鈾和重金屬復合污染對菊苣葉綠素熒光特性的影響
2015-12-22 05:22:12李華麗唐永金
湖南師范大學自然科學學報 2015年2期
李華麗,唐永金,b*,曾 峰
(西南科技大學a.生命科學與工程學院,b.核廢物與環境安全國防重點學科實驗室,中國綿陽 621010)
隨著核燃料與核工業技術的發展,核素在開采和利用過程中對環境造成的污染問題日益引起人們的關注.鈾是目前最主要的核原料,在自然條件下存在3 種同位素:234U,235U 和238U[1-3].鈾既是一種核素,同時也是一種重金屬,其毒性效應源于它的化學性質和放射性質,毒性的大小與鈾化合物的溶解性密切相關,溶解度越高,毒性越大[4].鈾的溶解態通常以鈾酰離子([UO2]2+)的形式存在.環境中的鈾污染主要來自于反應堆、核武器的研究試驗、核燃料產品、鈾礦山“三廢”排放、事故性核泄漏等[5].鈾礦往往伴生有鐵礦物、鈣鎂礦物、硅酸鹽、鋁硅酸鹽及錳化合物等,因此,在鈾污染的地區其他重金屬的含量也很高,如Fe,Mn,Mg,Ca,Cu 和Zn 等[6].被污染環境中的重金屬可以通過食物鏈等多種途徑進入人體,對人類帶來很大的危害,因此,開展鈾和其他重金屬的生物效應及其污染環境的生物修復方面的研究具有非常重要的意義.
目前,富集植物的篩選仍然是國內外有關鈾和重金屬污染生物修復研究的重要內容,已經發現美洲商陸(Phytolacca americana L.)可以富集Mn[7];羽葉鬼針草(Bidens maximowicziana Oett)可以富集Pb[8],并且對Pb 的耐受性強;鳳眼蓮(Eichhornia crassipes)對Cd2+,Pb2+有很高的去除率[9];向日葵(Helianthus annuus)和菊苣(Cichorium intybus L.)對鈾污染土壤具有潛在的修復能力[10-11];苧麻(Boehmeria nivea)對土壤Hg 有一定的吸收能力[12];蜈蚣草(Pteris vittata L.)是一種As 的超積累植物[13].此外,鈾和其他重金屬對植物種子萌發、抗氧化酶體系、光合作用的影響也有一些報道[14-16],但目前關于鈾和其他重金屬復合污染對植物光合作用的影響研究卻鮮有報道,大多集中于單一元素污染的影響.本實驗以菊苣(Cichorium intybus L.)為實驗材料,采用葉綠素熒光技術,研究鈾與Cd2+,Pb2+,Hg2+和Ni2+復合污染對菊苣葉片光系統活性的影響,為鈾和重金屬復合污染影響植物生長的生理機制提供一定的理論依據.
1 材料和方法
1.1 試驗材料
供試植物為菊苣(Cichorium intybus L.),種子由武漢慶發禾盛種業有限責任公司生產.唐永金等(2013)對14 種植物研究發現,在500 μg·g-1鈾污染的土壤中菊苣對鈾的富集量最高,達到728.7 μg·g-1DW[10].
供試土壤為壤土,土壤中鈾和5 種重金屬的本底值分別為鈾2.26 μg·g-1,鎘4.449 μg·g-1,鉛48.83 μg·g-1,汞0.117 μg·g-1,鎳103.79 μg·g-1,鉻149.27 μg·g-1.
供試藥劑為:UO2(CH3COO)2·2H2O,Cd(CH3COO)2·2H2O,Pb(CH3COO)2·3H2O,Ni(CH3COO)2·4H2O,(CH3COO)2Hg,Cr(CH3COO)3·6H2O,其中鈾的同位素為238U.
1.2 試驗方法
盆栽試驗,每盆2 kg 干土.鈾處理量分別為0,50,100,150 μg·g-1干土,每個鈾處理量下設計5 個重金屬處理,各重金屬的處理分為0、低含量、高含量.低含量大約為土壤背景值的1 倍(汞除外),高含量大約為土壤背景值的5 倍(表1).以鈾和重金屬的0 處理作為對照.3 次重復.試驗在西南科技大學核素生物效應試驗場通風透光的塑料避雨棚中進行.

