鐵皮石斛DORCA基因的克隆及表達分析
2016-03-23 02:25:46王潔瓊宋彩霞袁秀云
西北植物學報 2016年1期
崔 波,王潔瓊,宋彩霞,袁秀云,
蔣素華1,梁 芳1,劉 佳2,葉永忠2*
(1 鄭州師范學院,鄭州 450044;2 河南農業大學 生命科學學院,鄭州 450002)
?
鐵皮石斛DORCA基因的克隆及表達分析
崔波1,2,王潔瓊2,宋彩霞2,袁秀云1,
蔣素華1,梁芳1,劉佳2,葉永忠2*
(1 鄭州師范學院,鄭州 450044;2 河南農業大學 生命科學學院,鄭州 450002)
摘要:為了研究鐵皮石斛的光合特性,根據Rubisco活化酶(RCA)基因的保守序列設計兼并引物,采用RT-PCR和RACE方法從鐵皮石斛葉中分離到一個RCA基因,命名為DORCA(GenBank登錄號KT205841)。DORCA基因cDNA全長為1 724 bp,包含1 317 bp開放閱讀框(ORF),編碼438個氨基酸。系統進化分析顯示,DORCA與馬蹄蓮ZaRCA(AAK25801.1)親緣關系最近。生物信息學分析表明,DORCA與其他植物的RCA蛋白具有較高一致性,屬于RCA的β亞基。DORCA蛋白具有定位于葉綠體的N端轉運肽,2個保守的ATP結合結構域和多個磷酸化位點。實時熒光定量PCR(qRT-PCR)分析表明,DORCA基因在莖、葉中表達;在自然光周期條件下,DORCA基因在8:30時表達量最高,20:30時表達量最低,具有明顯的光誘導表達特性。
關鍵詞:鐵皮石斛;Rubisco活化酶;分子特征;表達特性
鐵皮石斛(DendrobiumofficinaleKimura et Migo)為蘭科(Orchidaceae)多年生附生草本植物,是石斛屬(Dendrobium)藥用植物中珍稀名貴的品種,在中國主要分布在浙江、云南、四川、福建、廣西、安徽等地。鐵皮石斛富含多糖、生物堿、菲類化合物等藥用成分,在提高人體免疫力、抗衰老、抗腫瘤、降血糖等方面具有顯著的功效。
近年來,光合機理研究日益受到研究者的重視,其中Rubisco(核酮糖1,5-二磷酸羧化酶/加氧酶)活性的調節機制是一個重要的研究領域。Rubisco是所有光合生物進行光合碳同化的關鍵酶,它能催化RuBP(1,5-二磷酸核酮糖)的羧化和加氧反應,并調節二者之間的關系,這2個反應的比值決定了植物的碳同化效率[1]。已有研究表明,雖然Rubisco含量占葉片可溶性蛋白的一半,但它的效率卻很低[2]。Rubisco只有處于活化態時,才能催化CO2固定。因此,提高Rubisco利用效率和活力是提高光合作用的重要途徑[3]。在植物體中Rubisco活性受Rubisco活化酶(rubisco activase,RCA)的調節和控制,RCA是一種核編碼的可溶性葉綠體蛋白,廣泛存在于光合生物中,具有ATPase活性,屬于AAA+蛋白家族成員[4]。RCA的主要功能是活化Rubisco,在適溫條件下,RCA利用光合磷酸化產生的ATP使Rubisco迅速與生理濃度CO2和Mg2+結合形成ECM復合體(酶-CO2-Mg)而達到最大的活化程度,解除磷酸糖對Rubisco活性的抑制作用,完成對Rubisco的活化[5]。
