秈稻資源WD15515中抗褐飛虱QTL的定位研究
2016-03-03 09:01:56石少階王卉穎上官欣欣劉丙芳荊勝利波陳榮智祝莉莉何光存武漢大學生命科學學院雜交水稻國家重點實驗室湖北武漢430072
作物學報 2016年3期
關鍵詞:水稻
鄧 釗 石少階 王卉穎 上官欣欣 劉丙芳 荊勝利 杜 波陳榮智 祝莉莉 何光存武漢大學生命科學學院雜交水稻國家重點實驗室, 湖北武漢430072
?
秈稻資源WD15515中抗褐飛虱QTL的定位研究
鄧釗**石少階**王卉穎上官欣欣劉丙芳荊勝利杜波陳榮智祝莉莉何光存*
武漢大學生命科學學院雜交水稻國家重點實驗室, 湖北武漢430072
摘要:褐飛虱是我國水稻生產上最嚴重的蟲害之一, 培育和種植抗褐飛虱水稻品種是控制褐飛虱的有效途徑。WD15515是一份高抗褐飛虱的秈稻種質資源。利用9311與WD15515雜交培育了F2群體, 對F2代100個植株進行SSR分子標記分析, 測定植株上褐飛虱的蜜露分泌量、蟲體增重量和增重比, 作為抗蟲性指標。通過QTL IciMapping 3.0進行作圖分析, 在第2、第4、第9染色體上共檢測到4個抗褐飛虱QTL。其中第2染色體上檢測到2個QTL, 以蜜露分泌量檢測到的qBph2-1位于SSR標記RM71–RM6911之間, LOD值為3.68, 表型貢獻率為11.08%; 以蟲體增重量和增重比檢測到的qBph2-2位于標記RM6911–RM521之間, LOD值分別為3.31、4.05, 表型貢獻率分別為7.81%、9.38%。以蜜露分泌量、蟲體增重量和增重比為指標, 在第4染色體上檢測到qBph4, 定位于標記RM16996–RM17075之間, LOD值分別為11.11、13.81、15.41, 表型貢獻率達到44.38%、45.24%、52.40%。同樣, 以蜜露分泌量、蟲體增重量和增重比在第9染色體上檢測到qBph9, 定位于標記RM219–RM6444之間, LOD值分別為2.59、4.04、3.63, 表型貢獻率分別為10.91%、12.39%、10.01%。上述結果表明, qBph4是一個抗褐飛虱主效基因。本項研究結果為抗褐飛虱水稻育種提供了新的基因資源。
關鍵詞:水稻; 抗性; 褐飛虱; 數量性狀基因座; 遺傳圖譜
本研究由國家自然科學基金項目(31230060)和國家重點基礎研究發展計劃(973計劃)項目(2013CBA01403)資助。
This study was supported by the National Natural Science Foundation of China (31230060) and the National Basic Key Research Program (973 Program) (2013CBA01403).
**同等貢獻(Contributed equally to this work)
第一作者聯系方式: 鄧釗, E-mail: dengzhao@lpht.com.cn; 石少階, E-mail: 724054008@qq.com
褐飛虱(Nilaparvata lugens Stal, 簡稱BPH)屬同翅目飛虱科昆蟲, 是水稻的主要害蟲。褐飛虱通過針狀口器吸食水稻維管束鞘汁液, 造成稻株發黃、倒伏甚至枯死。褐飛虱在取食時傳播水稻草狀叢矮病和齒葉矮縮病, 同時也促使水稻紋枯病、小球菌核病的傳播。隨著高產不抗蟲水稻品種的推廣, 以及水稻種植方式和耕種制度的改變, 褐飛虱發生頻次增加, 危害程度加大, 已經成為我國和世界上水稻生產的首要蟲害, 對糧食生產安全造成嚴重威脅[1-2]。實踐表明, 選育和推廣具有抗蟲能力的水稻品種是防治褐飛虱最為經濟、有效的方法。因此, 發掘新的抗源和抗性基因對于培育具有持久抗蟲性的品種、防治褐飛虱具有重要意義。
