小麥–黑麥易位系T1BL.1RS在小麥品種中的分布及其與小麥赤霉病抗性的關聯
2016-03-03 09:01:51秦勝男顧世梁
作物學報 2016年3期
李 韜 鄭 飛 秦勝男 李 磊 顧世梁
揚州大學江蘇省作物遺傳生理重點實驗室 / 糧食作物現代產業技術協同創新中心 / 教育部植物功能基因組學重點實驗室 / 小麥研究中心, 江蘇揚州 225009
?
小麥–黑麥易位系T1BL.1RS在小麥品種中的分布及其與小麥赤霉病抗性的關聯
李韜*鄭飛秦勝男李磊顧世梁
揚州大學江蘇省作物遺傳生理重點實驗室 / 糧食作物現代產業技術協同創新中心 / 教育部植物功能基因組學重點實驗室 / 小麥研究中心, 江蘇揚州 225009
摘要:黑麥1R染色體短臂(1RS)攜帶條銹病、葉銹病、稈銹病、白粉病和蚜蟲等抗性基因。為了檢測1RS上是否攜帶與赤霉病抗性相關的基因, 本研究采用1RS特異標記Xscm9對192個來自不同國家的品種/系構成的小麥自然群體和1個重組自交系(RIL)群體(寧7840與Chokwang雜交的F7群體, 共184個系)進行了分子檢測, 并在2011—2013年采用單花滴注法于溫室中進行赤霉病抗性鑒定。結果發現, 自然群體中22個品種攜帶1RS, 攜帶1RS的株系三季赤霉病平均病小穗率(PSS)均顯著低于不攜帶1RS株系的PSS (P<0.01), 表明1RS對降低病小穗率有顯著作用。分子標記和基因組原位雜交(GISH)檢測結果表明, 寧7840攜帶1RS。通過對寧7840/Chokwang衍生的RIL群體進行赤霉病抗性鑒定和基因型分析, 發現不論主效赤霉病抗性基因Fhb1 (標記Xsts142)存在與否, 攜帶1RS株系的PSS顯著低于不攜帶1RS株系的PSS (P<0.01); 方差分析表明, 寧7840攜帶的Fhb1與1RS在赤霉病抗擴展性上無顯著互作(P>0.05)。因此認為, 黑麥1RS染色體很可能攜帶赤霉病擴展抗性相關基因, 與Fhb1基因有累加效應。
關鍵詞:小麥; 黑麥; T1BL.1RS易位系; 赤霉病
本研究由國家轉基因生物新品種培育科技重大專項(2012ZX08009003-004), 國家自然科學基金項目(31171537)和江蘇省高校優勢學科建設工程項目(PAPD)資助。
This study was supported by the National Major Project of Breeding for New Transgenic Organisms (2012ZX08009003-004), the National Natural Science Foundation of China (31171537), and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