表1 重金屬處理設置(單位:μg·g -1)Tab.1 Concentration of heavy metals used in this test(unit:μg·g -1)
按照土壤最大持水量法[15],先將鈾和重金屬配制成母液,根據各處理組合混合在一起,然后用自來水定容至720 mL(預備試驗確定的土壤最大持水量)澆入土壤中,保證試驗土壤被藥液均勻污染.土壤藥劑處理在2012年12月28日進行,在陰涼干燥處放置兩個月以上,使土壤吸附固定,代表污染土壤[17].2013年3月18日播種,出苗后2 周每盆定苗9 株.每1~2 天澆一次水,保持土壤濕度在田間持水量的60%~70%.5月21日開始測定葉綠素熒光.
1.3 測定方法
葉綠素熒光參數的測定采用便攜式脈沖調制葉綠素熒光儀(PAM-2500,Walz).測定前暗適應30 min,每盆測定植株上部最長最大的葉片.
(1)熒光誘導曲線參數的測定 打開測量光測定初始熒光(F0),再執行一個飽和脈沖光測定最大熒光(Fm),然后打開光化光(光強為300 μmol·m-2·s-1)持續5 min,并隔20 s 執行一次飽和脈沖.PSⅡ的最大光合量子產量(Fv/Fm)、PSⅡ的實際光合量子產量(Yield,即Y(Ⅱ))、通過PSⅡ的電子傳遞速率(ETR)、光化學猝滅系數(qP)、非光化學猝滅系數(qN)等熒光參數通過PAM-WIN 軟件(Walz)獲得.
(2)光響應曲線參數的測定 光響應曲線參數的測定參考White(1999)[18]和陳梅(2013)[19]的方法.光合有效輻射(PAR)梯度設定為0,5,24,110,222,413,713,1 287,1 842,2 593 和3 768 μmol·m-2·s-1,每個光強持續時間為20 s.測定完光誘導曲線后,將熒光儀調到光響應曲線測定模式,獲得表觀量子效率(α)、最大電子傳遞速率(ETRmax)和半飽和光強(Ik)等參數.
1.4 數據處理
測得的數據用DPS 9.5 進行方差分析和Duncan 新復極差法多重比較.因Cr 處理的土壤中植物大部分死亡,故不在此進行分析.
2 結果與分析
2.1 鈾和重金屬復合污染對菊苣F0,Fm 和Fv/Fm 的影響
F0,Fm和Fv/Fm是植物葉片葉綠素熒光特性的重要參數.F0可以反映逆境脅迫對植物葉片PSⅡ永久性傷害的程度,Fm反映了通過PSⅡ的最大電子傳遞潛力,Fv/Fm可以反映出完整植物葉片的光合器官的生理狀況.
由表2 可知,在鈾單獨作用下,隨鈾含量的增加,F0逐漸升高,Fv/Fm則有一定程度的下降,當鈾為150 μg·g-1時,F0比0 處理增加了18.98%(P <0.05),Fv/Fm比CK(鈾和重金屬均為0 處理)降低了6.05%(P<0.05).在重金屬單獨作用下(鈾處理為0 μg·g-1),F0隨Cd2+和Ni2+處理量的增大而升高,隨Pb2+和Hg2+處理量的增加而降低,但是均高于對照值0.374;當Cd2+為25 μg·g-1時比對照增加了21.66%,增加顯著(P <0.05).Fm隨著金屬含量增加而減小,并且低含量處理下均大于對照值1.960,高含量處理中除Pb2+處理外都小于對照.Fv/Fm隨著Cd2+含量升高而顯著降低(P <0.05),隨Pb2+含量升高而略有增加.
在鈾和4 種重金屬分別共存的條件下,w(238U)≤100 μg·g-1時,F0值高于單獨的鈾處理,并且在同一鈾含量下,隨著Cd2+含量增加而增加,當Cd2+為25 μg·g-1時達到最大值,顯著(P <0.05)高于單獨的鈾處理(w(238U)=50 μg·g-1.),增加了21.87%.在同一鈾含量處理下,Cd2+含量增加時,Fm和Fv/Fm減小,當w(Cd2+)=25 μg·g-1時小于單獨的鈾處理,當w(Pb2+)=50 μg·g-1及w(Ni2+)=100 μg·g-1時Fm大于單獨的鈾處理,Fv/Fm變化較小.當w(238U)=150 μg·g-1時,各重金屬使F0均低于單獨的鈾處理,當w(Cd2+)=25 μg·g-1和w(Hg2+)=5 μg·g-1時F0分別比單獨的鈾處理降低了14.38%和13.9%,相反的,Fv/Fm均高于單獨的鈾處理.
