內蒙古不同草地類型隱子草種群對放牧強度的響應
2016-04-05 08:28:57秦潔韓國棟王忠武喬江武倩內蒙古農業大學生態環境學院內蒙古呼和浩特0008中國農業科學院草原研究所內蒙古呼和浩特0000
生態環境學報 2016年1期
秦潔,韓國棟*,王忠武,喬江,2,武倩.內蒙古農業大學生態環境學院,內蒙古 呼和浩特 0008;2.中國農業科學院草原研究所,內蒙古 呼和浩特 0000
?
內蒙古不同草地類型隱子草種群對放牧強度的響應
秦潔1,韓國棟1*,王忠武1,喬江1,2,武倩1
1.內蒙古農業大學生態環境學院,內蒙古 呼和浩特 010018;2.中國農業科學院草原研究所,內蒙古 呼和浩特 010010
摘要:放牧脅迫使隱子草(Cleistogenes)種群成為退化草地的優勢種,而繼續加重放牧致使隱子草逐步衰退。由于在退化過程中隱子草作為退化指示種和重度退化的過渡種其種群格局將發生變化,故以內蒙古草甸草原、典型草原、荒漠草原為研究對象,設置輕度、中度、重度以及對照樣地4個放牧強度,經過5年的連續野外試驗和研究,對隱子草在不同放牧強度下的變化趨勢進行分析,以期為草地管理提供理論基礎。研究結果表明,(1)無論是草甸草原還是典型草原、荒漠草原,退化草地指示種隱子草都呈現明顯的隨放牧強度增加而先增加后減少的趨勢,只是不同草地類型對于年度以及峰值出現的情況是有差異的。(2)草甸草原和典型草原都是以中度放牧強度顯著最高,而荒漠草原只在降水相對較高的2008年以中度放牧最高,在降水較少的2006年、2007年則以輕度放牧顯著最高。(3)不同草地類型雙因素方差分析結果表明只在典型草原中,年度因素對無芒隱子草(Cleistogenes songorica)有顯著影響。草地類型和放牧對隱子草有顯著影響(P<0.01),且草地類型、放牧和年度的兩兩交互作用對隱子草的影響顯著(P<0.01)。
關鍵詞:草地;隱子草;放牧;降水
QIN Jie,HAN Guodong,WANG Zhongwu,QIAO Jiang,WU Qian.Response of Cleistogenes to Grazing Intensity in Different Grasslands of Inner Mongolia [J].Ecology and Environmental Sciences,2016,25(1):36-42.
草地是全球分布面積最大的陸地生態系統,約占陸地面積的三分之一(White et al.,2000),具有重要的生態和社會功能(Wrage et al.,2011),為人類提供了許多生活產品(O'Mara,2012)和生態服務(Huyghe,2008)。我國是草地資源大國,擁有草地總面積近4×108hm2,約占國土總面積的41.7%(白永飛等,2014)。內蒙古草原是我國畜牧業的重要生產基地,內蒙古草原植被分布具有明顯的地帶性,由東到西依次分布有草甸草原、典型草原和荒漠草原3種草原類型(白永飛等,2000)。