在小麥遺傳改良中, 來自小麥近緣種黑麥的基因發揮了重要作用, 尤其以黑麥1R染色體短臂(1RS)應用最為廣泛[1]。該短臂上攜帶抗條銹病基因Yr9、抗葉銹病基因Lr26、抗稈銹病基因Sr31以及抗白粉病基因Pm8和Pm17, 目前已經將這些抗病基因轉移到小麥栽培品種中[2-3]。1RS攜帶俄羅斯蚜蟲[4-5]和麥二叉蚜[6-7]的抗性基因。1RS對增加小麥根系生物量[8]、提高小麥產量[9]、促進小麥適應性[10]也有作用。一般認為, T1BL.1RS易位系對小麥加工品質有負向影響[12], 1RS攜帶的部分抗病基因已被病原菌毒性生理小種克服[11], 但據肖永貴等[12]報道, T1BL.1RS易位系白粉病抗性和千粒重顯著高于非易位系[13]。因此, T1BL.1RS易位系在小麥育種中的利用價值仍值得深入研究。目前, 利用1RS特異鑒別標記Xscm9可以快速鑒別和有效區分來源不同的T1AL.1RS和T1BL.1RS易位系[14-16]。
小麥赤霉病是由禾谷鐮刀菌(Fusarium graminearum)引起的一種破壞性病害, 在濕潤和半濕潤地區廣泛發生[17]。在我國, 赤霉病一直是淮河以南及長江中下游麥區發生最嚴重的病害之一。東北春麥區、黃淮海平原麥區和西北麥區也曾多次發生大流行, 造成嚴重損失。自2000年以來, 我國先后9次發生小麥赤霉病大流行, 尤以2012年最為嚴重, 估計受害小麥面積達1000萬公頃, 部分田塊產量損失高達100%。近年來, 隨著玉米種植面積不斷擴大及秸稈還田技術的推廣, 留在田間的作物殘茬成為赤霉病菌繁殖的良好寄主, 致使小麥赤霉病有逐漸加重的趨勢[18], 不僅造成產量損失, 而且使食品安全面臨巨大挑戰。目前, 研究人員已經在普通小麥或其近緣種中定位了超過100多個抗性QTL[19], 但只對其中極少數主效QTL進行了比較精細的定位, 并用于抗病育種, 如3BS染色體上的Fhb1位點、6B染色體上Fhb2位點和7A染色體上的QTL等, 其中以蘇麥3號攜帶的Fhb1應用最為廣泛[20-23]。據報道,偃麥草、披堿草等小麥近緣種屬中也攜帶赤霉病抗性基因[24-26], 但黑麥1RS染色體與小麥赤霉病抗性的關系, 國內外至今未見報道。本研究主要通過小麥單花滴注鑒定和1RS特異標記分析, 在小麥自然群體和重組近交系群體(RIL)中分別解析1RS與赤霉病抗性的關系, 進一步拓展對T1BL.1RS功能的認識。
1 材料與方法
1.1供試材料
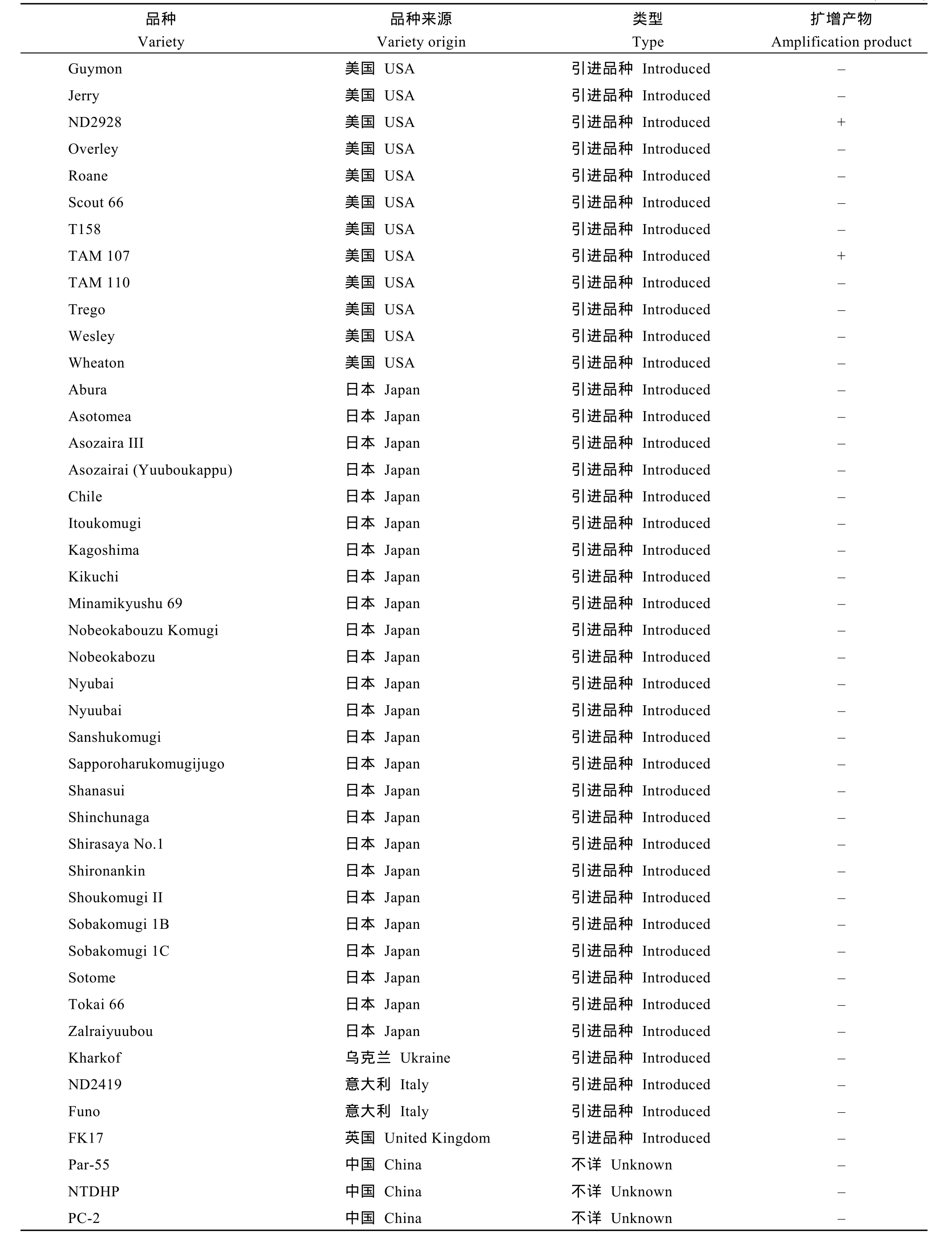
自然群體由192份小麥品種(系)組成, 包括中國小麥地方品種48個、育成品種74個, 日本品種27個,美國品種26個, 其他國家或機構的引進品種16個(見附表)。重組近交系群體為寧7840/Chokwang經單籽粒傳法衍生的F7代, 由184個系構成。寧7840是我國抗赤霉病小麥品種, 其系譜為Aurora/安徽11//蘇麥3號; Chokwang是韓國小麥品種, 對赤霉病表現中抗。
2011—2013年連續3季, 將自然群體、RIL群體及其親本種植于美國堪薩斯州立大學溫室, 所有材料先在4。C下春化處理7周, 然后移栽到4 cm × 4 cm塑料盆缽中, 于溫室(晝22±5℃/夜17±2℃)隨機區組排列, 每系2個重復(2盆), 每重復5株。
1.2赤霉病菌接種和抗性鑒定方法