Fv/Fm是評價植物遭受脅迫的最重要指標.在低鈾含量下,鎘、鉛、汞離子的存在,降低了植物的Fv/Fm值,增加了植物的脅迫,但鎳離子的存在幾乎不產生影響;在高鈾含量下,重金屬離子的存在,提高了Fv/Fm值,降低了鈾離子單獨的脅迫.
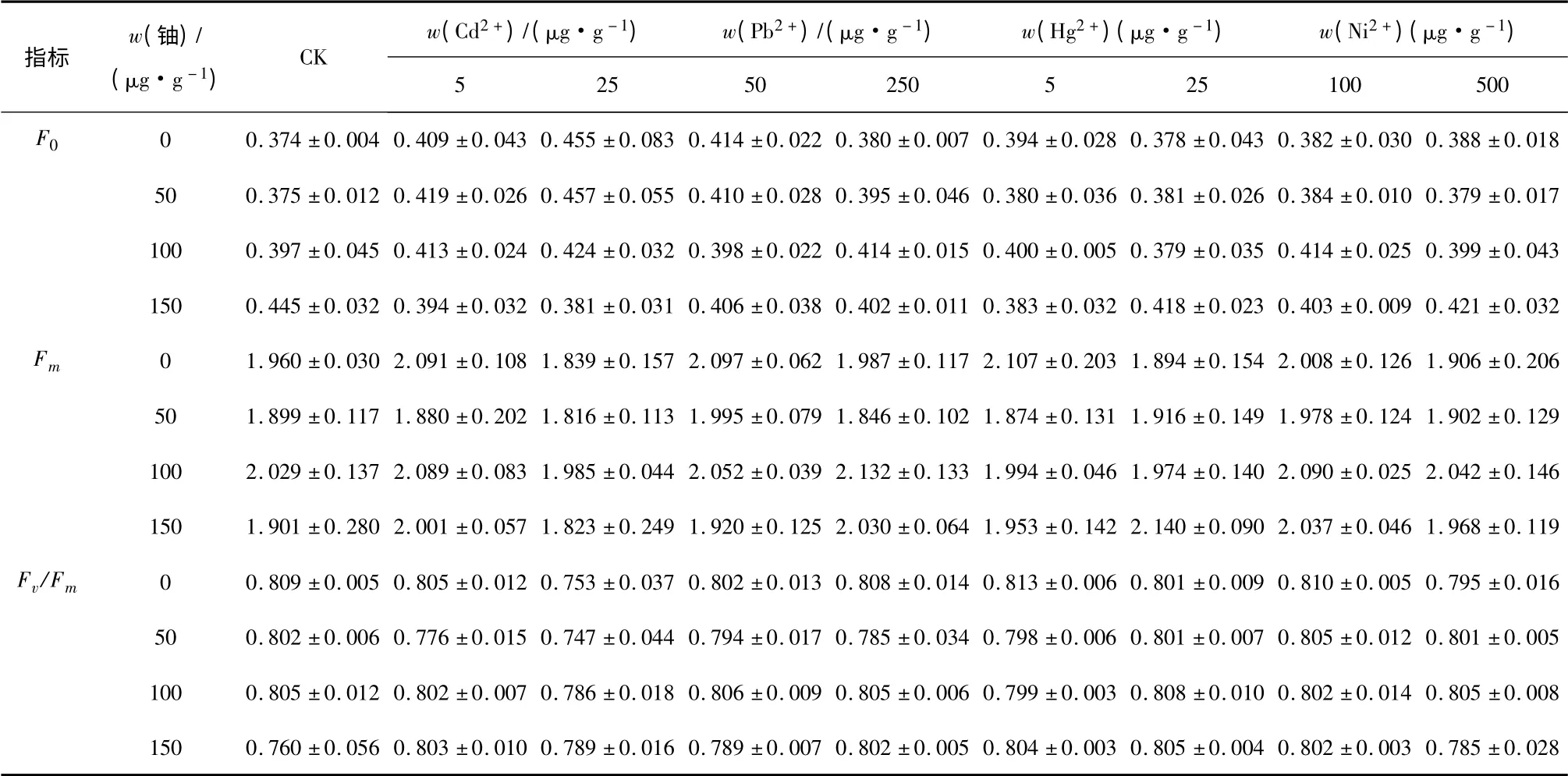
表2 鈾和重金屬復合污染下菊苣葉片F0、Fm、及Fv/Fm 的變化(平均值±標準差,n=3)Tab.2 Changes in the minimal fluorescence yield(F0),the maximum fluorescence yield(Fm),and Fv/Fm of the leaves of Cichorium intybus L.exposed to multiple contamination of uranium and heavy metals.Values are means ± SD(standard deviation)of three measurements
2.2 鈾和重金屬復合污染對菊苣Yield,ETR,qP 和qN 的影響
PSⅡ的實際光合量子產量(Yield)反映了PSⅡ反應中心有部分關閉情況下的原初光能捕獲效率.由表3可知,在鈾單獨作用下,Yield 值隨鈾濃度增加先升高后降低,當w(238U)=50 μg·g-1時比0 處理升高了5.73%.在重金屬單獨作用下,Pb2+,Hg2+及Cd2+低含量使Yield 值高于對照值0.593,Ni2+沒有影響.鈾和重金屬共存時,在同一鈾含量處理下,Yield 值隨Cd2+,Pb2+和Ni2+含量的升高而降低,在Cd2+,Pb2+的高含量及Ni2+處理下小于單獨鈾處理.