放牧是內蒙古草原的主要利用方式,放牧強度、放牧制度、放牧季節和放牧家畜種類都在很大程度上影響著草地生態系統的物種組成、草地生產力及土壤的理化性質。適度放牧使草地生物量、根系和凋落物數量、土壤動物和微生物的多樣性提高,從而促進土壤呼吸。但過度放牧導致草地群落覆蓋率降低、草地地上植被和地下根系生物量下降等草地退化現象發生(楊陽等,2012)。而我國目前有90%左右的草地發生了不同程度的退化,其中嚴重退化草地達60%以上(白永飛等,2014)。近年來由于我國在畜牧業生產中對草地的生產功能利用過度,而忽視了草地的生態功能,造成超載過牧、人-草-畜關系失衡和草地的大面積退化,并誘發了沙塵暴、荒漠化等嚴重的生態災難(白永飛等,2014)。
放牧是草地發生退化的主要因子,在退化的草甸草原和典型草原,放牧脅迫使糙隱子草(Cleistogenes squarrosa)種群在群落中處于優勢地位,當嚴重退化的群落圍欄封育后,隨著恢復演替時間的推移,放牧脅迫的影響逐漸消失,糙隱子草的優勢地位開始下降(王鑫廳等,2013)。而在西部地區的荒漠草原,短花針茅占優勢的草原退化成為以無芒隱子草(Cleistogenes songorica)和冷蒿(Artemisia frigida)為優勢種的群落(劉鐘齡等,1998;王彥榮等,2002)。所以放牧正是隱子草種群在退化過程中產生格局變化的關鍵,糙隱子草和無芒隱子草在草地退化過程中都表現出逐漸增長的趨勢,而在嚴重退化階段趨于衰退(劉鐘齡等,1998)。所以糙隱子草和無芒隱子草是內蒙古草地開始退化的指示種也是重度退化過程中的過渡種,具有重要的研究意義。
植物初級生產力是反映草地功能的重要指標(Loreau et al.,2001;Hooper et al.,2005)。環境因子(Swemmer et al.,2008;Yang et al.,2008;Bai et al.,2008)與土地利用方式(王靜等,2005;田大栓等,2009)是決定草地生產力的關鍵因素,生物和資源有效性等也可能是植物生產力大小的主導因子(Yang et al.,2008),所以植物生物量是草地退化過程中探討植被格局變化的重要指標。由于隱子草具有退化指示種的特征,研究其種群格局形成機制可以進一步認識其在放牧退化過程中優勢地位發生變化的實質。但是,隱子草在不同草地類型中隨著放牧強度增加的變化趨勢是否一致?分析其一致或差異的原因,探索隱子草對人為干擾的響應機制。對三類草地不同放牧強度梯度進行連續5年(2005—2009年)的調查研究,探索在不同放牧強度下隱子草的動態變化規律,為進一步揭示內蒙古草原放牧退化過程提供參考。
1 實驗與方法
1.1研究區概況
選取西烏珠穆沁旗作為草甸草原研究區,克什克騰旗作為典型草原研究區,四子王旗作為荒漠草原研究區。3個研究區概況見表1。

表1 不同草地類型的氣候狀況Table 1 The climate in different grassland types
草甸草原在研究期內降水較少,而典型草原和荒漠草原的降水波動較大。2008年是研究期內是三類草原中降水最高的一年(表2)。