迄今為止, 國內外已報道定位30個抗褐飛虱主效基因[3], 其中Bph14、Bph26和Bph3等已被克隆[4-6]。多個抗褐飛虱基因已廣泛用于水稻育種并在水稻生產中推廣應用。據國際水稻研究所報道, 一些只含單一主效抗蟲基因的品種在推廣種植幾年后, 會因褐飛虱種群變化而逐漸喪失原有的抗性, 具有多個抗蟲位點或微效位點的品種, 抗性則更為持久。水稻品種IR64含有主效抗蟲基因Bph1, 但在生產應用中較其他含Bph1的品種抗性更為持久。深入分析發現, IR64除含Bph1外, 還含多個抗蟲QTL[7]。到目前為止, 已利用不同的水稻材料鑒定了多個抗褐飛虱QTL, Huang等[8]研究抗性材料B5, 定位了2個主效QTL Qbp1和Qbp2和3個微效QTL。Xu等[9]以Lemont/Teqing的RIL群體材料, 定位了7個抗褐飛虱QTL; 蘇昌潮等[10]應用Nipponbare/Kasalath// Nipponbare回交重組自交系群體定位了3個抗褐飛虱QTL。上述研究結果, 為水稻抗褐飛虱育種提供了豐富的基因資源。
我們的前期研究結果表明, 秈稻品種資源WD15515對褐飛虱表現高抗(石少階, 未發表資料)。本研究構建了9311/WD15515 F2群體, 檢測并分析抗褐飛虱的基因位點, 并用BC1F1群體驗證了定位結果。研究結果為水稻抗褐飛虱分子輔助育種, 并為精細定位和克隆抗褐飛虱新基因打下了基礎。
1 材料與方法
1.1試驗材料
以對褐飛虱敏感的品種9311為母本, 以WD15515為父本, 通過雜交和自交得到了F2群體, 選取其中的100株進行抗性鑒定和分子標記分析。與此同時, 以F1與9311回交得到BC1F1群體, 用于F2群體QTL定位結果的驗證。于2006年在武漢田間采集褐飛虱, 在TN1稻株上飼養繁殖。
1.2水稻的褐飛虱抗性評定
通過測定褐飛虱在稻株上的表現值, 評價抗蟲性。于塑料桶中種植供試水稻的親本、F2及BC1F1,生長至40 d左右進行抗蟲性評價。收集新近羽化的褐飛虱短翅雌蟲, 制作30 mm × 30 mm蠟袋并放入雌蟲1頭, 然后將蠟袋綁定至水稻莖稈上, 由褐飛虱取食水稻(圖1), 48 h后, 終止取食, 取下蠟袋。在褐飛虱取食前和取食后, 在十萬分之一精度天平上分別稱取蠟袋重量和蟲體重量。取食前后蠟袋的重量差即為褐飛虱蜜露分泌量, 取食前后蟲體重量之差即為蟲體增重量, 用蟲體增重量除以取食前蟲體重量即為蟲體增重比。每個單株上的放蟲實驗重復3次以上, 確保取得精確、可重復的結果。以蜜露分泌量、蟲體增重量和蟲體增重比的多重復平均值來評價每個水稻植株的抗蟲性水平。

圖1 水稻抗褐飛虱性能鑒定示意圖Fig. 1 Schematic diagram showing BPH-resistance evaluation A: 每個水稻莖稈上綁定3個放蟲蠟袋。B: 鑒定結束后從蠟袋中取出待稱重的褐飛虱。A: three parafilm bags with BPH were bound in each stem of rice. B: BPH insects taken from parafilm bags.