目前關于RCA的研究主要集中在糧食作物如水稻[6]、小麥[7]、玉米[8]和觀賞性園藝植物如彩葉草[9]、羽衣甘藍[10]等。鐵皮石斛作為名貴中草藥,因獨特的保健功能使其市場需求逐年增加,但是生長卻比較緩慢。因此,促進鐵皮石斛的生長,提高生產效率,是目前鐵皮石斛產業發展的當務之急。為了更好地研究鐵皮石斛的光合特性,本研究采用RACE法克隆了RCA基因,并對其分子特性和表達模式進行了分析,為進一步研究鐵皮石斛的光合作用奠定了基礎。
1材料和方法
1.1植物材料
實驗材料為1年生鐵皮石斛(Dendrobiumofficinale),來源于鄭州師范學院蘭花工程技術研究中心。選取生長一致的幼苗,從當晚20:30開始每隔4 h取樣1次,到次日20:30結束,且采相同部位的葉片,保存于-80 ℃冰箱,用于RNA提取。
1.2方法
1.2.1RNA提取和cDNA合成采用多糖多酚植物總RNA提取試劑盒(Tiangen公司)提取實驗材料總RNA,用1%瓊脂糖凝膠電泳鑒定RNA完整性;用DU800核酸蛋白分析儀(美國Beckman)測定其A260、A280值,計算RNA濃度和純度。用M-MLV反轉錄酶(TaKaRa公司)以提取的總RNA為模板,合成單鏈cDNA,具體方法參照試劑盒的說明書。
1.2.2鐵皮石斛RCA基因全長cDNA克隆利用生物軟件DNAMAN和Primer 5.0,根據已有植物RCA序列的保守區,設計1對簡并引物,上游引物RCA-F1(5′-GAYGACCAGCAGGACATAACCA-3′)和下游引物RCA-R1(5′-TTGTTGACGGTGTACTGGGTG-3′),以葉cDNA為模板,擴增鐵皮石斛RCA基因的保守片段。PCR反應體系為20 μL,包含10×PCR緩沖液2.0 μL,dNTP Mix 2.0 μL,上下游引物(10 μmol/L)各1.0 μL,cDNA模板 1.0 μL,rTaq酶(5 U/μL)0.2 μL,加滅菌雙蒸水至總體積20 μL。反應程序為94 ℃預變性4 min;94 ℃變性40 s,56 ℃復性40 s,72 ℃延伸40 min,35個循環;最后72 ℃延伸20 min。PCR產物經1%瓊脂糖凝膠電泳檢測后利用凝膠試劑盒(Tiangen公司)回收目的片段,連接pGEM-T Easy載體,轉化E.coliDH5α挑選陽性克隆送至上海英駿生物技術公司測序。
根據上步所得的RCA基因片段序列,設計1對3′端特異引物GSP1(5′-TGTGCTGCCTCTTCATCAATG-3′)和GSP2(5′-ACCACCCAGTACACCGTCAA-3′),1對5′端特異引物GPS3(5′-ATACTGACGAAGACCTTGGCTGAT-3′)和GPS4(5′-CACCCTTTCCTCTGGTTATGTCCT-3′),按Clontech公司SMARTer RACE cDNA Amplification Kit操作說明進行反應,回收擴增產物,連接到pGEM-T Easy載體上,轉入大腸桿菌 DH5α菌株,鑒定后進行測序。
1.2.3鐵皮石斛RCA基因生物信息學分析用DNAMAN軟件分析拼接cDNA測序序列,得到正確的鐵皮石斛RCA基因序列。鐵皮石斛RCA蛋白的分子特征主要采用生物信息學方法進行分析。Blast搜索分析、ORF finding、核苷酸翻譯以及蛋白保守結構域均在NCBI(http://www.ncbi.nlm.nih.gov/)網站上進行;不同RCA蛋白序列的多重比對利用ClustalX和DNAMAN軟件生成比對結果,再利用MEGA 5.0進行系統進化樹構建;應用ExPASy、SMART、Net Phos.2.0 server、TMPRED、TargetP 1.1 server等綜合軟件分析包預測RCA蛋白的基本性質、結構特點、磷酸化位點、跨膜結構域和亞細胞定位等。