參照Yu等[27]描述的方法準備赤霉菌孢子懸浮液(5000孢子 mL-1), 采用單花滴注法接種。于小麥始花期, 用注射器吸取10 μL孢子液注射入麥穗中間小穗的一個小花中, 每盆接種6~10穗。單花接種后立即置22~26。C迷霧室保濕48~72 h, 出現癥狀后從迷霧室移至溫室。接種21 d后調查病小穗數, 計算病小穗率。病小穗率(%) = (病小穗數/總小穗數) × 100。用Matlab軟件(MathWorks Inc., Natick, MA, USA)進行表型數據的相關分析及方差分析。
1.31RS特異引物擴增與分析
取小麥幼苗, 采用CTAB法提取基因組DNA[28]。參照Yu等[27]報道的PCR體系和Touch-down PCR擴增程序, 擴增引物Xscm9是1RS的特異鑒別標記,能夠擴增出207 bp的特異片段[15-16]。用ABI 3730 DNA分析儀(Applied Biosystems, FosterCity, CA, USA)分離PCR擴增產物, 用GeneMarker 1.6 (Softgenetics Inc. LLC, State College, PA)分析分子標記數據。
1.4基因組原位雜交(GISH)分析
以黑麥DNA片段作為探針, 以寧7840及其親本的DNA作為模板, 進行熒光原位雜交。參照Gustafson[29]描述的方法進行探針標記、雜交和洗滌,以CCD攝像機(Olympus, Japan)捕獲圖像。
2 結果與分析
2.1T1BL.1RS易位系在自然群體中分布及與赤霉病抗性的關系
用T1BL.1RS特異分子標記Xscm9對自然群體192份材料進行檢測, 在22個品種(系)中擴增出預期的207 bp特異帶(見附表), 占品種總數的11.5%, 包
括12個我國小麥育成品種(系), 1個我國小麥地方品種, 6個美國品種, 2個阿根廷品種和1個俄羅斯品種。在日本品種中, 未檢測到Xscm9標記陽性品種。
2011—2013年連續3季進行赤霉病菌人工接種的抗性調查, 其平均PSS分別為0.25 (0.03~1.00)、0.40 (0.05~1.00)和0.52 (0.06~1.00)。PSS在品種間和年度間差異顯著(P<0.001), 而且存在品種×年度互作(P<0.001), 但年度間PSS表現顯著正相關(P<0.001),相關系數分別為0.75、0.60和0.68。
對比分子標記檢測和抗病性鑒定結果, 發現2011、2012和2013年攜帶1RS材料的PSS平均值分別為0.34、0.35和0.42, 而不攜帶1RS材料的平均值分別為0.42、0.48和0.46 (圖1)。t測驗結果顯示, 攜帶1RS材料的PSS顯著低于不攜帶1RS材料的PSS (t=10.249, t0.05=3.84)。
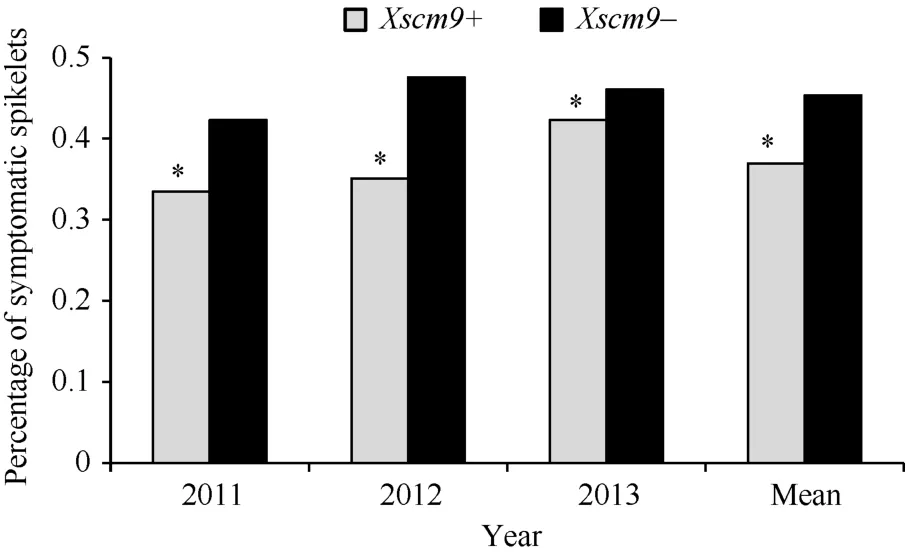
圖1 自然群體中Xscm9陽性和陰性品種間病小穗率Fig. 1 Percentage of symptomatic spikelets of the two groups contrasting in 1RS*表示兩類品種間有顯著差異(P<0.05)。* indicates significant difference between the Xscm9 positive and negative groups at P<0.05.