1.3DNA抽提
以CTAB法制備水稻DNA。取新鮮水稻葉片200~300 mg至2 mL離心管, 于植物組織研磨儀中破碎, 加入800 μL CTAB提取液, 于65℃水浴30 min, 每5 min搖勻一次; 加入800 μL氯仿, 12 000×g離心10 min; 轉移上清液至另一1.5 mL離心管, 加600 μL異丙醇, 12 000×g離心10 min; 棄上清液, 加800 μL 70%的乙醇, 12 000×g離心5 min; 棄70%乙醇, 于真空干燥機中烘干; 加100~200 μL 0.1×TE,溶解后于4℃保存備用。
1.4SSR標記分析
根據GRAMENE網站(http://www.gramene.org/ markers/index.html)公布的水稻SSR引物信息, 合成引物。10 μL的PCR反應體系含ddH2O 5.8 μL、10×
buffer 1.0 μL、dNTP (10 mmol L–1) 0.8 μL、PrimerF 0.15 μL、PrimerR 0.15 μL、Taq DNA聚合酶(5 U μL–1) 0.1 μL、DNA 2 μL。在ABI PCR儀中進行擴增反應, 95℃預變性5 min, 94℃變性30 s, 56℃退火30 s, 72℃延伸40 s, 32個循環, 72℃延伸8 min。擴增反應完成后加入6×lodding buffer 4 μL備用。PCR產物以8%非變性聚丙烯酰胺凝膠進行電泳分析。利用熒光燈箱分析DNA條帶, 選取在親本間有明顯多態性的標記用于F2群體的分析。
1.5連鎖圖譜構建與QTL的分析
選擇覆蓋水稻全基因組的1000對SSR引物檢測WD15515與9311之間的多態性。再利用篩到的多態引物檢測WD15515/9311 F2群體中各單株的基因型, 按照QTL IciMapping軟件的要求, 將與WD15515相同的帶型記為“2”, 與9311相同的帶型記為“0”, 與F1雜種帶型相同的記為“1”, 其他或缺失帶型記為“-1”。
使用QTL IciMapping 3.0分析QTL[11-12], 使用Map功能構建遺傳圖譜, 先用“Group”命令進行連鎖分群, 再用“Ordering”命令確定各連鎖群中各標記的最佳順序, 最后以“Rippling”命令確定各標記在染色體上的排列順序及間距。將LOD值定為3.0, 作圖時以Kosambi函數將重組率轉換為遺傳距離[13]。隨后使用bip功能, 以LOD值2.5為閾值, 進行QTL掃描, 遵循McCouch等[14]原則命名QTL。
2 結果與分析
2.1SSR遺傳圖譜的構建
利用覆蓋水稻基因組的1000對SSR引物檢測親本WD15515與9311間的多態性, 共篩選到多態性引物320對。選取其中帶型清晰、差異明顯、均勻分布在12條染色體上的135對引物進行分子標記連鎖圖譜的構建。
以100個F2個體構建了包含135個SSR標記的遺傳連鎖圖譜(圖2)。該圖覆蓋水稻基因組的1194.35 cM, 每2個標記間平均距離為9.71 cM, 標記的順序與已發表的連鎖圖譜基本一致[15]。
2.2親本和F2群體的抗褐飛虱表型評價
在抗褐飛虱親本WD15515植株上取食48 h后,褐飛虱單頭雌蟲的蜜露分泌量為1.87 mg, 蟲體增重量為-0.04 mg, 表明在抗蟲植株上褐飛虱的取食受到嚴重抑制, 生長發育受阻。在感蟲親本9311植株上, 褐飛虱單頭雌蟲的蜜露分泌量達到4.94 mg, 體重增加1.88 mg, 蟲體增重比為0.86, 表明褐飛虱的取食和生長發育均正常。在F2群體中單株的抗蟲性變異較廣, 蜜露分泌量最小值為0.03 mg, 最大值為13.78 mg, 平均值為2.97 mg; F2群體蟲體增重量最小值為-0.43 mg, 最大值為2.60 mg, 平均值為0.71 mg; 蟲體增重比最小值為-0.17, 最大值為1.15, 平均值為0.32 (表1)。3種表型值在F2群體中表現為連續分布(圖3), 提示在抗性材料WD15515中可能存在多個抗性位點。相關性分析發現, 蜜露分泌量、蟲體增重和蟲體增重比在0.01水平上顯著相關(表2)。用QTL掃描分析結果中LOD值最高的分子標記RM16996分析F2群體各單株的基因型, F2群體的基因型和表型頻率分布如圖3, 從圖中可知, 基因型為純合和雜合WD15515的單株表現出較低的蜜露分泌量, 較低的增重量和增重比, 并且增重量和增重比出現了負增長的情況。
2.3F2群體中抗褐飛虱QTL分析