1.2.4鐵皮石斛RCA基因表達分析利用Primer 5.0設計鐵皮石斛RCA基因實時熒光定量PCR引物qRT-RCAF(5′-TGTCAACCGAGTTCCACTTACC-3′)和qRT-RCAR(5′-TCGAGGTCAGCAGCCAAGA-3′)。以鐵皮石斛的EF1α作為內參基因[11],引物為EF1αF(5′-TCAGGCTGACTGTGCTGTCCT-3′)和EF1αR(5′-GTGGTGGCGTCCATCTTGTT-3′)。分別提取鐵皮石斛不同組織部位,不同時間點的總RNA,使用PrimeScript RT reagent Kit with gDNA Eraser(TaKaRa)反轉錄合成單鏈cDNA。實時熒光定量PCR反應體系和程序參照寶生物工程(大連)有限公司SYBR PremixExTaqTMⅡ(Tli RNaseH Plus)試劑盒說明書。反應體系為20 μL,所用儀器為Eppendorf Mastercycler ep realplex 2。反應程序為95 ℃預變性15 s,58 ℃變性15 s,72 ℃退火15 s,40個循環。每樣品重復3次,采用公式2-ΔΔCt法計算基因相對表達量。
2結果與分析
2.1鐵皮石斛RCA基因全長cDNA克隆
以鐵皮石斛葉片cDNA為模板,擴增出1條長度為503 bp的特異保守片段(圖1),測序并在NCBI上Blast分析,該片段與多種植物的RCA序列有較高一致性(80%以上),證明該片段為RCA片段。根據該片段設計RACE引物,利用3′端引物GSP1和GSP2擴增得到3′端目的片段,用5′端引物GPS3和GPS4擴增得到5′端目的片段(圖2)。將特異保守片段、3′-RACE和5′-RACE序列拼接獲得鐵皮石斛RCA的cDNA,命名為DORCA,全長為1 724 bp,利用NCBI在線工具ORF finder確定該基因的ORF長1 317 bp,編碼438個氨基酸,包含起始密碼子和終止密碼子。5′端非編碼區為81 bp,3′端非編碼區為325 bp。
2.2DORCA蛋白序列的理化性質
DORCA基因編碼一個438 aa的前體蛋白,蛋白的分子量為48.25 kD,理論等電位點為6.19。應用在線分析軟件TargetP 1.1 Server對DORCA氨基酸序列進一步分析發現,此氨基酸序列的前57個氨基酸殘基為葉綠體轉運肽(圖3),因此成熟蛋白分子量為41.1 kD,推測以β(41~43 kD)亞基存在。TMPRED對氨基酸序列進行跨膜結構預測表明在N端區域內(第17~33位aa)具有跨膜結構。NetPhos 2.0 Server預測表明,DORCA蛋白有8個Ser磷酸化位點,4個Thr磷酸化位點和5個Tyr磷酸化位點,這表明蛋白翻譯后修飾對DORCA蛋白功能的實現起重要作用。SMART和CDS保守結構域搜索表明,DORCA蛋白含有一個ATP酶的高度保守的P-loop NTPase Superfamily結構域(圖3),其中在162~309 aa位點范圍內包含AAA+蛋白家族折疊區,含有2個保守的ATP結合基序(motif),分別為“WGGKGQGKS”(A區)和“GKMCCLFINDLD”(B區)。