2.2RIL群體中1RS與赤霉病抗性的關系
在寧7840及其親本之一Aurora中都檢測到Xscm9的特異擴增產物, 但在寧7840另兩個親本蘇麥3號和安徽11中未檢測到該擴增產物(見附表)。結合系譜分析, 初步推定寧7840中1RS的易位片段來自Aurora。該推斷被以黑麥DNA為探針的GISH實驗證實, GISH圖像顯示Aurora和寧7840的染色體上帶有熒光標記, 而安徽11和蘇麥3號染色體上沒有熒光信號(圖2)。
寧7840高抗赤霉病, 攜帶主效抗性 QTL Fhb1和易位染色體T1BL.1RS; Chokwang中抗赤霉病, Xscm9擴增為陰性, 不攜帶1RS (見附表)。由這兩個品種衍生的RIL群體各系的PSS在基因型間和年度間存在顯著差異(P<0.001), 并且二者互作顯著(P< 0.001); 3年鑒定結果一致(P<0.001), 相關系數分別為0.55、0.60和0.56。2011、2012和2013年平均PSS分別為0.47、0.34和0.49, 變異幅度分別為0.06~1.00、0.06~0.83和0.06~1.00; 2011年與2013年赤霉病發病情況相似, 2012年病情明顯較輕。

圖2 寧7840及其親本的GISH圖像Fig. 2 GISH for Ning7840 and its parentsA: 寧7840; B: 安徽11; C: 蘇麥3號; D: Aurora。箭頭示黑麥1RS染色體臂。A: Ning 7840; B: Anhui 11; C: Sumai 3; D: Aurora.Arrows show the chromatin from rye.
寧7840的3BS上存在一個主效赤霉病抗性QTL Fhb1, 能夠解釋16.7%~28.6%的PSS表型變異[19]。STS標記Xsts142與Fhb1位點緊密連鎖[30], 且在雙親間有多態性。由表1可見, Fhb1和1RS均與抗擴展性顯著關聯(P<0.001), 但兩者之間沒有顯著互作(P=0.48)。
從RIL群體的Xsts142陽性株系(攜帶Fhb1)看,同時還攜帶1RS株系的平均PSS為0.25, 只攜帶Fhb1株系的平均PSS為0.35 (表2), 二者差值(0.10) 為1RS的估計貢獻率(P<0.01)。同理, 從Xsts142陰性株系估計, 1RS的貢獻率為0.07 (P<0.05); 從Xscm9陽性株系估計, Fhb1的貢獻率為0.26 (P<0.001); 從Xscm9陰性株系估計, Fhb1的貢獻率為0.230 (P<0.001)。可見, 不論T1BL.1RS存在與否, Fhb1株系的PSS均顯著小于非Fhb1株系; 不論Fhb1存在與否, 1RS株系的PSS始終顯著小于非1RS的株系(圖3)。1RS的估計貢獻率平均值為0.08, Fhb1的估計貢獻率平均值為0.24, 說明Fhb1和1RS對赤霉病的貢獻率大約分別為0.24和0.08, 1RS雖然可增強赤霉病抗性, 但其效應小于Fhb1。

表1 Fhb1和1RS組合的赤霉病擴展抗性Table 1 Analysis of variance of 1RS and Fhb1 to type 2 resistance to Fusarium head blight
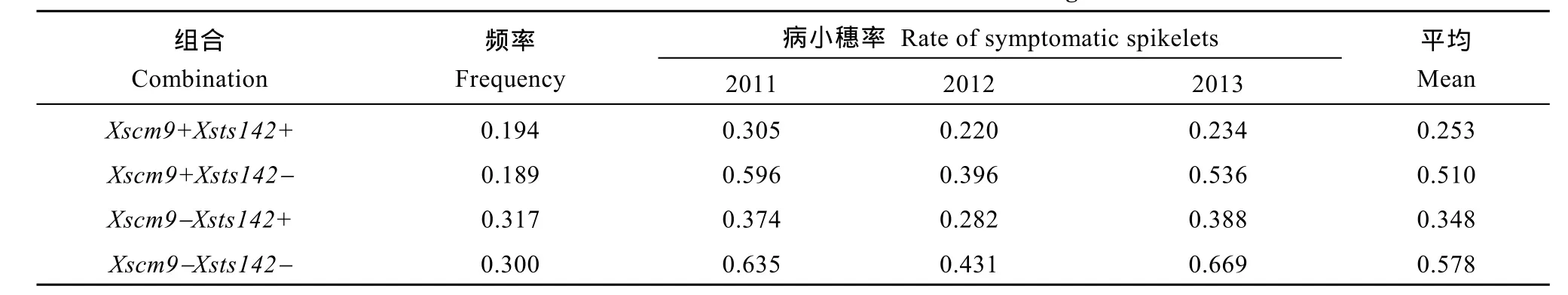
表2 Fhb1、1RS不同組合型品種的赤霉病病小穗率Table 2 Rate of symptomatic spikelets of combinations contrasting in Fhb1 and 1RS
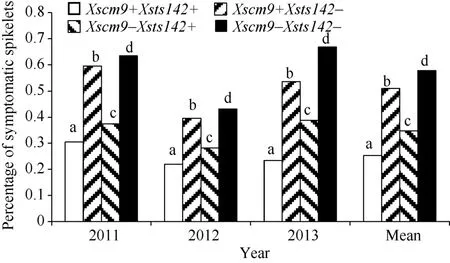
圖3 Fhb1與1RS不同組合型品種的赤霉病擴展抗性Fig. 3 Type 2 resistance for groups of different Fhb1 and 1RS combinations不同字母表示兩類品種間有顯著差異(P<0.05)。Bars superscripted by different letters are significantly different between the Xscm9 positive and negative groups at P<0.05.
3 討論
由于1RS攜帶抗白粉, 抗條銹、葉銹、稈銹, 以及抗蚜蟲等基因, T1BL.1RS易位系曾在生產上廣泛應用[31], 但部分抗病基因被新出現的毒性小種所克服, 因此目前育種中銹病和白粉病防治很少依賴T1BL.1RS攜帶的抗病基因。1RS上攜帶的赤霉病抗性基因與白粉病、銹病及蚜蟲等抗性位點的遺傳關系是一因多效還是緊密連鎖還需要進一步的研究。本研究在192個小麥品種(系)中, 通過1RS特異標記Xscm9鑒定到攜帶T1BL.1RS的22個品種, 通過對Xscm9陽性品種寧7840及其親本Aurora的GISH實驗, 驗證了Xscm9用于鑒定1RS易位系的可靠性。T1BL.1RS在自然群體中的頻率約為11.5%, 包括21個小麥育成品種(系)和一個中國地方品種火燒麥。育成品種中1RS可能是育種過程中通過親本組配和雜交實現了T1BL.1RS定向導入, 而火燒麥中的1RS易位系可能與異交有關, 具體原因有待進一步分析。在自然群體中, 1RS與非1RS的赤霉病病小穗率平均相差0.084, 估計1RS貢獻率為8.4%; 而在RIL群體中, 1RS與非1RS病小穗率平均相差0.081, 估計貢獻率為8.1%, 二者非常接近, 因此推斷1RS可能攜帶一個微效赤霉病抗性基因, 其效應顯著小于Fhb1位點。在本研究中, 寧7840/Chokwang衍生的RIL群體中Fhb1的估計貢獻率為24.3%, 介于文獻中報道的Fhb1貢獻率16.7%~28.6%[19]。這一結果表明, 1RS對降低病小穗率和提高赤霉病抗性有正向效應, 1RS與Fhb1沒有顯著的互作效應(表1), 二者效應可以累加(表2和 圖3), 因此更有利于育種實踐中同時或單獨利用Fhb1和T1BL.1RS易位系。