基于F2群體基因型和3種表型值結果, 應用QTL IciMapping 3.0軟件進行完備區間作圖分析, 共檢測到4個QTL (圖4), 分別位于水稻第2、第4、第9染色體上, 其中在第2染色體上以蜜露分泌量為指標檢測到抗性位點qBph2-1, 位于分子標記RM71–RM6911之間, LOD值為3.68, 表型貢獻率為11.08%。以蟲體增重量和增重比為指標檢測到位于RM6911–RM521之間的抗性位點qBph2-2, 其LOD值分別為3.31、4.05, 表型貢獻率分別為7.81%、9.38%。以蜜露分泌量、蟲體增重量和增重比在第4染色體上檢測到QTL qBph4位于分子標記RM16996–RM17075之間, LOD值分別為11.11、13.81、15.41, 表型貢獻率分別達到44.38%、45.24%、52.40%。表明qBph4是一個對褐飛虱取食和生長均有顯著效應的主效位點。以蜜露分泌量、蟲體增重和增重比為指標在第9染色體分子標記RM219~ RM6444之間檢測到qBph9, 其LOD值分別為2.59、4.04、3.63, 表型貢獻率分別為10.91%、12.39%、10.01% (表3)。4個QTL來源于WD15515的等位基因均能增強對褐飛虱的抗性。
2.4主效抗性位點的確認

圖2 利用9311/WD15515 F2群體構建的水稻分子標記連鎖圖譜Fig. 2 Molecular linkage map constructed by SSR markers based on 9311/WD15515 F2population

表1 親本及F2群體的性狀表現Table 1 Performance of observed traits in the parents and F2population

表2 3種抗褐飛虱表型之間的相關系數Table 2 Correlation coefficient between observed traits in the F2population
為了進一步驗證主效抗性位點qBph4, 構建了9311/WD15515//9311 BC1F1群體, 該群體共有94個單株。測定了在BC1F1各植株上褐飛虱的蜜露分泌量、蟲體增重量和增重比值3種表型值, 用標記RM16996對群體各單株進行基因型檢測, 得到
BC1F1群體的基因型和表型頻率分布圖(圖5)。結果表明, qBph4位于RM16996–RM17075之間, 這一結果和F2群體得到的結論一致。含有WD15515基因來源的雜合基因型單株表現出較低的蜜露分泌量,較低的增重量和增重比, 證明該抗性位點在雜合情況下也有較好的抗性水平。

圖3 9311/WD15515 F2群體的RM16996基因型和抗蟲性表型頻率分布圖Fig. 3 Frequency distributions of BPH-resistance phenotypes and genotypes in F2population derived from the cross of 9311/WD15515黑色a、白色b和灰色h分別表示WD15515、9311和雜合的基因型。The black a, white b, and gray h bars denote the genotypes of WD15515 homozygotes, 9311 homozygotes and heterozygotes, respectively.
3 討論
目前, 報道的褐飛虱抗性鑒定方法較多, 如水稻苗期抗性鑒定、水稻分蘗盛期抗性鑒定、褐飛虱存活率測定以及水稻分蘗期蜜露量測定等。水稻苗期集團法作為標準的褐飛虱抗性鑒定方法, 具有速度快、可以大批量篩選等優點, 但用蟲量大, 不能做F2分離群體的單株抗蟲性鑒定。劉光杰等[16]采用苗期群體篩選, 苗期單株接蟲鑒定, 分蘗期單株鑒定以及蜜露量測定等對12個水稻品種進行褐飛虱抗性鑒定表明, 水稻分蘗期蜜露分泌量測定的結果與水稻分蘗期抗性鑒定以及苗期抗蟲鑒定的結果一致。該研究是以測定褐飛虱取食水稻后, 滴落在濾紙上經茚三酮染色后蜜露所呈現面積大小評價水稻品種的抗性, 測定蜜露分泌量相對較為繁瑣。本研究中, 利用稱重法測定褐飛虱的蜜露量, 同時測定褐飛虱蟲體重量的變化和增重比值來綜合評價稻株的抗性水平, 具有更好的準確性和可重復性。我們前期研究結果表明, 這種以褐飛虱的表現來衡量水稻植株抗性, 與苗期集團法鑒定結果有高度一致性。

圖4 水稻抗褐飛虱QTL在染色體上的分布Fig. 4 Distribution of the rice brown planthopper resistance QTLs on chromosome HW: 蜜露分泌量; WB: 蟲體增重; WRB: 蟲體增重比。HW represents honeydew weight; WB represents weight increment of BPH; WRB represents BPH weight increased ratio.