M.Marker;1.PCR產物
M.Marker;1.5′-RACE;2.3′-RACE
2.3不同植物RCA基因系統進化分析
用NCBI上的Blastp程序和DNAMAN多序列對比分析表明,DORCA蛋白序列與許多植物的RCA具有較高的一致性,其中與油棕(Elaeisguineensis)的一致性最高,達87%。將DORCA基因氨基酸序列與GenBank蛋白質數據庫進行氨基酸序列同源性比對,發現具有較高一致性的植物包括:海棗(Phoenixdactylifera)、荷花(Nelumbonu-cifera)、葡萄(Vitisvinifera)、可可(Theobromacacao)、麥冬(Ophiopogonjaponicus)、馬蹄蓮(Zantedeschiaaethiopics)和大豆(Glycinemax)等,與這些基因的一致性都在80%以上。在MEGA 5.0軟件上運用Custal X對這些氨基酸序列進行系統進化樹構建(圖4),結果表明,DORCA與單子葉植物馬蹄蓮(Zantedeschiaaethiopics)的親緣關系最近。多序列比對也表明DORCA蛋白的C末端沒有受硫氧化蛋白f (Trx-f)的氧化還原作用所調節的2個Cys殘基(圖5),表明DORCA蛋白屬于β亞基,這與DORCA蛋白分子量為41.1 kD屬于β亞基的分析一致。
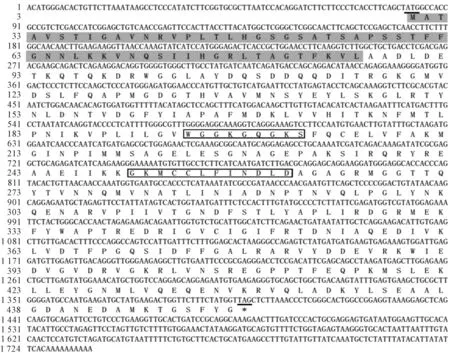
起始密碼子ATG用下劃線表示,終止子TAG用星號和下劃線表示;灰色背景表示定位于

方框代表本研究的DORCA在系統進化樹中的位置;圖中分支上的數字表示Bootstrap驗證中

α亞基C末端的特異C-extension結構用方框表示;2個推測的ATP結合位點P-環序列用方框A、B表示;
2.4DORCA表達模式分析
為了研究DORCA基因在鐵皮石斛組織中的表達模式,采用實時熒光定量RT-PCR對鐵皮石斛的根、莖、葉進行分析,結果顯示,DORCA主要在綠色組織中表達,其中葉片中的表達量最高,在莖中相對表達量較少,而在根中幾乎檢測不到(圖6)。本研究得出的結論與玉米[8]、水稻[6]等RCA只在綠色組織中特異性表達相一致。研究表明,大量光合作用相關基因受光照和生物鐘節律的調控,為了檢測DORCA是否受光調節,選取生長在每天12 h自然光照(6:00~18:00)和12 h黑暗條件(18:00~6:00)下的鐵皮石斛幼苗,提取相應時間點的葉RNA,進行qRT-PCR表達分析。結果顯示(圖6),在自然光周期條件下,DORCA表達量變化呈現出明顯的生物鐘特性。黑暗周期過程中,DORCA的相對表達量逐漸增加,至上午8:30時達到最大值,隨后逐漸降低,到晚上20:30降至最低。另外結果還顯示,DORCA基因表達水平較高的時間段主要集中在4:30~12:30。DORCA受光調節的特點與玉米[8]、蘋果[11]等多種植物RCA受光調節并具有晝夜節奏的表達特性相同。
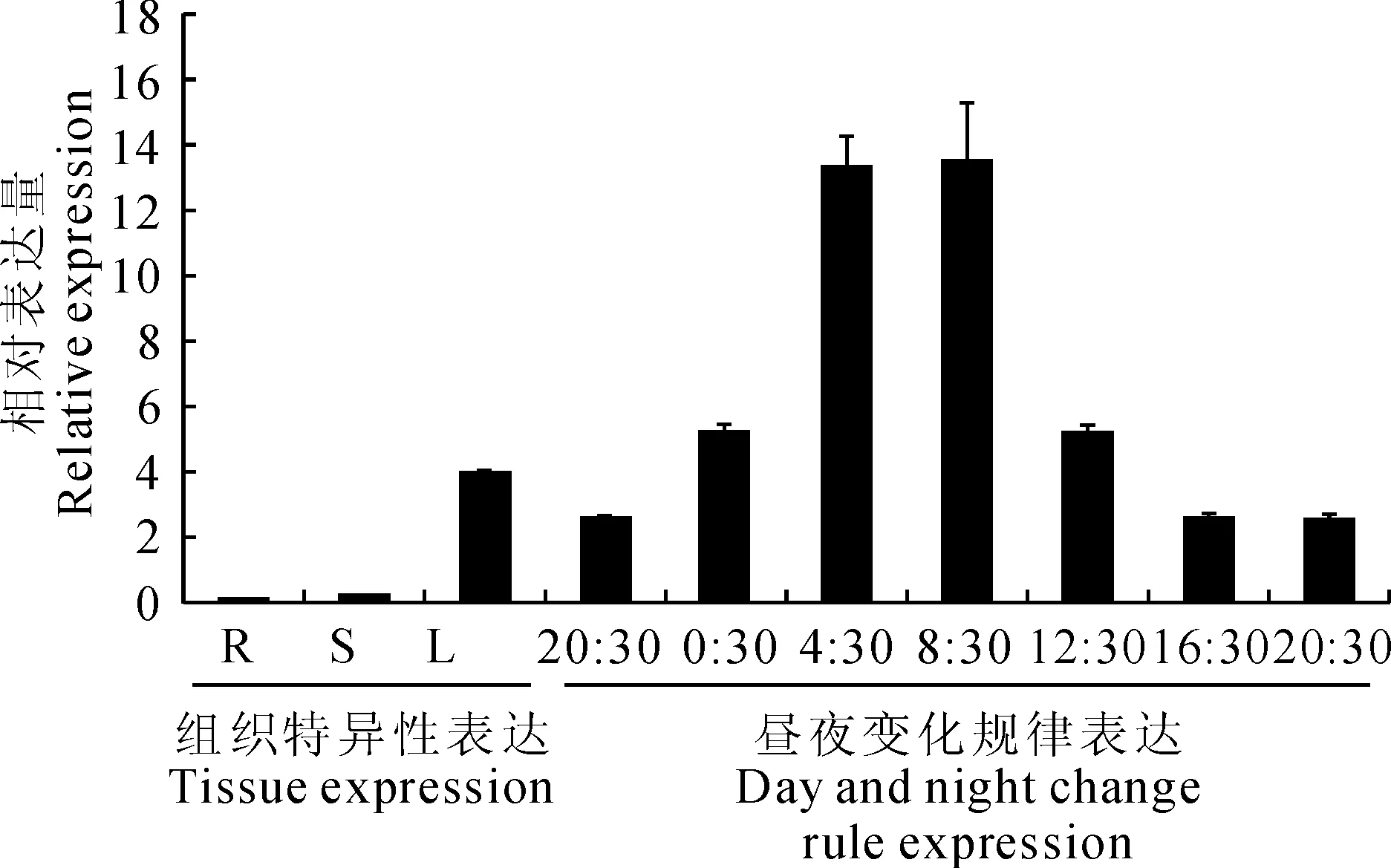
R、S和L分別代表根、莖、葉
3討論