4 結論
1RS對增強赤霉病抗性有正向效應。本文192
份國內外種質資源T1BL.1RS的鑒定結果, 對育種家選配雜交親本提供了有益的信息。
致謝: 自然群體赤霉病鑒定由張大棟博士(美國堪薩斯州立大學)協助完成, 本實驗材料由Bai Guihua博士(美國堪薩斯州立大學)提供, 熒光原位雜交實驗由Bernd Friebe博士(美國堪薩斯州立大學)協助完成, 謹致謝意。
References
[1] Rabinovich S. Importance of wheat-rye translocations for breeding modern cultivar of Triticum aestivum L. Euphytica, 1998, 100: 323–340
[2] Mago R, Miah H, Lawrence G J, Wellings C R, Spielmeyer W, Bariana H S, McIntosh R A, Pryor A J, Ellis J G. High-resolution mapping and mutation analysis separate the rust resistance genes Sr31, Lr26 and Yr9 on the short arm of rye chromosome 1. Theor Appl Genet, 2005, 112: 41–50
[3] Singh N, Shepherd K, McIntosh R. Linkage mapping of genes for resistance to leaf, stem and stripe rusts and ω-secalins on the short arm of rye chromosome 1R. Theor Appl Genet, 1990, 80: 609–616
[4] Anderson G R, Papa D, Peng J, Tahir M, Lapitan N L. Genetic mapping of Dn7, a rye gene conferring resistance to the Russian wheat aphid in wheat. Theor Appl Genet, 2003, 107: 1297–1303
[5] Peng J, Wang H, Haley S D, Peairs F B, Lapitan N L V. Molecular mapping of the Russian wheat aphid resistance gene Dn2414 in wheat. Crop Sci, 2007, 47: 2418–2429
[6] Lu H, Rudd J C, Burd J D, Weng Y. Molecular mapping of greenbug resistance genes Gb2 and Gb6 in T1AL.1RS wheat–rye translocations. Plant Breed, 2010, 129: 472–476
[7] Mater Y, Baenziger S, Gill K, Graybosch R, Whitcher L, Baker C, Specht J, Dweikat I. Linkage mapping of powdery mildew and greenbug resistance genes on recombinant 1RS from ‘Amigo’ and ‘Kavkaz’ wheat–rye translocations of chromosome 1RS.1AL. Genome, 2004, 47: 292–298
[8] Maheepala D C, Ehdaie B, Waines J G. Yield performance of wheat isolines with different dosages of the short arm of rye chromosome 1. J Agron Crop Sci, 2015, 201: 152–160
[9] Howell T, Hale I, Jankuloski L, Bonafede M, Gilbert M, Dubcovsky J. Mapping a region within the 1RS.1BL translocation in common wheat affecting grain yield and canopy water status. Theor Appl Genet, 2014, 127: 2695–2709
[10] Ehdaie B, Whitkus R W, Waines J G. Root biomass, water-use efficiency, and performance of wheat-rye translocations of chromosomes 1 and 2 in spring bread wheat ‘Pavon’. Crop Sci, 2003, 43: 710–717