表2 3個研究區2005—2009年降水情況Table 2 Three research area precipitation in 2005─2009
1.2樣地選取
天然草原表現為以居民點為中心,周圍相繼出現環帶狀退化系列,即由居民點向外輻射,沿半徑方向構成草原群落的放牧強度梯度,且越靠近居民點的地方受到的放牧干擾越大,而距離居民點較遠的草場受放牧活動的影響較小,從而使整個樣地內的植被、土壤狀況呈現漸變狀態,植物群落形成一個植被連續體,即隨著放牧強度的增大,形成一個退化系列。于2005年開始設置樣地,在不同的草地類型中選擇植被類型、土壤、地形均相似的草地利用單元,在同一個草地利用單元上選擇3戶家庭牧場,并以居民點為中心進行自由放牧式家庭放牧,其家庭人口及所養牲畜種類及數量相似,以這3戶家庭牧場作為研究的重復樣地。研究選取的樣地及試驗處理位于連續的同一地段,地勢平坦,環境條件相對均質,有效地避免了空間異質性的干擾。以牲畜飲水點或者牲畜棚圈為中心,沿草原群落變化方向每隔50 m建立一個20 cm×50 cm的樣方,在樣方內調查植被組成、產量、覆蓋度,依據植被重要值聚類分析分為3個組,也就是3個放牧強度,即重度放牧區(heavy grazing intensity,簡稱H)、中度放牧區(moderate grazing intensity,簡稱M)和輕度放牧區(light grazing intensity,簡稱L),另外選擇圍欄外不受干擾的樣地為對照樣地(control check,簡稱CK)。依據樣線的前期調查和數據分析,劃分出輕、中、重3個放牧梯度離中心的距離(見圖1),在草甸草原分別為740、520、240 m,在典型草原分別為420、300、180 m,在荒漠草原分別為2400、1700、600 m。對3個放牧強度調查統計草地利用率,其中輕度放牧強度下的草地利用率為24%~30%,中度放牧強度下的草地利用率為40%~44%,重度放牧強度下的草地利用率為65%~70%。沿樣線確定3個放牧強度的中心,以每個中心10 m處的3個不同的方向設置樣方作為每個重復樣地上的每個放牧梯度的3個重復,在每個草地類型中設置36個樣方。
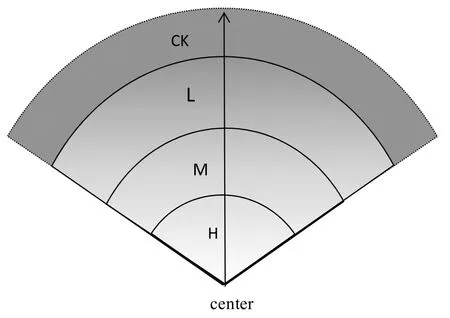
圖1 放牧系統原理圖Fig.1 Schematic diagram of the grazing system
1.3數據采集
于每年8月中旬植被生長旺盛期,進行樣方的測定,樣方大小為1 m×1 m。在草甸草原和典型草原中主要采集糙隱子草、在荒漠草原中主要采集無芒隱子草,而樣方內全部植物采用齊地面刈割的方法采集,使樣品風干后再在65 ℃下烘12 h后測定樣品的干重。
1.4數據分析
利用Excel進行數據整理。
隱子草相對生物量=隱子草生物量/總生物量×100%
利用SPSS 20.0廣義線性模型(generalized linear model,GLM)完成3因素的方差分析(Three-way ANOVAs),以確定草原類型(T)、放牧強度(G)、年度(Y)以及交互作用(T×G)、(T×Y)、(G×Y)、(T×G×Y)對隱子草相對生物量的影響,具體模型如下:
C=a+T+G+Y+(T×G)+(T×Y)+(G×Y)+(T×G×Y)+e
其中:C為依變量,a為總體平均值,T為不同草原類型對依變量的固定影響,G為不同放牧強度對依變量的固定影響,Y為年度對依變量的固定影響,C為年度對依變量的固定影響,T×G為草原類型和放牧強度的交互作用,T×Y為草地類型與年度的交互作用,G×Y為放牧強度與年度的交互作用,T×G×Y為草地類型、放牧強度和年度3因子的交互作用,e為試驗的隨機誤差。
進一步對3個草地類型分別進行年度和放牧強度的雙因素方差分析(Two-way ANOVAs)。對隱子草相對生物量進行不同放牧梯度間的單因素方差分析(One-way ANOVA),并進行Duncan多重比較,年度之間也進行相同的單因素方差分析和多重比較。
2 結果與分析
不同草地類型間隱子草具有顯著差異(P<0.01),放牧對隱子草相對生物量有顯著影響(P<0.01),年度的影響不顯著,草地類型、放牧和年度3個因子的兩兩交互作用對隱子草的影響顯著(P<0.01),但是草地類型、放牧強度和年度3因素交互對羊草的影響沒有達到顯著水平(表3)。

表3 草原類型、放牧強度和年度對隱子草的3因素方差分析結果Table 3 Three-way ANOVAs of Cleistogenes under different grassland types,grazing intensities and years
草甸草原中不同放牧強度下糙隱子草相對生物量有顯著差異,中度放牧強度下糙隱子草相對生物量顯著高于其他放牧處理,這一差異在2007、2008和2009年達到顯著水平(P<0.05)。不同年度之間糙隱子草相對生物量沒有達到顯著差異(圖2)。但是雙因素方差分析結果表明,放牧、年度以及兩因素的交互對草甸草原糙隱子草均沒有顯著影響(表4)。
典型草原中糙隱子草在不同放牧強度之間有顯著差異(圖3),2007年的輕度放牧顯著高于其他放牧強度,在2008年中度放牧顯著高于其他放牧強度(P<0.05)。典型草原年度對糙隱子草相對生物量的影響較為明顯(P<0.05)(圖3、表4),輕度放牧在2008年顯著高于2005年,中度放牧在2008年顯著高于其他年份(P<0.05)。但是雙因素方差結果表明,放牧以及與年度的交互作用對典型草原糙隱子草相對生物量沒有顯著影響(表4)。
在荒漠草原中,無芒隱子草相對生物量在不同放牧強度下有顯著差異(圖4),輕度放牧中顯著高于其他放牧強度且差異在2006年和2007年達到顯著水平,中度放牧高于其他放牧強度且差異在2008年達到顯著水平(P<0.05)。年度間無芒隱子草有顯著差異(圖4),在輕度放牧樣地中2006年和2007年顯著高于其他年份(P<0.05)。但是雙因素方差結果表明,放牧強度、年度以及雙因素的交互作用對荒漠草原無芒隱子草沒有達到顯著影響(表4)。
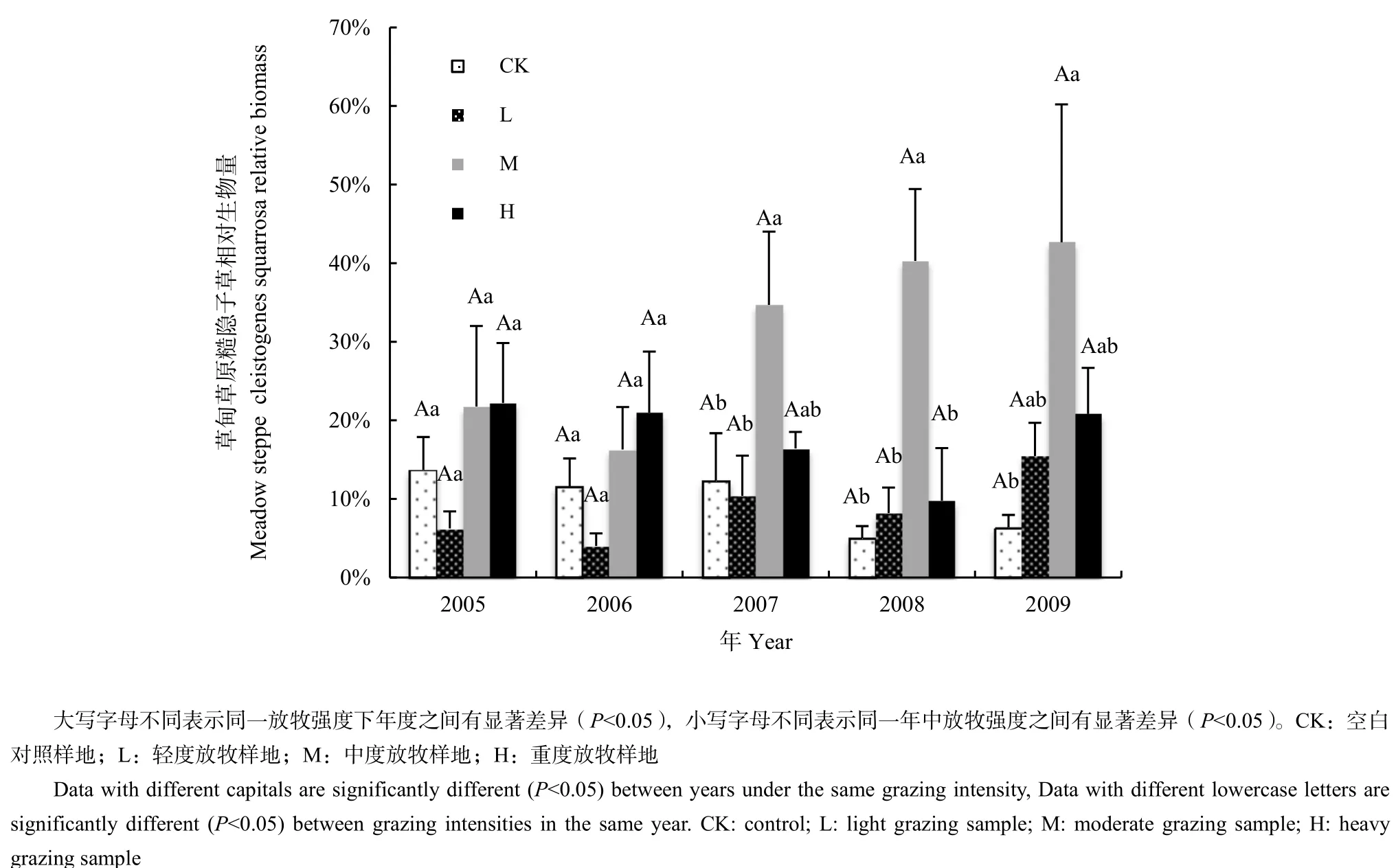