在植物光合作用的研究中,關于 Rubisco活化酶活性調節機制一直是一個重要的研究領域。Rubisco活化酶是維持和調節Rubisco活性所必需的,在植物體中只有Rubisco活化酶存在時Rubisco才能保持較高的催化活性[12]。在本研究中利用RT-PCR和RACE技術從鐵皮石斛中擴增出了一個RCA基因的全長cDNA序列DORCA(GenBank登錄號KT205841)。并利用生物信息學方法對DORCA蛋白的分子特征進行分析和預測。結果表明DORCA基因cDNA全長1 724 bp,編碼438個氨基酸的前體蛋白,N端具有57個氨基酸的葉綠體轉運肽,是核基因編碼的葉綠體蛋白。DORCA編碼的成熟蛋白具有RCA蛋白都含有的2個保守的ATP結合基序,屬于AAA+蛋白家族。系統進化樹顯示DORCA基因的氨基酸序列與其他已知植物RCA具有高度一致性,且與馬蹄蓮親緣關系最近屬于同一支。
大量研究表明,植物RCA基因具有組織表達特異性。Liu等[13]研究發現擬南芥RCA基因只在綠色組織如葉、莖、角果中表達。Watillon等[14]研究發現蘋果RCA基因同樣在葉片中能大量檢測到,但在根和莖中幾乎檢測不到。本研究利用實時熒光定量RT-PCR方法得出的檢測結果與前人結果相似[13-14],且與Rubisco僅在綠色組織中特異表達相一致。同時多數植物RCA基因的表達還受到光的誘導及生物鐘的調控[15-16],本研究中同樣發現鐵皮石斛RCA基因晝夜表達變化節奏性,開始時表達水平最低,然后表達量逐漸增加,至上午8:30時表達量達到最大值,隨后逐漸降低,表現出了生物鐘特性。
不同植物RCA亞基的種類和數量是不同的,如小麥[7]、水稻[6]、擬南芥[17]、大豆[18]、菠菜[17]、棉花[19]等植物均含有α和β亞基,而有些植物如黃瓜[20]、玉米[8]、煙草[15]等卻只表達β亞基。植物RCA的α亞基大小范圍在45~47 kD,而且只有α亞基的C末端具有調節硫氧還蛋白f (Trx-f)的2個Cys殘基,β亞基的大小為41~44 kD,C末端沒有保守結構[16]。DORCA蛋白經多重序列對比表明,其C端沒有α亞基所具有的2個Cys殘基,并且成熟蛋白大小為41.1 kD,說明屬于β亞基形式。研究表明無論何種形式的亞基,來源于同種或是不同種的植物抗體之間都有可能發生交叉反應[18]。另外,RCA基因的表達方式也各不相同,有的是由一個RCA基因通過選擇性剪接編碼2個亞基,有的是不同基因編碼同一個亞基[21]。因此,在鐵皮石斛中是否還具有α亞基形式,以及其β亞基是否是由一個或者多個基因編碼的,還需要利用Southern blotting及Western blotting等技術進一步進行分析確定。
蘇文華等[22]研究表明,鐵皮石斛葉片具有兼性CAM植物的特點,晴天呈CAM途徑、陰雨天呈C3途徑,多云天氣在上述兩種途徑之間變化。本試驗關于DORCA基因的分子克隆與表達特性的研究,為進一步研究鐵皮石斛復雜的光合作用奠定了基礎。目前正在構建DORCA基因的超表達載體進行遺傳轉化,試圖進一步解釋其調控的分子機制。
參考文獻:
[1]MAKINO A,MAE T,OHIRA K.Purification and storage of ribulose 1,5-bisphosphate carboxylase from rice leaves[J].PlantandCellPhysiology,1983,24(6):1 169-1 173.