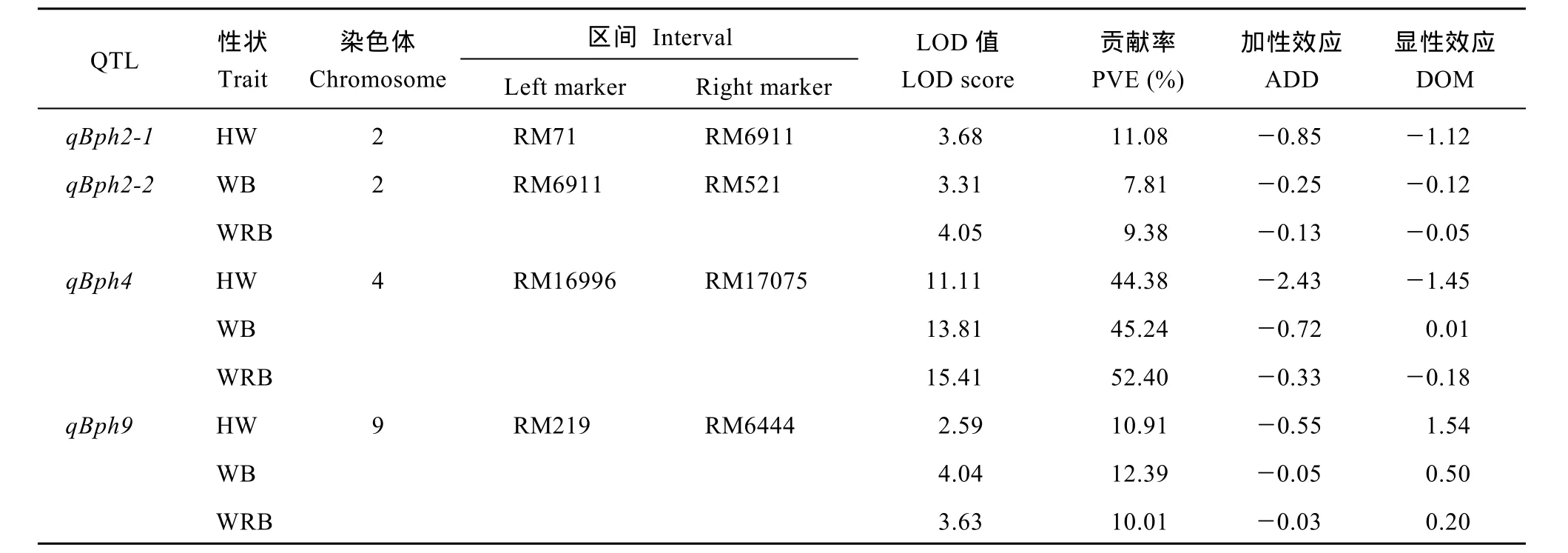
表3 水稻9311/WD15515 F2群體中抗褐飛虱QTL檢測結果Table 3 Brown planthopper resistance QTLs detected in 9311/WD15515 F2population
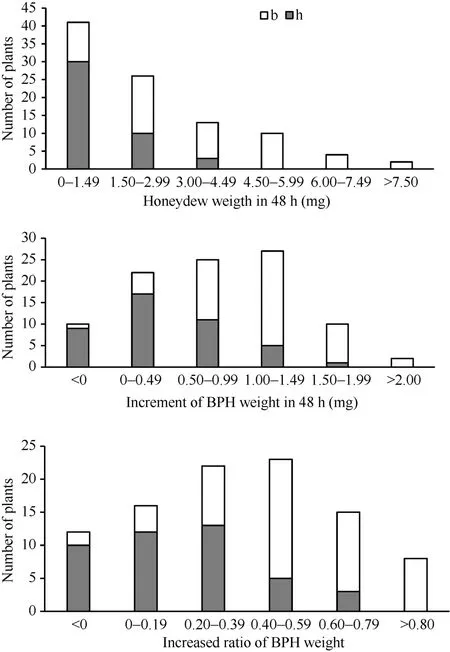
圖5 WD15515/9311//9311 BC1F1群體中抗褐飛虱表型和RM16996基因型頻率分布圖Fig. 5 Frequency distributions of BPH-resistance phenotypes and genotypes of RM16996 in the BC1F1population derived from the cross of WD15515/9311//9311圖示為SSR標記RM16996在BC1F1群體中的基因型。白色b和灰色h柱子分別表示9311和雜合的基因型。Genotypes for the SSR marker RM16996 in the BC1F1population is shown. The white b and gray h bars denote the genotypes of 9311 homozygotes and heterozygotes, respectively.