PSⅡ的電子傳遞速率(ETR)反映PSⅡ反應中心在光適應條件下的實際電子傳遞情況.光化學淬滅系數(qP)表明植物吸收的光能用于光化學反應部分的大小,qP增加反映PSⅡ的電子傳遞活性升高.由表3 可知,在不同的處理組合中(高含量Cd2+除外)ETR 和qP均大于CK,說明向土壤中添加鈾和重金屬離子有助于菊苣葉片PSⅡ反應中心的電子傳遞.在鈾和重金屬單獨作用下,隨著鈾和重金屬含量的升高,ETR 和qP表現出增加的趨勢.當w(238U)≥100 μg·g-1時,鈾和高含量的Cd2+,Pb2+及Hg2+,Ni2+共存的條件下,ETR 值小于單獨鈾處理,但是高于單獨的重金屬處理,qP小于單獨的鈾及重金屬處理.
非光化學淬滅系數(qN)反映PSⅡ天線色素吸收的部分光能沒有用于光合電子傳遞而以熱形式耗散掉了,是植物的一種自我保護機制,能對光合機構起到一定的保護作用.從表3 中的數據可見,在單獨的鈾作用下,當w(238U)=100 μg·g-1時qN達到最大值,比CK 增加了20.86%,當w(238U)=150 μg·g-1時則比0 處理降低了11.14%.在重金屬單獨作用下,qN隨Cd2+含量增加而增加;隨Pb2+和Ni2+含量的增加而略有降低,但仍然高于0 處理.鈾和Cd2+,Ni2+共存時,qN隨鈾和Cd2+,Ni2+含量的增加而升高,當w(238U)=150 μg·g-1和w(Cd2+)=25 μg·g-1,w(Ni2+)=500 μg·g-1時達到最大值,分別比對照增加了45.02%和81.67%(P <0.01).在鈾和Pb2+共存的處理中,當w(238U)=50 μg·g-1時qN隨Pb2+含量增加而降低,當w(238U)≥100 μg·g-1時則表現出相反的變化趨勢;含Hg2+的處理與Pb2+處理的變化趨勢相反.