[2]MANN C C.Genetic engineers aim to soup up crop photosynthesis[J].Science,1999,283(5 400):314-316.
[3]PORTIS J R A R.Rubisco activase-Rubisco’s catalytic chaperone[J].PhotosynthesisResearch,2003,75(1):11-27.
[4]張國,王瑋,鄒琦.Rubisco活化酶的分子生物學[J].植物生理學通訊,2004,40(5):633-637.
ZHANG G,WANG W,ZOU Q.Molecular biology of Rubisco activase[J].PlantPhysio.Commu.,2004,40(5):633-637.
[5]LILLEY R M,PORTIS A R J R.Activation of ribulose-1,5-bisphosphate carboxylase/oxygenase(rubisco) by rubisco activase[J].PlantPhysiol.,1990,94:245-247.
[6]TO K Y,SUEN D F,CHEN S C G.Molecular characterization of ribulose-1,5-bisphosphate carboxylase/oxygenase activase in rice leaves[J].Planta,1999,209(1):66-76.
[7]LAW R D,CRAFTS-BRANDNER S J.High temperature stress increases the expression of wheat leaf ribulose-1,5-bisphosphate carboxylase/oxygenase activase protein[J].ArchivesofBiochemistryandBiophysics,2001,386(2):261-267.
[8]AYALA-OCHOA A,VARGAS-SUAREZ M,LOZA-TAVERA H,etal.In maize,two distinct ribulose 1,5-bisphosphate carboxylase/oxygenase activase transcripts have different day/night patterns of expression[J].Biochimie,2004,86(7):439-449.
[9]祝欽瀧,謝先榮,眭順照,等.彩葉草Rubisco活化酶基因SsRCA的分子特征及其表達模式[J].西北植物學報,2011,31(6):1 097-1 104.
ZHU Q L,XIE X R,GUI X ZH,etal.Molecular characteristics and expression patterns of a novel Rubisco activase gene,SsRCA,fromSolenostemonscutellarioides(L.) Codd[J].ActaBot.Boreal.-Occident.Sin.,2011,31(6):1 097-1 104.
[10]朱明庫,陳國平,周爽,等.羽衣甘藍Rubisco活化酶基因BoRCA的克隆,生物信息學及表達分析[J].云南大學學報(自然科學版),2013,35(6):857-865.
ZHU M K,CHEN G P,ZHOU SH,etal.Cloning,bioinformatics and expression analysis of Rubisco activase geneBoRCAfromBrassicaoleracea[J].JournalofYunanUniversity(Nat.Sci.Edi.),2013,35(6):857-865.
[11]RUNDLE S J,ZIELINSKI R E.Organization and expression of two tandemly oriented genes encoding ribulosebisphosphate carboxylase/oxygenase activase in barley[J].JournalofBiologicalChemistry,1991,266(8):4 677-4 685.
[12]張崗,趙明明,張大為,等.鐵皮石斛實時定量PCR內參基因的篩選[J].中國藥學雜志,2013,19:15.
ZHANG G,ZHAO M M,ZHANG D W,etal.Reference gene selection for real-time quantitative PCR analysis ofDendrobiumofficinale[J].Chin.Pharm.J.,2013,48(19):1 664-1 668.
[13]LIU Z,TAUB C C,MCCLUNG C R.Identification of anArabidopsisthalianaribulose-1,5-bisphosphate carboxylase/oxygenase activase (RCA) minimal promoter regulated by light and the circadian clock[J].PlantPhysiology,1996,112(1):43-51.
[14]WATILLON B,KETTMANN R,BOXUS P,etal.Developmental and circadian pattern of rubisco activase mRNA accumulation in apple plants[J].PlantMolecularBiology,1993,23(3):501-509.