本研究測定褐飛虱在F2植株上的蜜露分泌量、蟲體增重量和增重比值作為表型值, 定位了4個抗褐飛虱QTL, 即qBph2-1、qBph2-2、qBph4和qBph9,分別位于水稻第2、第4、第9染色體上。在已報道的水稻抗褐飛虱基因中, 第2染色體上已經定位的有Bph13(t), 位于長臂末端的標記RM250~RM240之間[17], 與本研究的qBph2-1和qBph2-2相距較遠,因此這兩個位點是新的抗褐飛虱位點。迄今為止, 第4染色體上已經定位了8個褐飛虱抗性基因, 分別為Bph3、Bph6、bph12、Bph12、Bph15、Bph17、Bph20(t)、Bph27、Bph27(t)[18]。其中, Bph3、Bph12、Bph15、Bph17、Bph20(t)定位于第4染色體短臂[19-20], 與qBph4定位區間相距較遠, 而Bph6、bph12、bph18(t)、Bph27和Bph27(t)等均被定位于第4染色體長臂上, 與qBph4位置接近。bph12被定位于RFLP標記G271和R93之間比較寬的一個區間內[23]; Bph6來源于秈稻品種Swarnalata, 已被精細定位于STS標記Y19和Y9之間的25 kb區域內, 靠近bph12[24]。bph18(t)來源于普通野生稻, 被定位于標記RM273和RM6506之間[25]; Bph27來源于廣西野生稻, 被精細定位于SSR標記RM16846和RM16853之間的86.3 kb區域[26]; Bph27(t)是從Balamawee材料中新鑒定出來的顯性抗蟲基因, 被定位于InDel標記Q52和Q20之間的63 kb[27]。通過與第4染色體長臂已定位的基因位置對比分析,本研究中定位的qBph4與Bph6位置相近, 二者是否為等位或同一基因需做進一步分析。第9染色體上僅報道過Bph(t)基因[28], 位于長臂末端的RFLP標記
CDO412~RZ404之間, 與qBph9相距較遠, 因此, qBph9是一個新的抗褐飛虱基因位點。
4 結論
共定位了4個水稻抗褐飛虱QTL, 即qBph2-1、qBph2-2、qBph4、qBph9, 共解釋表型變異率超過65%, 其中qBph4的貢獻率達到44%~52%, 是一個主效位點, 該位點在雜合狀態時也有較好的抗性。因此, qBph4可應用于雜交水稻育種。其他3個效應值較小的QTL, 將在培育穩定持久抗蟲品種的育種項目中發揮作用。
References
[1] He G C, Du B, Chen R Z. Insect resistance. In: Zhang Q F, Wingeds R A, eds. Genetics and Genomics of Rice. New York: Springer, 2013. p 399
[2] 翟保平. 稻飛虱: 國際視野下的中國問題. 應用昆蟲學報, 2011, 48: 1184–1193
Zhai B P. Rice planthoppers: a China problem under the international perspectives. Chin J Appl Entomol, 2011, 48: 1184–1193 (in Chinese with English abstract).
[3] Cheng X Y, Zhu L L, He G C. Towards understanding of molecular interactions between rice and the brown planthopper. Mol Plant, 2013, 6: 621–634
[4] Du B, Zhang W L, Liu B F, Hu J, Wei Z, Shi Z Y, He R F, Zhu L L, Chen R Z, Han B, He G C. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc Natl Acad Sci USA, 2009, 106: 22163–22168
[5] Tamura Y, Hattori M, Yoshioka H, Yoshioka M, Takahashi A, Wu J Z, Sentoku N, Yasui H. Map-based cloning and characterization of a brown planthopper resistance gene BPH26 from Oryza sativa L. ssp. indica cultivar ADR52. Sci Rep, 2014, 4: 5872
[6] Liu Y, Wu H, Chen H, Wu H, Chen H, Liu Y L, He J, Kang H Y, Sun Z G, Pan G, Wang Q, Hu J L, Zhou F, Zhou K N, Zheng X M, Ren Y L, Chen L M, Wang Y H, Zhao Z G, Lin Q B, Wu F Q, Zhang X, Guo X P, Cheng X N, Jiang L, Wu C Y, Wang H Y, Wan J M. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat Biotechnol, 2015, 33: 301–305
[7] Cohen M B, Alam S N, Medina E B, Bernal C C. Brown planthopper, Nilaparvatalugens, resistance in rice cultivar IR64: mechanism and role in successful N. lugens management in Central Luzon, Philippines. Entomol Exp Appl, 1997, 85: 221–229
[8] Huang Z, He G C, Shu L H, Li X H, Zhang Q F. Identification and mapping of two brown planthopper resistance genes in rice. Theor Appl Genet, 2001, 102: 929–934
[9] Xu X F, Mei H W, Luo L J, Chen X N, Li Z K. RFLP-facilitated investigation of the quantitative resistance of rice to brown planthopper (Nilaparvata lugens). Theor Appl Genet, 2002, 104: 248–253
[10] 蘇昌潮, 程遐年, 翟虎渠, 萬建民. 利用回交重組自交群體檢測水稻抗褐飛虱數量性狀基因座. 遺傳學報, 2002, 29: 332–338