圖2 草甸草原不同放牧強度5年中糙隱子草相對生物量比較(n=137)Fig.2 The comparison of Cleistogenes squarrosa relative biomass in different grazing intensities in meadow steppe in five years

表4 不同草原類型中放牧強度和年度對隱子草的雙因素方差分析結果影響Table 4 Two-way ANOVAs of Cleistogenes under different grazing intensities and years in different grassland types
3 結論
不同草地類型中隱子草有顯著差異(P<0.01),放牧對隱子草相對生物量有顯著影響(P<0.01),年度的影響不顯著,草地類型、放牧和年度的兩兩交互作用對隱子草的影響顯著(P<0.01),但是草地類型、放牧強度和年度三者交互對羊草的影響沒有達到顯著水平。不同草地類型雙因素方差分析結果表明只在典型草原中年度對隱子草有顯著影響(P<0.05)。
無論是草甸草原還是典型草原、荒漠草原,退化草地指示種隱子草生物量都呈現明顯的隨放牧強度增加而先增加后減少的趨勢,只是不同草地類型對于年度以及峰值出現的情況是有差異的。草甸草原和典型草原都是在中度放牧強度下顯著高于其他處理,而荒漠草原只在2008年降水相對高的年份中以中度放牧最高,在2006、2007年降水較少的年份則以輕度放牧顯著最高。
4 討論
4.1放牧對隱子草的影響
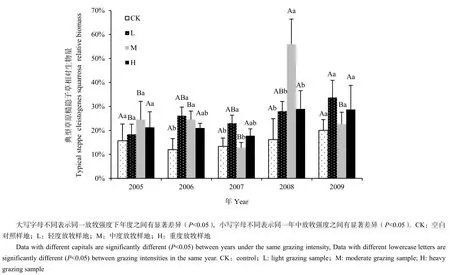
圖3 典型草原不同放牧強度5年中糙隱子草相對生物量比較(n=187)Fig.3 The comparison of Cleistogenes squarrosa relative biomass in different grazing intensity in typical steppe in five years
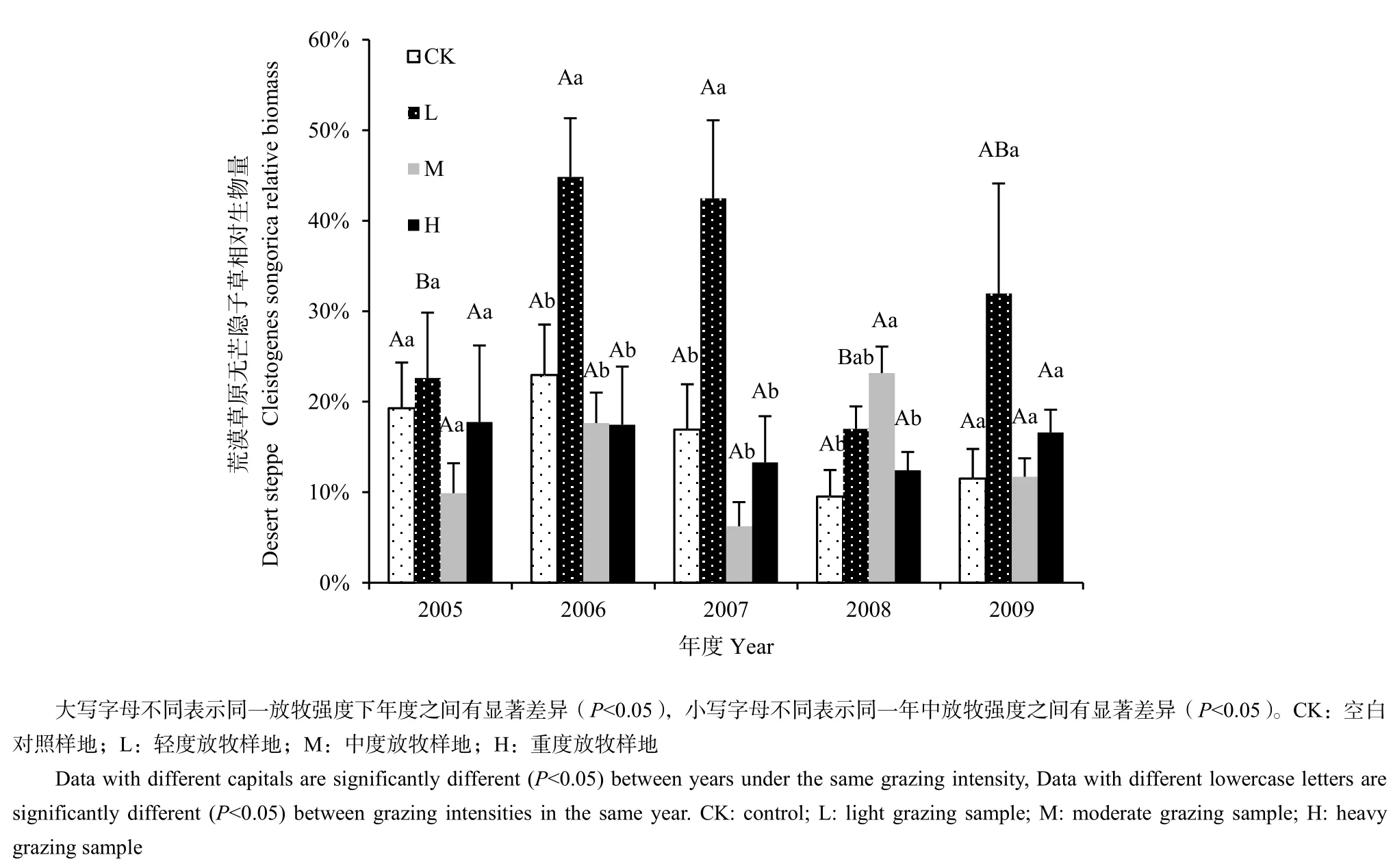
圖4 荒漠草原不同放牧強度五年中無芒隱子草相對生物量比較(n=234)Fig.4 The comparison of Cleistogenes songorica relative biomass in different grazing intensities in desert steppe in five years
放牧使得草地大量植被被牲畜啃食并對表層土壤造成破壞,同時動物糞便也改變了土壤表層的理化性質,放牧不同程度地影響到植被自身以及植被生長的環境(韓大勇等,2011;楊陽等,2012;孫宗玖等,2014)。三類草原中,隱子草都呈現隨放牧強度的增加而先增加后減少的趨勢,并在輕牧或者中牧階段達到高峰。而過牧后,由于土壤鹽堿化加重,不耐鹽堿植物逐漸消退,鹽生植物種群優勢度迅速增加,群落發生演替(王仁忠等,1995)。糙隱子草作為家畜喜食的植物,在輕、中牧條件下相對生物量比較大,說明其耐牧性較高。