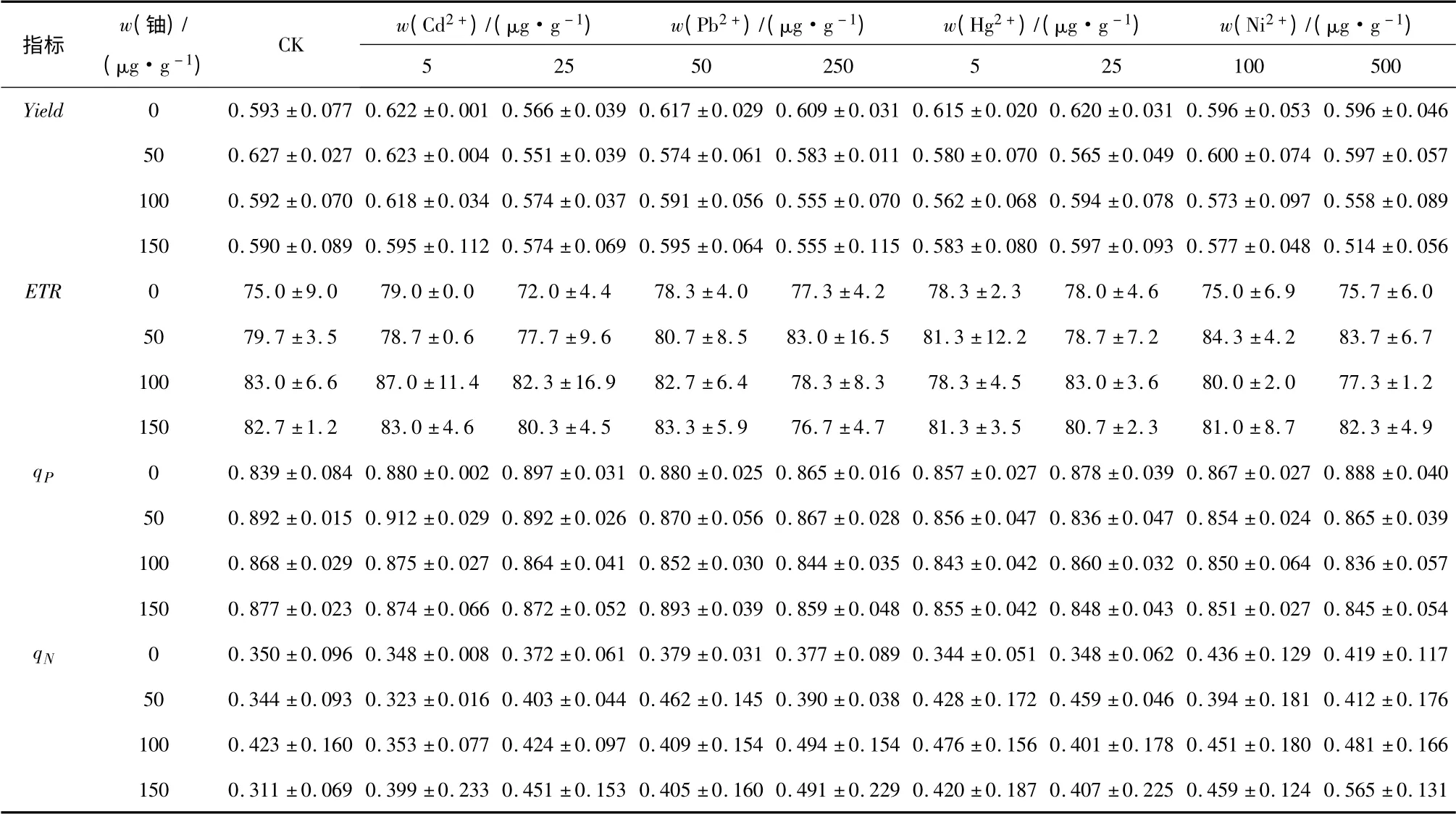
表3 鈾和重金屬復合污染下菊苣葉片Yield、ETR、qP 及qN 參數的變化(平均值±標準差,n=3)Tab.3 Changes in the actual photochemical efficiency of PSII(Yield),the photosynthetic electron transport(ETR),the photochemical quenching coefficient(qP)and the non-photochemical quenching coefficient(qN)of the leaves of Cichorium intybus L.exposed to multiple contamination of uranium and heavy metals.Values are means ± SD(standard deviation)of three measurements
實驗結果表明,同一鈾含量處理下,重金屬離子的存在提高了植物葉片的qN,植物的自我保護機制增強;同時高含量重金屬離子的存在降低了Yield、ETR 和qP,植物天線色素的捕光功能下降,電子傳遞鏈受阻.