[11] Yang M Y, Ren T H, Yan B J, Li Z, Ren Z L. Diversity resistance to Puccinia striiformis f. sp. tritici in rye chromosome arm 1RS expressed in wheat. Genet Mol Res, 2014, 13: 8783–8793
[12] 劉建軍, 肖永貴, 程敦公, 李豪圣, 劉麗, 宋健民, 劉愛峰, 趙振東, 何中虎. 利用揉面特性鑒定小麥1BL/1RS易位系. 作物學報, 2009, 35: 79–86
Liu J J, Xiao Y G, Cheng D G, Li H S, Liu L, Song J M, Liu A F, Zhao Z D, He Z H. Identification of 1BL/1RS translocation based on Mixograph parameters in common wheat. Acta Agron Sin, 2009, 35: 79–86 (in Chinese with English abstract)
[13] 肖永貴, 閻俊, 何中虎, 張勇, 張曉科, 劉麗, 李天富, 曲延英,夏先春. 1BL/1RS易位對小麥產量性狀和白粉病抗性的影響及其QTL分析. 作物學報, 2006, 32: 1636–1641
Xiao Y G, Yan J, He Z H, Zhang Y, Zhang X K, Liu L, Li T F, Qu Y Y, Xia X C. Effect of 1BL.1RS translocation on yield traits and powdery mildew resistance in common wheat and QTL analysis. Acta Agron Sin, 2006, 32: 1636–1641 (in Chinese with English abstract)
[14] 余利, 何方, 陳桂玲, 崔法, 亓曉蕾, 王洪鋼, 李興鋒. 利用1RS特異標記和染色體原位雜交技術鑒定小麥1BL·1RS 易位系. 作物學報, 2011, 37: 563–569
Yu L, He F, Chen G L, Cui F, Qi X L, Wang H G, Li X F. Identification of 1BL·1RS wheat–rye chromosome translocations via 1RS specific molecular markers and genomic in situ hybridization. Acta Agron Sin, 2011, 37: 563–569 (in Chinese with English abstract)
[15] Saal B, Wricke G. Development of simple sequence repeat markers in rye (Secale cereale L.). Genome, 1999, 42: 964–972
[16] Weng Y, Azhaguvel P, Devkota R N, Rudd J C. PCR-based markers for detection of different sources of 1AL.1RS and 1BL.1RS wheat–rye translocations in wheat background. Plant Breed, 2007, 126: 482–486
[17] Bai G, Shaner G. Management and resistance in wheat and barley to Fusarium head blight. Annu Rev Phytopathol, 2004, 42: 135–161
[18] 曾娟, 姜玉英. 2012年我國小麥赤霉病暴發原因分析及持續監控與治理對策. 中國植保導刊, 2013, 33(4): 38–41
Zeng J, Jiang Y Y. The causal factors for the epidemics of wheat Fusarium head blight in the year of 2012 in China and the strategies for continuous monitoring and prevention. China Plant Prot, 2013, 33(4): 38–41 (in Chinese)
[19] Buerstmayr H, Ban T, Anderson J A. QTL mapping and marker-assisted selection for Fusarium head blight resistance in wheat: a review. Plant Breed, 2009, 128: 1–26
[20] Draeger R, Gosman N, Steed A, Chandler E, Thomsett M, Schondelmaier J, Buerstmayr H, Lemmens M, Schmolke M, Mesterhazy A. Identification of QTLs for resistance to Fusarium head blight, DON accumulation and associated traits in the winter wheat variety Arina. Theor Appl Genet, 2007, 115: 617–625