Su C C, Cheng X N, Zhai H Q, Wan J M. Detection and analysis of QTL for resistance to the brown planthopper, Nilaparvata lugens (Stal), in rice (Oryza sativa L.), using backcross inbred lines. Acta Genet Sin, 2002, 29: 332–338 (in Chinese with English abstract)
[11] 王建康. 數量性狀基因的完備區間作圖方法. 作物學報, 2009, 35: 239–245
Wang J K. Inclusive composite interval mapping of quantitative trait genes. Acta Agron Sin, 2009, 35: 239–245 (in Chinese with English abstract)
[12] 李慧慧, 張魯燕, 王建康. 數量性狀基因定位研究中若干常見問題的分析與解答. 作物學報, 2010, 36: 918–931
Li H H, Zhang L Y, Wang J K. Analysis and answers to frequently asked questions in quantitative trait locus mapping. Acta Agron Sin, 2010, 36: 918–931 (in Chinese with English abstract).
[13] Kosambi D D. The estimation of map distances from recombination values. Ann Eugenics, 1994, 12: 172–175
[14] McCouch S R, Cho Y G, Yano M, Paul E, Blinstrub M, Morishima H, Kinoshita T. Report on QTL nomenclature. Rice Genet Newsl, 1997, 14(11): 11-13
[15] McCouch S R, Teytelman L, Xu Y, Lobos K B, Clare K, Walton M, Fu B Y, Maghirang R, Li Z K, Xing Y Z. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res, 2002, 9(6): 199–207
[16] 劉光杰, 鄭宜才, 桂麗琴, 沈君輝. 水稻品種抗稻飛虱鑒定方法的比較研究. 浙江農業學報, 1999, 11: 306–310
Liu G J, Zheng Y C, Gui L Q, Shen J H. A comparative study of the rice planthopper resistance identification method. Zhejiang Agric Sci, 1999, 11: 306–310 (in Chinese)
[17] 劉國慶, 顏輝煌, 傅強, 錢前, 張志濤, 翟文學, 朱立煌. 栽培稻的緊穗野生稻抗褐風虱主效基因的遺傳定位. 科學通報, 2001, 46: 738–742
Liu G Q, Yan H H, Fu Q, Qian Q, Zhang Z T, Zhai W X, Zhu L H. Mapping of a new gene for brown planthopper resistance in cultivated rice introgressed from Oryza eichingeri. Chin Sci Bull, 2001, 46: 1495–1462 (in Chinese)
[18] Ali M P, Chowdhury T R. Tagging and mapping of genes and QTLs of Nilaparvatalugens resistance in rice. Euphytica, 2014, 195: 1–30
[19] Qiu Y F, Guo J P, Jing S L, Zhu L L, He G C. Development and characterization of japonica rice lines carrying the brown planthopper-resistance genes BPH12 and BPH6. Theor Appl Genet, 2012, 124: 485–494
[20] Yang H Y, You A Q, Yang Z F, Zhang F, He R F, Zhu L L, He G C. High-resolution genetic mapping at the Bph15 locus for brown planthopper resistance in rice (Oryza sativa L.). Theor Appl Genet, 2004, 110: 182–191
[21] Sun L H, Su C C, Wang C M, Zhai H Q, Wan J M. Mapping of a major resistance gene to the brown planthopper in the rice cultivar Rathu Heenati. Breed Sci, 2005, 55: 391–396
[22] Rahman M L, Jiang W, Chu S H, Qiao Y, Ham T H, Woo M O, Lee J, Khanam M S, Chin J H, Jeung J U. High-resolution mapping of two rice brown planthopper resistance genes, Bph20(t) and Bph21(t), originating from Oryza minuta. Theor Appl Genet, 2009, 119: 1237–1246
[23] Hirabayashi H, Ogawa T. Identification and utilization of DNA markers linked to genes for reistance to brown planthopper (BPH) in rice. Adv Breed (Japan), 1999, 41: 71–74
[24] Qiu Y F, Guo J P, Jing S L, Zhu L L, He G C. High-resolution mapping of the brown planthopper resistance gene Bph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isogenic backgrounds. Theor Appl Genet, 2010, 121: 1601–1611
