氮含量對植物殘體分解的影響[J].植物生態學報,2003,27(2):183-188.
[22] 安韶山,黃懿梅,鄭粉莉.黃土丘陵區草地土壤脲酶活性特征及其與土壤性質的關系[J].草地學報,2005,13(3):233-237.
[23] 張焱華,吳敏,何鵬,等.土壤酶活性與土壤肥力關系的研究進展[J].安徽農業科學,2007,35(34):11139-11142.
[24] Bosalo D A,Girvan M S,Verchot L.Soil microbial community response to land use change in an agricultural landscape of western Kenya[J].Micorbial Ecology,2005,49(1):50-62.
[25] 夏北成,Zhou J,Tiedje J M.土壤細菌類克隆群落及其結構的生態學特征[J].生態學報,2001,21(4):574-578.
[26] 周濤,戴全厚,吳秀芹,等.喀斯特山區退耕還林地土壤養分效應及評價[J].水土保持研究,2011,18(6):71-74.