[21] Cuthbert P A, Somers D J, Brulé-Babel A. Mapping of Fhb2 on chromosome 6BS: a gene controlling Fusarium head blight field resistance in bread wheat (Triticum aestivum L.). Theor Appl Genet, 2007, 114: 429–437
[22] Liu S, Zhang X, Pumphrey M O, Stack R W, Gill B S, Anderson J A. Complex microcolinearity among wheat, rice, and barley revealed by fine mapping of the genomic region harboring a major QTL for resistance to Fusarium head blight in wheat. Funct Integr Genomic, 2006, 6: 83–89
[23] Ma H, Zhang K, Gao L, Bai G, Chen H, Cai Z, Lu W. Quantitative trait loci for resistance to Fusarium head blight and deoxynivalenol accumulation in Wangshuibai wheat under field conditions. Plant Pathol, 2006, 55: 739–745
[24] Zeng J, Cao W, Hucl P, Yang Y, Xue A, Chi D, Fedak G. Molecular cytogenetic analysis of wheat–Elymus repens
introgression lines with resistance to Fusarium head blight. Genome, 2013, 56: 75–82
[25] Turner M K, DeHaan L R, Jin Y, Anderson J A. Wheatgrass–wheat partial amphiploids as a novel source of stem rust and Fusarium head blight resistance. Crop Sci, 2013, 53: 1994–2005 [26] Zhang X, Shen X, Hao Y, Cai J, Ohm H W, Kong L. A genetic map of Lophopyrum ponticum chromosome 7E, harboring resistance genes to Fusarium head blight and leaf rust. Theor Appl Genet, 2011, 122: 263–270
[27] Yu J B, Bai G H, Cai S B, Ban T. Marker-assisted characterization of Asian wheat lines for resistance to Fusarium head blight. Theor Appl Genet, 2006, 113: 308–320
[28] Porebski S, Bailey L G, Baum B R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol Biol Rep, 1997, 15: 8–15
[29] Gustafson J, Dille J. Chromosome location of Oryza sativa recombination linkage groups. Proc Natl Acad Sci USA, 1992, 89: 8646–8650
[30] Liu S, Zhang X, Pumphrey M O, Stack R W, Gill B S, Anderson J A. Complex microcolinearity among wheat, rice, and barley revealed by fine mapping of the genomic region harboring a major QTL for resistance to Fusarium head blight in wheat. Funct Integr Genomic, 2006, 6: 83–89
[31] Kim W, Johnson J W, Baenziger P S, Lukaszewski A J, Gaines C S. Agronomic effect of wheat–rye translocation carrying rye chromatin (1R) from different sources. Crop Sci, 2004, 44: 1254–1258