一定程度的過牧可刺激根莖發育從而促進地上生長(汪詩平等,2001),糙隱子草自身存在超補償性生長是其和家畜表現出協同進化的適應機制(劉振國等,2006),是其對環境變化所采取的適應策略(王鑫廳等,2013)。植物對于放牧的適應性策略也是多樣的,除補償性生長外,其生殖策略的改變也是對放牧干擾的適應。在生態環境較好條件下,植被多以有性繁殖為主(閆芊等,2007),當環境趨于惡劣則多以無性繁殖為主,所以隨著放牧脅迫的加重,以無性繁殖為主的植物逐漸成為優勢種(劉振國等,2004;田大栓等,2009)。隱子草可以進行有性繁殖也可以進行無性繁殖,在惡劣環境下可以改變生殖策略通過無性繁殖完成種群的擴展(張曉娜等,2010),并由于其多年生和叢生無性繁殖的特性,使得種群在放牧脅迫下可以保持長期穩定(張繼義等,2004)。有研究表明在放牧導致的退化群落中,放牧脅迫的作用使隱子草種群在群落中處于優勢地位,種群個體間的相互作用為正相互作用,種內相互促進,競爭力加強(王鑫廳等,2013)。糙隱子草在環境改變后通過其生殖策略的變化,、自身補償作用以及種內關系等一系列的變化使其很快適應環境以提高自身優勢度并使得種群的穩定性保持下去。
4.2隱子草在放牧演替過程中的變化趨勢
生態系統非生物環境中水分條件的變化,會對生態系統的生物構成產生影響,從而導致生態系統的演化和演替。即便只是小尺度的土壤養分和土壤水分空間分布格局的改變也可能成為草原群落向不同方向演替的主要驅動因子(白永飛等,2002),反過來植被自然發展與進化也會影響到地下水分再分配(劉美珍等,2006)。在草甸草原中水土保持良好,年度氣候波動并不會影響到隱子草在整個群落中的生長情況,而半濕潤半干旱的典型草原受降水的影響較大,所以年度對典型草原的影響較為顯著。且在荒漠草原和典型草原都出現過在降水較少的年份隱子草相對生物量在輕度放牧強度下達到最高,干旱促使隱子草優勢度增加,加速了放牧導致的逆向演替。對糙隱子草的研究(秦潔等,2015)也表明輕度退化草地中降水量的增加可以促使糙隱子草根系小型化,這對于退化草地的恢復是有利的,而干旱反而加重草地的退化程度。
放牧影響下草原植物的生態適應對策,決定其能否忍耐或適應放牧生境而維持生存并進行種群更新。隱子草在入侵-擴展-優勢-衰退的種群動態過程,其變化模式可以用1條單峰曲線來描述,研究結果與張繼義等人從生態位角度研究種群動態趨勢的結果相一致,隨著草地的退化其生態位寬度要經歷從小到大,再從大到小的過程(張繼義等,2003)。不同植物對于放牧壓力增加所采取的適應對策不同,從而決定了隱子草在植物群落中的替代規律(魏紹成等,1999),這也正是放牧影響下植被替換的機理所在。
參考文獻:
BAI Y F,WU J G,QI X,et al.2008.Primary Production and Rain Use Efficiency across a Precipitation Gradient on the Mongolia Plateau [J].Ecology,89(8):2140-2153.