2.3 鈾和重金屬復合污染對菊苣葉片快速光響應曲線參數的影響
表觀量子效率(α)表示光響應曲線的初始斜率,反映了葉片對光能,尤其是對弱光的利用效率.最大電子傳遞速率(rETRmax)是不存在光抑制時的最大光合速率,即潛在的最大相對電子傳遞速率,代表葉片光合作用的最大能力.半飽和光強(Ik)可以反映葉片對強光的耐受能力.在鈾單獨作用下,α,rETRmax和Ik變化均不顯著,呈現出先上升后下降的趨勢.α 在w(238U)≥100 μg·g-1時小于CK;rETRmax和Ik在w(238U)=100 μg·g-1時達到最大值,與CK 相比分別增加了14.76%和21.90%.在重金屬單獨作用下,α,rETRmax和Ik的變化不顯著.與CK 相比,Pb2+和Hg2+的存在提高了3 個指標;在高含量Cd2+作用下,α 和rETRmax小于對照,Ik大于對照;在高含量Ni2+作用下,α 增加了3.32%,rETRmax和Ik有所降低.
鈾和重金屬共存的處理中,當w(238U)≥100 μg·g-1時,向土壤中添加重金屬離子后,α 大于單獨的鈾處理,增幅大于1%;其中,w(238U)=150 μg·g-1,w(Cd2+)=25 μg·g-1增加了16.28%(P <0.05)時,增加顯著.rETRmax和Ik的變化一致,在加入高含量的重金屬后,這兩個指標都低于單獨的鈾處理.在同一鈾含量處理下,Hg2+濃度的變化對rETRmax和Ik的影響不大,隨著Cd2+,Pb2+和Ni2+濃度的增加,rETRmax和Ik逐漸減小.rETRmax和Ik都在w238U=150 μg·g-1,w(Ni2+)=500 μg·g-1時達到最低值,比單獨的鈾處理減少27.51%和34.95%,比對照減少24.39%和28.73%,變化不顯著.
單獨的鈾污染對α 的影響表現為低濃度促進,高濃度抑制,對rETRmax和Ik表現為促進作用.復合污染情況下,高含量鈾與重金屬離子共存提高了α,降低了rETRmax和Ik,增加了植物葉片對光能的利用效率,降低了植物的潛在最大電子傳遞速率和對強光的耐受能力.
3 討論
光合作用是植物生長的重要生理過程,葉綠素熒光與光合作用的各個反應過程都有緊密的關系,能夠反映內外環境條件的變化對光合系統Ⅱ(PSⅡ)的影響[20].正常環境中高等植物的Fv/Fm在0.832 ± 0.004范圍內,當植物受到脅迫時Fv/Fm會顯著下降,因此該指標被廣泛用于研究各種逆境環境對植物生長的影響,其降低的幅度可以反映逆境對植物光合作用的傷害程度[21-22].Fv/Fm的下降是由F0升高及Fm降低引起的.本實驗的結果表明,土壤中高濃度的鈾引起菊苣葉片F0顯著升高,Fv/Fm顯著下降,Fm有所降低,但是不顯著,說明高濃度鈾污染土壤對菊苣的光合生理有毒害作用;同時,Cd2+,Pb2+,Hg2+,Ni2+4 種重金屬對這3 個指標也有類似的影響,其中Cd2+的毒害較為嚴重.當土壤中有多重元素同時存在時,適當濃度的不同元素之間會發生交互作用,加強或者減弱單一元素對植物的影響.馮韶輝等(2013)等的研究結果顯示,Cd 與較高質量濃度的Cu(1.0 mg·L-1)聯合脅迫使熱帶海草泰來藻的光保護機制喪失,而Cd 與較低質量濃度的Cu(0.1 mg·L-1)聯合卻可以激發泰來藻的光保護機制[23].本實驗也表明,Cd2+,Pb2+,Hg2+和Ni2+都可以在一定程度上緩解高含量鈾對菊苣的傷害,主要表現為當w(238U)=150 μg·g-1時,含有重金屬的處理與單獨的鈾處理相比F0下降,Fv/Fm和Fm升高.