[15]QIAN J.,RODERMEL S.R.Ribulose-l,5-bisphosphate carboxylase/oxygenase activase cDNAs fromNicotianatabacum[J].PlantPhysiol.,1993,102:683-684.
[16]ROESLER K R,OGREN W L.Primary structure ofChlamydomonasreinhardtiiribulose 1,5-bisphosphate carboxylase/oxygenase activase and evidence for a single polypeptide[J].PlantPhysiology,1990,94(4):1 837-1 841.
[17]WERNEKE J M,CHATFIELD J M,OGREN W L.Alternative mRNA splicing generates the two ribulosebisphosphate carboxylase/oxygenase activase polypeptides in spinach andArabidopsis[J].ThePlantCell,1989,1(8):815-825.
[18]YIN Z,MENG F,SONG H,etal.Expression quantitative trait loci analysis of two genes encoding rubisco activase in soybean[J].PlantPhysiology,2010,152(3):1 625-1 637.
[19]SALVUCCI M E,VAN DE LOO F J,STECHER D.Two isoforms of Rubisco activase in cotton,the products of separate genes not alternative splicing[J].Planta,2003,216(5):736-744.
[20]ZHANG N,KALLIS R P,EWY R G,etal.Light modulation of Rubisco inArabidopsisrequires a capacity for redox regulation of the larger Rubisco activase isoform[J].ProceedingsoftheNationalAcademyofSciences,2002,99(5):3 330-3 334.
[21]李海霞,王真梅,曾漢來.植物 Rubisco 活化酶的研究進展[J].植物生理學報,2010,46(11):1 092-1 100.
LI H X,WANG Z M,ZENG H L.The research progresses in Rubisco activase in plant[J].PlantPhysio.Commu.,2010,46(11):1 092-1 100.
[22]蘇文華,張光飛.鐵皮石斛葉片光合作用的碳代謝途徑[J].植物生態學報,2003,27(5):631-637.
SU W H,ZHANG G F.The photosynthesis pathway in leaves ofDendrobiumofficinale[J].ActaPhytoecologicaSinica,2003,27(5):631-637.
(編輯:宋亞珍)
Cloning and Expression Analysis ofDORCAGene fromDendrobiumofficinale
CUI Bo1,2,WANG Jieqiong2,SONG Caixia2,YUAN Xiuyun1,JIANG Suhua1,LIANG Fang1,LIU Jia2,YE Yongzhong2*
(1 Zhengzhou Normal College,Zhengzhou 450044,China;2 College of Life Sciences,Henan Agricultural University,Zhengzhou 450002,China)
Abstract:To study the photosynthesis of Dendrobium officinale,we isolated a full-length cDNA of Rubisco activase DORCA(Accession No.KT205841) from this plant by the method of RT-PCR and RACE using degenerate primers designed according to the conserved region of RCA.The full-length cDNA of DORCA was 1 724 bp,with an ORF of 1 317 bp encoding 438 putative amino acids.Phylogenetic analysis showed that DORCA closed to ZaRCA(AAK25801.1).Bioinformatics analysis showed that DORCA belonged to β isoform of RCA protein and had high identities with other plant RCAs.The DORCA contained a predicted N-terminal transit peptide to chloroplast,two high conserved ATP-binding domains and multiple phosphorylation sites.The real-time quantitative PCR(qRT-PCR) results showed that DORCA was only detected in green tissues.In the 12 h dark and 12 h natural light photoperiods,maximal and minimal DORCA mRNA expression levels were detected at 8:30 and 20:30,respectively,which indicated that DORCA has the obvious characteristics of light-induced expression.
Key words:Dendrobium officinale;Rubisco activase;molecular characteristics;expression patterns
中圖分類號:Q785;Q786
文獻標志碼:A
作者簡介:崔波(1962-),男,博士,教授,主要從事花卉分子生物學研究。E-mail:laocuibo@163.com*通信作者:葉永忠,教授,博士生導師,主要從事植物生態學研究。E-mail:yeyzh@163.com
基金項目:河南省科技攻關項目(092102110128);鄭州市科技攻關項目(141PPTGG420)
收稿日期:2015-09-25;修改稿收到日期:2015-12-08
文章編號:1000-4025(2016)01-0023-07
doi:10.7606/j.issn.1000-4025.2016.01.0023