HOOPER D U,CHAPIN F S,EWEL J J,et al.2005.Effects of biodiversity on ecosystem functioning:A consensus of current knowledge [J].Ecological Monographs,75(1):3-35.
HUYGHE C.2008.Multi-function grasslands in France:I.Production functions [J].Cahiers Agricultures,17(5):427-435.
LOREAU M,NAEEM S,INCHAUSTI P,et al.2001.Biodiversity and Ecosystem Functioning:Current Knowledge and Future Challenges [J].Science,294(5543):804-808.
O'MARA F P.2012.The role of grasslands in food security and climate change [J].Annals Of Botany,110(6):1263-1270.
SWEMMER A M,KNAPP A K.2008.Defoliation Synchronizes Aboveground Growth of Co-Occurring C? Grass Species [J].Ecology,89(10):2860-2867.
WHITE R,MURRAY S,ROHWEDER M.2000.Pilot analysis of global ecosystems:Grassland ecosystems [M].Washington DC,World Resources Institute:11-28.
WRAGE N,STRODTHOFF J,CUCHILLO H M,et al.2011.Phytodiversity of temperate permanent grasslands:ecosystem services for agriculture and livestock management for diversity conservation [J].Biodiversity and Conservation,20(14):3317-3339.
YANG Y,FANG J,MA W,et al.2008.Relationship between variability in aboveground net primary production and precipitation in global grasslands [J].Geophysical Research Letters,35(23):1-4.
白永飛,黃建輝,鄭淑霞,等.2014.草地和荒漠生態系統服務功能的形成與調控機制[J].植物生態學報,38(2):93-102.
白永飛,許志信,李德新.2000.內蒙古高原針茅草原群落α多樣性研究[J].生物多樣性,8(4):353-360.
白永飛,許志信,李德新.2002.內蒙古高原針茅草原群落土壤水分和碳、氮分布的小尺度空間異質性[J].生態學報,22(8):1215-1223.
韓大勇,楊永興,楊楊,等.2011.放牧干擾下若爾蓋高原沼澤濕地植被種類組成及演替模式[J].生態學報,31(20):5946-5955.
劉美珍,孫建新,蔣高明,等.2006.植物-土壤系統中水分再分配作用研究進展[J].生態學報,26(5):1550-1557.
劉振國,李鎮清.2004.不同放牧強度下冷蒿種群小尺度空間格局[J].生態學報,24(2):227-234.
劉振國,李鎮清.2006.退化草原冷蒿群落13年不同放牧強度后的植物多樣性[J].生態學報,26(2):475-482.
劉鐘齡,王煒,梁存柱,等.1998.內蒙古草原植被在持續牧壓下退化演替的模式與診斷[J].草地學報,6(4):244-251.
秦潔,鮑雅靜,李政海,等.2015.退化草原糙隱子草根系特征及地上高度對水分梯度的響應[J].中國草地學報,37(3):80-86.
孫宗玖,朱進忠,張鮮花.2014.短期放牧強度對昭蘇草甸草原植被特征及多樣性影響[J].新疆農業大學學報,37(1):35-39.
田大栓,包祥,關其格,等.2009.內蒙古草原3種針茅屬植物不同組織水平的生物量生殖分配[J].植物生態學報,33(1):97-107.
汪詩平,王艷芬.2001.不同放牧率下糙隱子草種群補償性生長的研究[J].植物學報,43(4):413-418.