在單一鈾元素處理中,菊苣的ETR 和qP大于0 處理,Yield 和qN在低含量時大于0 處理,含量升高后則降低;低含量的Cd2+使Yield 和ETR 升高,高含量Cd2+卻使其降低,qP和qN則隨著Cd2+含量的升高而增加;而Pb2+,Hg2+和Ni2+處理使得這4 個參數均有所增加.這說明向土壤中添加適當濃度的鈾和重金屬有助于菊苣葉片的PSⅡ對原初光能的捕獲效率和電子傳遞;低含量的鈾和重金屬脅迫下,菊苣可以通過熱耗散的方式使光合機構免于或減輕傷害,而過高含量的鈾和重金屬脅迫,如w(238U)=150 μg·g-1和w(Ni2+)=500 μg·g-1,則會嚴重傷害這種光保護機制,使其喪失功能.鈾與重金屬共存時,qN升高,Yield、ETR 和qP降低,說明適量重金屬離子的存在可以減輕鈾對光保護機制的傷害程度,甚至使其有所增強,卻不利于菊苣葉片PSⅡ的原初光能捕獲效率和電子傳遞.另外,w(238U)≥100 μg·g-1時,單獨的鈾作用下,α 降低,rETRmax和Ik略有升高;向土壤中添加重金屬后α 升高,rETRmax和Ik降低,說明Cd2+,Pb2+,Hg2+和Ni2+可以緩解高濃度鈾對菊苣葉片的光能利用能力的傷害,卻減弱了鈾對菊苣葉片的電子傳遞速率和對強光的耐受能力的促進作用,甚至帶來負面影響.
與單一脅迫相比,聯合脅迫對植物生長的影響復雜得多[24-25],僅用一個指標來判斷鈾和重金屬復合污染對植物生長的影響程度是不夠的,只有同時結合多種指標才能更好地判斷植物的生長狀況.一般認為,重金屬的復合作用改變了金屬的生物有效性或毒性,危害更大[26-27].王天弘(2013)研究了鉛鎘復合污染對互花米草光合作用的影響,結果顯示鉛鎘相互作用使葉綠素的含量明顯降低[28].有研究表明Pb 對土壤中Cd的有效性起激活的作用,而Zn 卻對土壤中Cd 的有效性起抑制作用[29],說明不同的重金屬之間復合對植物造成的影響不同.本文研究結果認為,土壤中適量的重金屬可以減輕鈾對菊苣毒性作用,可能是因為重金屬改變了鈾的生物效應.
4 結論
聯合脅迫對植物生長的影響比較復雜.本研究表明:鈾與Cd2+,Pb2+,Hg2+和Ni2+復合污染對植物葉綠素熒光參數的變化有一定的影響.(1)在低鈾濃度下,Cd2+,Pb2+和Hg2+的存在降低了植物的Fv/Fm值,增加了對植物的脅迫,但Ni2+的存在幾乎沒有影響;在高鈾濃度下,重金屬離子的存在提高了Fv/Fm值,減輕了鈾離子單獨脅迫對植物的傷害;(2)高濃度的鈾和重金屬復合污染下qN升高,Yield、ETR 和qP降低,植物的自我光保護能力增強,天線色素的捕光功能下降,電子傳遞鏈受到影響;(3)高濃度鈾與重金屬復合污染使植物葉片的α 值升高,rETRmax和Ik下降,植物對光能,尤其是弱光的利用能力增強,潛在的最大電子傳遞速率和對強光的耐受能力降低.
[1]LOZANO J C,VERA T F,BLANCO R P,et al.A sequential method for the determination of210Pb,226Ra,and uranium and thorium radioisotopes by LSC and alpha-spectrometry[J].Appl Radiat Isot,2010,68(4/5):828-831.
[2]GRABOWSKI P,BEM H.Determination of210Po and uranium in high salinity water samples[J].J Radioanal Nucl Chem,2010,286(2):455-460.
[3]VERMAERCKE P,ARBOCCO F F,SNEYERS L,et al.Environmental monitoring for safeguards using κ0-standardized neutron activation analysis[J].IEEE T Nucl Sci,2010,57(5):2773-2776.
[4]GAVRILESCU M,VASILE P L,CRETESCU I.Characterization and remediation of soils contaminated with uranium[J].J Hazard Mater,2009,163(2/3):475-510.
[5]EBBS S D,BRADY D J,KOCHIAN L V.Role of uranium speciation in the uptake and translocation of uranium by plants[J].J Exp Bot,1998,49(324):1183-1190.
[6]劉 娟,李紅春,王 津,等.華南某鈾礦開采利用對地表水環境質量的影響[J].環境化學,2012,31(7):981-989.