[25] Li R B, Li L S, Wei S M, Wei Y P, Chen Y Z, Bai D L, Yang L, Huang F K, Lu W L, Zhang X J, Li X Y, Yang X Q, Wei Y W. The evaluation and utilization of new genes for brown planthopper resistance in common wild rice (Oryza rufipogon Griff.). Mol Plant Breed, 2006, 4: 365–371
[26] Huang D, Qiu Y, Zhang Y, Huang F, Meng J, Wei S, Li R, Chen B. Fine mapping and characterization of BPH27, a brown planthopper resistance gene from wild rice (Oryza rufipogon Griff.). Theor Appl Genet, 2013, 126: 219–229
[27] He J, Liu Y, Liu Y Q, Liu Y L, Jiang L, Wu H, Kang H Y, Liu S J, Chen L M, Liu X, Cheng X N, Wan J M. High-resolution mapping of brown planthopper (BPH) resistance gene Bph27(t) in rice (Oryza sativa L.). Mol Breed, 2013, 31: 549–557
[28] Mei M, Zhuang C, Wan R, Wu J, Hu W, Kochert G. Genetic analysis and tagging of gene for brown planthopper resistance in indica rice. In: Rice Genetics III. Proceedings of the Third International Rice Genetics Symposium. IRRI, Los Ba?os, Philippines, 1996. pp 16–20
URL: http://www.cnki.net/kcms/detail/11.1809.S.20151207.1041.014.html
Analysis of QTLs for Brown Planthopper Resistance in Indica Rice WD15515
DENG Zhao**, SHI Shao-Jie**, WANG Hui-Ying, SHANG-GUAN Xin-Xin, LIU Bing-Fang, JING Sheng-Li, DU Bo, CHEN Rong-Zhi, ZHU Li-Li, and HE Guang-Cun*
State Key Laboratory of Hybrid Rice, College of Life Sciences, Wuhan University, Wuhan 430072, China
Abstract:Brown planthopper (BPH) is one of the most destructive insect pests of rice. WD15515 is an indica germplasm highly resistant to BPH. An F2population was developed from the cross between 9311 and WD15515. One hundred F2plants were genotyped by using SSR markers and evaluated for BPH-resistance by measuring the honeydew weight secreted by BPH (HW), the body weight increment (WB) and the body weight increment ratio (WRB). A total of four QTLs for BPH resistance were identified. The qBph2-1, based on HW (honeydew weight), was detected between RM71 and RM6911 on chromosome 2, with LOD score of 3.68 and explaining the 11.08% of phenotypic variation. The qBph2-2, based on both WB and WRB, was mapped between RM6911 and RM521 on chromosome 2, with LOD score of 3.31 and 4.05 and explaining 7.81% and 9.38% of the phenotypic variation, respectively. The qBph4, based on HW, WB and WRB, was detected between RM16996 and RM17075 on chromosome 4, with LOD score of 11.11, 13.81, and 15.41 and explaining 44.38%, 45.24%, and 52.40% of the phenotypic variation, respectively. The qBph9, based on HW, WB, and WRB, was detected on chromosome 9 between RM219 and RM6444, with LOD score of 2.59, 4.04, and 3.63. This locus explained 10.91%, 12.39%, and 10.01% of the phenotypic variation in this population, respectively. qBph4 is a major gene for BPH-resistance. This result provides the new resources for BPH-resistance breeding.
Keywords:Rice; Resistance; Brown planthopper; QTL; Genetic map
收稿日期Received(): 2015-07-08; Accepted(接受日期): 2015-11-20; Published online(網絡出版日期): 2015-12-07.
通訊作者*(Corresponding author): 何光存, E-mail: gche@whu.edu.cn, Tel: 027-68752384
DOI:10.3724/SP.J.1006.2016.00353
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