附表 1RS特異標記Xscm9在供試群體中的分布Supplementary table Distribution of the 1RS-specific marker Xscm9 in the wheat population tested
(續附表)

(續附表)

(續附表)
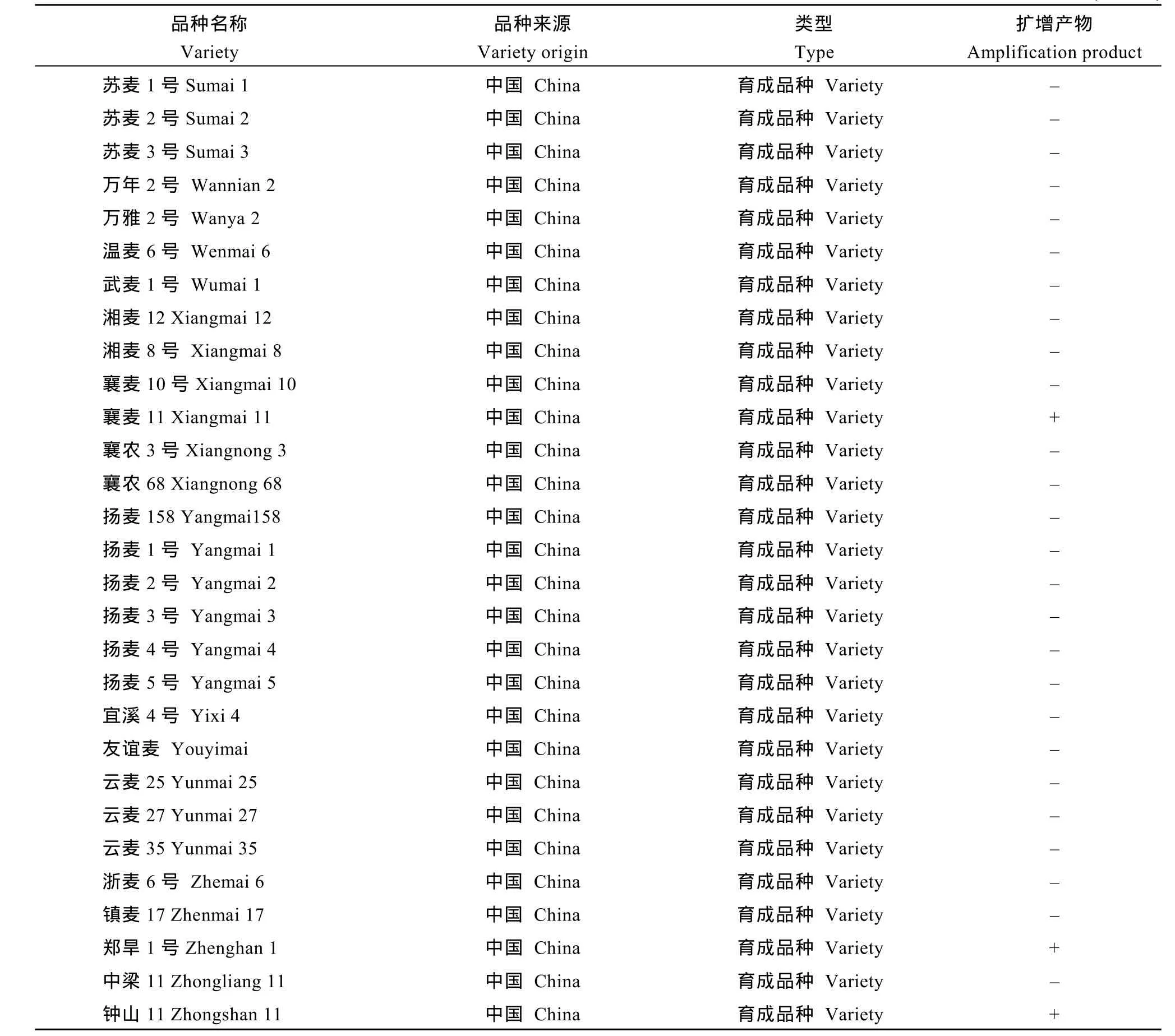
(續附表)
URL: http://www.cnki.net/kcms/detail/11.1809.S.20151218.0915.018.html
Distribution of Wheat–Rye Translocation Line T1BL.1RS in Wheat and Its Association with Fusarium Head Blight Resistance
LI Tao*, ZHENG Fei, QIN Sheng-Nan, LI Lei, and GU Shi-Liang
Jiangsu Provincial Key Laboratory of Crop Genetics and Physiology / Co-innovation Center for Modern Production Technology of Grain Crops / Key Laboratory of Plant Functional Genomics of Ministry of Education, Yangzhou University, Yangzhou 225009, China
Abstract:The short arm of 1R chromosome (1RS) of rye carries resistant genes to stripe rust, leaf rust, stem rust, powdery mildew and aphids. To understand if 1RS also mediates resistance to wheat Fusarium head blight (FHB), we genotyped a panel of 192 wheat accessions from diverse geographic regions and a population of recombinant inbred lines (RIL) consisting of 184 lines developed from the cross of Ning 7840 and Chokwang by 1RS-specific molecular marker Xscm9 and evaluated FHB severities in three consecutive seasons in greenhouses using single floret inoculation method. The results demonstrated that 22 of 192 accessions carried a T1BL.1RS translocation, and the mean FHB severity (PSS) of varieties carrying T1BL.1RS translocation was significantly lower than that of lines without the translocation across the three experiments (P < 0.05), indicating 1RS may have a positive effect on reducing FHB severity. 1RS-specific marker Xscm9 and Genome in situ hybridization (GISH) showed Ning 7840 carries T1BL.1RS translocation. In the population of RILs, irrespective of Fhb1 locus, the mean PSS of lines with T1BL.1RS translocation was significantly lower than that of those lines without T1BL.1RS. The effects of Fhb1 and 1RS on FHB resistance were additive and the interactions between them were not significant (P = 0.48). The results of this study suggested that 1RS of rye most likely carries the genes resistant to FHB.
Keywords:Wheat; Rye; T1BL.1RS translocation; Fusarium head blight
收稿日期Received(): 2015-09-11; Accepted(接受日期): 2015-11-20; Published online(網絡出版日期): 2015-12-18.
通訊作者*(Corresponding author): 李韜, E-mail: taoli@yzu.edu.cn
DOI:10.3724/SP.J.1006.2016.00320