[7]鐵柏清,袁 敏,唐美珍.美洲商陸(Phytolacca americana L.):一種新的Mn 積累植物[J].農業環境科學學報,2005,24(2):340-343.
[8]劉秀梅,聶俊華,王慶仁.6 種植物對Pb 的吸收及耐性研究[J].植物生態學報,2002,26(5):533-537.
[9]葉雪均,邱樹敏.3 種草本植物對Pb-Cd 污染水體的修復研究[J].環境工程學報,2010,4(5):1023-1026.
[10]唐永金,羅學剛,曾 峰,等.不同植物對高濃度鈾脅迫的響應與鈾富集植物篩選[J].核農學報,2013,27(12):1948-1954.
[11]DUSHENKOV S,VASUDEV D,KAPULNIK Y,et al.Enhanced accumulation of Pb in indian mustard by soil-applied chelating agents[J].Environ Sci Technol,1997,31(12):3468-3474.
[12]黃 慧,陳 宏.植物修復重金屬汞鎘鉻污染土壤的研究進展[J].中國農學通報,2010,26(24):326-329.
[13]陳同斌,韋朝陽,黃澤春,等.砷超富集植物蜈蚣草及其對砷的富集特征[J].科學通報,2002,47(3):207-210.
[14]唐永金,羅學剛,江世杰,等.鍶、銫、鈾對5 種植物種子發芽的影響[J].種子,2013,32(4):1-4.
[15]魯 艷,李新榮,何明珠,等.不同濃度Ni、Cu 處理對駱駝蓬光合作用和葉綠素熒光特性的影響[J].應用生態學報,2011,22(4):936-942.
[16]鐘鉬芝,王 丹,徐長合,等.螯合劑對鈾鎘污染土壤中蠶豆幼苗生理特性影響[J].農業環境科學學報,2011,30(4):639-644.
[17]唐永金,羅學剛.植物吸收和富集核素的研究方法[J].核農學報,2011,25(6):1292-1299.
[18]WHITE A J,CRITCHLEY C.Rapid light curves:A new fluorescence method to assess the state of the photosynthetic apparatus[J].Photosynth Res,1999,59(1):63-72.
[19]陳 梅,唐運來.高溫脅迫下莧菜的葉綠素熒光特性[J].生態學雜志,2013,32(7):1813-1818.
[20]高建欣,張文輝,王校鋒.Cd2+處理對5 個柳樹無性系氣體交換參數及葉綠素熒光參數的影響[J].西北植物學報,2013,33(9):1874-1884.
[21]楊 娟,謝 琳,張 新,等.干旱脅迫對轉IrrE 基因甘藍型油菜生理生化指標的影響[J].作物雜志,2012(1):26-30.
[22]梁文斌,薛生國,沈吉紅,等.錳脅迫對垂序商陸光合特性及葉綠素熒光參數的影響[J].生態學報,2010,30(3):619-625.
[23]馮韶輝,黃小平,張景平.Cu 和Cd 聯合脅迫對海草泰來藻光合作用的影響[J].生態學雜志,2013,32(6):1545-1550.
[24]朱啟紅,夏紅霞.蜈蚣草對Pb、Zn 復合污染的響應[J].環境化學,2012,3(7):1029-1035.
[25]LI L,HUANG X P,BORTHAKUR D,et al.Photosynthetic activity and antioxidative response of seagrass Thalassia hemprichii to trace metal stress[J].Acta Oceanol Sin,2012,31(3):98-108.
[26]孟昭福,薛澄澤,張增強,等.土壤中重金屬復合污染的表征[J].農業環境保護,1999,18(2):87-91.
[27]周東美,王玉軍,蒼 龍,等.土壤及土壤-植物系統中復合污染的研究進展[J].環境污染治理技術與設備,2004,5(10):1-8.
[28]王天弘.鉛鎘污染對互花米草光合作用的影響[J].安徽農業科學,2013,41(16):7149-7162.
[29]吳 丹,王友保,胡 珊,等.吊蘭生長對重金屬鎘鉛鋅復合污染土壤修復的影響[J].土壤通報,2013,44(5):1245-1252.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
當代陜西(2019年7期)2019-04-25 00:22:18
領導決策信息(2018年26期)2018-10-12 02:18:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
都市麗人(2015年5期)2015-03-20 13:33:49