王靜,楊持,王鐵娟.2005.放牧退化群落中冷蒿種群生物量資源分配的變化[J].應用生態學報,16(12):2316-2320.
王仁忠,李建東.1995.羊草草地放牧退化演替中種群消長模型的研究[J].植物生態學報,19(2):170-174.
王鑫廳,王煒,梁存柱,等.2013.不同恢復演替階段糙隱子草種群的點格局分析[J].應用生態學報,24(7):1793-1800.
王彥榮,曾彥軍,付華,等.2002.過牧及封育對紅砂荒漠植被演替的影響[J].中國沙漠,22(4):14-20.
魏紹成,孫振權,王紅俠,等.1999.內蒙東部大針茅草原群落類型和群落間的生態關系[J].草業學報,8(1):1-8.
閆芊,陸健健,何文珊.2007.崇明東灘濕地高等植被演替特征[J].應用生態學報,18(5):1099-1103.
楊陽,韓國棟,李元恒,等.2012.內蒙古不同草原類型土壤呼吸對放牧強度及水熱因子的響應[J].草業學報,21(6):8-14.
張繼義,趙哈林,張銅會,等.2003.科爾沁沙地植物群落恢復演替系列種群生態位動態特征[J].生態學報,23(12):2741-2746.
張繼義,趙哈林.2004.科爾沁沙地草地植被恢復演替進程中群落優勢種群空間分布格局研究[J].生態學雜志,23(2):1-6.
張曉娜,哈達朝魯,潘慶民.2010.刈割干擾下內蒙古草原兩種叢生禾草繁殖策略的適應性調節[J].植物生態學報,34(3):253-262.
Response of Cleistogenes to Grazing Intensity in Different Grasslands of Inner Mongolia
QIN Jie1,HAN Guodong1*,WANG Zhongwu1,QIAO Jiang1,2,WU Qian1
1.College of Ecology and Environmental Science,Inner Mongolia Agricultural University,Huhhot 010018,China; 2.Grassland Research Institute of chinese academy of agricultural science,Huhhot 010010,China
Abstract:Cleistogenes populations become dominant species under moderate grazing pressure,and began to decline with a heavier grazing in the degraded grassland.There is a special pattern that Cleistogenes are indicator species in degraded grasslands and a transient species in the severe degradation grasslands during the degradation process.In this research,we focused on Cleistogenes populations changes under four grazing intensity (Light grazing,moderate grazing,heavy grazing and control) in meadow steppe,typical steppe and desert steppe in Inner Mongolia five years continuous experiment and tried to analysis the variation tendency of Cleistogenes with grazing intensity,to provide theoretical basis for the management of rangeland.The results showed that:(1) Cleistogenes had an increasing trend at first and then decreased in th three types of steppes with the increasing grazing intensity.But the peak points were different in different years and steppes.(2) The relative biomass of Cleistogenes squarrosas was the the highest under moderate grazing in meadow steppe and typical steppe in every experimental year,while the relative biomass of Cleistogenes songorica was the highest only in 2008 with more precipitation under moderate grazing,and in 2006 and 2007 with insufficient precipitation under light grazing in desert steppe.(3) By comparison of two-factor analysis of different steppe types,the years have significant effect on Cleistogenes squarrosas in the typical steppe.The steppe types,grazing intensities and the interaction have strong effect on Cleistogenes.And the effects of the interaction between years and steppe types,the interaction between years and grazing intensities on Cleistogenes were significant.
Key words:grassland; degradation; Cleistogenes; grazing
收稿日期:2015-09-23
作者簡介:秦潔(1987年生),女,博士研究生,主要研究退化草地恢復和草地群落演替。E-mail:qinjie331313@163.com*通信作者。韓國棟,E-mail:nmghanguodong@163.com
基金項目:國家自然科學基金項目(31270502);內蒙古科技攻關項目“不同生態類型區域生態監測與評估及風險預測技術研究”;中澳國際合作項目(LPS/2008/048);研究生科研創新項目(B20151012906Z)
中圖分類號:Q948; X17; S812
文獻標志碼:A
文章編號:1674-5906(2016)01-0036-07
DOI:10.16258/j.cnki.1674-5906.2016.01.006
引用格式:秦潔,韓國棟,王忠武,喬江,武倩.內蒙古不同草地類型隱子草種群對放牧強度的響應[J].生態環境學報,2016,25(1):36-42.
