茂原鏈霉菌產谷氨酰胺轉氨酶發酵培養基優化
2016-04-12 03:39:40羅水忠鐘昔陽姜紹通
合肥工業大學學報(自然科學版) 2016年2期
汪 燕, 羅水忠, 蔡 靜, 鐘昔陽, 姜紹通, 鄭 志
(1.合肥工業大學 生物與食品工程學院,安徽 合肥 230009; 2.安徽省農產品精深加工重點實驗室,安徽 合肥 230009)
?
茂原鏈霉菌產谷氨酰胺轉氨酶發酵培養基優化
汪燕1,2,羅水忠1,2,蔡靜1,2,鐘昔陽1,2,姜紹通1,2,鄭志1,2
(1.合肥工業大學 生物與食品工程學院,安徽 合肥230009; 2.安徽省農產品精深加工重點實驗室,安徽 合肥230009)
摘要:文章考察了不同培養基成分對茂原鏈霉菌產谷氨酰胺轉氨酶的影響,以酶活和生物量為測定指標,采用單因素試驗和正交試驗確定發酵培養基最適成分及其質量濃度,以期為未來菌株擴大培養和發酵中試化提供參考。試驗結果表明,優化后的培養基為:葡萄糖40 g/L,蛋白胨40 g/L,硝酸銨1 g/L,酵母膏2 g/L,硫酸鎂2 g/L,磷酸氫二鉀2 g/L。該條件下酶活可達(1.30±0.07) U/mL,生物量為(9.69±0.23) g/L。
關鍵詞:茂原鏈霉菌;谷氨酰胺轉氨酶;發酵培養基;正交試驗
0引言
谷氨酰胺轉氨酶(Transglutaminase,EC.2.3.2.13,TG酶),又稱轉谷氨酰胺酶,可以催化蛋白質賴氨酸上的ε-氨基和谷氨酸上的γ-酰胺基結合,從而導致蛋白質(或多肽分子)之間發生共價交聯,形成相應的聚合產物[1]。
TG酶有著“21世紀超級黏合劑”的美稱,在食品加工領域應用廣泛。它可以改善肉制品的質構,提高原料的利用率[2-3];增加乳制品的凝膠強度,減少脂肪含量[4-5];生產可食性薄膜,利于食品的包裝和貯藏[6-7]等。這些優越的特性使得TG酶成為近年來研究的熱點,同時獲得高產量高酶活的TG酶也成為研究的關鍵。目前,研究主要集中于誘變育種篩選高效產酶菌株[8-9]、菌株發酵條件優化[10-11]、發酵酶液分離純化[12-13]和通過基因工程技術對現有的微生物進行基因改造[14-15]等。
微生物發酵法是工業上生產TG酶的主要方法,微生物代謝產酶的能力,除受菌株自身特性影響外,還受發酵條件如培養基、培養溫度[16]、搖床轉速等諸多客觀因素的影響[17]。文獻[18-19]報道對鏈霉菌HS-1進行培養基優化,酶活分別提高了16.04%和16.40%,因此,優化菌株產酶發酵條件對產物質量濃度、產量和產率具有重要的意義。本文通過單因素試驗和正交試驗對茂原鏈霉菌產TG酶的發酵培養基組成進行優化,為TG酶的工業化應用提供參考。
1材料與方法
1.1材料與試劑
1.1.1菌種
茂原鏈霉菌(Streptomycesmobaraensis)為合肥工業大學農產品加工研究院選育。
1.1.2培養基
斜面保存培養基:可溶性淀粉 20 g/L,KNO31 g/L,MgSO4·7H2O、K2HPO4·H2O、NaCl均為0.5 g/L,FeSO4·H2O 0.01 g/L,瓊脂20 g/L,pH值為7.2~7.4。
液體種子培養基:可溶性淀粉和蛋白胨各為20 g/L,酵母膏、MgSO4、K2HPO4均為2 g/L,pH值為7.0。
基礎發酵培養基:可溶性淀粉和蛋白胨各為30 g/L,酵母膏、NH4(SO4)2、MgSO4、K2HPO4均為2 g/L,pH值為7.0。
1.1.3主要試劑
N-α-CBZ-Gln-Gly和L-谷氨酸-γ-單羥肟酸,美國Sigma公司;還原型谷胱甘肽,北京索萊寶科技有限公司;其他均為國產分析純試劑。
1.2方法
1.2.1培養方法
液體種子的培養方法為:用接種環挑取少許生長良好的菌種置于新鮮斜面培養基上,28 ℃恒溫培養7 d。經活化后的菌種再接入種子培養基中,30 ℃,170 r/min,培養48 h。
搖瓶發酵的培養方法為:初始pH值為7.0,250 mL三角瓶裝液量25 mL,接種量10% (體積分數),30 ℃、170 r/min培養72 h。
1.2.2培養基優化試驗設計
分別考察發酵培養基中不同碳源及其質量濃度(可溶性淀粉、蔗糖、麥芽糖、葡萄糖、甘油)、不同有機氮源及其質量濃度(胰蛋白胨、魚粉蛋白凍、酪蛋白、蛋白胨、玉米漿)、不同無機氮源及其質量濃度(硫酸銨、硝酸銨、硝酸鈉)對茂原鏈霉菌發酵產TG酶的影響。根據單因素試驗確定的培養基成分及其質量濃度范圍,設計三因素三水平正交試驗。試驗因素水平見表1所列。

表1 正交試驗因素水平 g/L
1.2.3測定方法
(1) TG酶活的測定。按Grossowicz比色法[20]測定酶活:反應液A由0.2 mol/L Tris-HCl(pH=6.0)、0.1 mol/L鹽酸羥胺、0.01 mol/L還原型谷胱甘肽和0.03 mol/L N-α-CBZ-Gln-Gly組成,終止液B由3 mol/L鹽酸、12% 三氯乙酸、5% 六水三氯化鐵(溶解于0.1 mol/L鹽酸中)3種試劑等體積混合而成。樣品測定時,取200 μL酶液加入1 mL試劑A于37 ℃反應10 min后,加入1 mL試劑B終止反應,4 ℃、7 000 r/min離心10 min后在525 nm處比色。對照組為200 μL樣品先加試劑終止液B后于37 ℃反應10 min,再加試劑A,其他操作相同。1單位TG酶活性定義為:37 ℃條件下反應,每1 min生成1 μmol 的單羥肟酸所需的酶量,單位為U/mL。
(2) 菌體生物量(DCW)的測定。事先稱好過濾用的定量濾紙,將發酵液過濾后,用蒸餾水洗滌菌體3次,105 ℃干燥至恒質量。在干燥器中降至室溫后稱質量,菌體生物量為2次濾紙質量差,單位為g/L。
1.3數據處理
每組試驗重復3次,結果取平均值,試驗數據以“平均值±標準差”表示。
2結果與分析
2.1碳源種類對發酵的影響
不同碳源種類對發酵的影響如圖1所示。
碳源在微生物生長和代謝過程中有著舉足輕重的作用,是提供細胞生命活動所需能量和合成產物的碳架[21]。由圖1可知,不同碳源條件下培養茂原鏈霉菌產TG酶,其酶活和菌體生長差別較大。葡萄糖作為碳源時,酶活和生物量均為最大,分別為(0.86±0.01)U/mL和(8.11±0.11) g/L。因此,選擇適宜的碳源為葡萄糖。
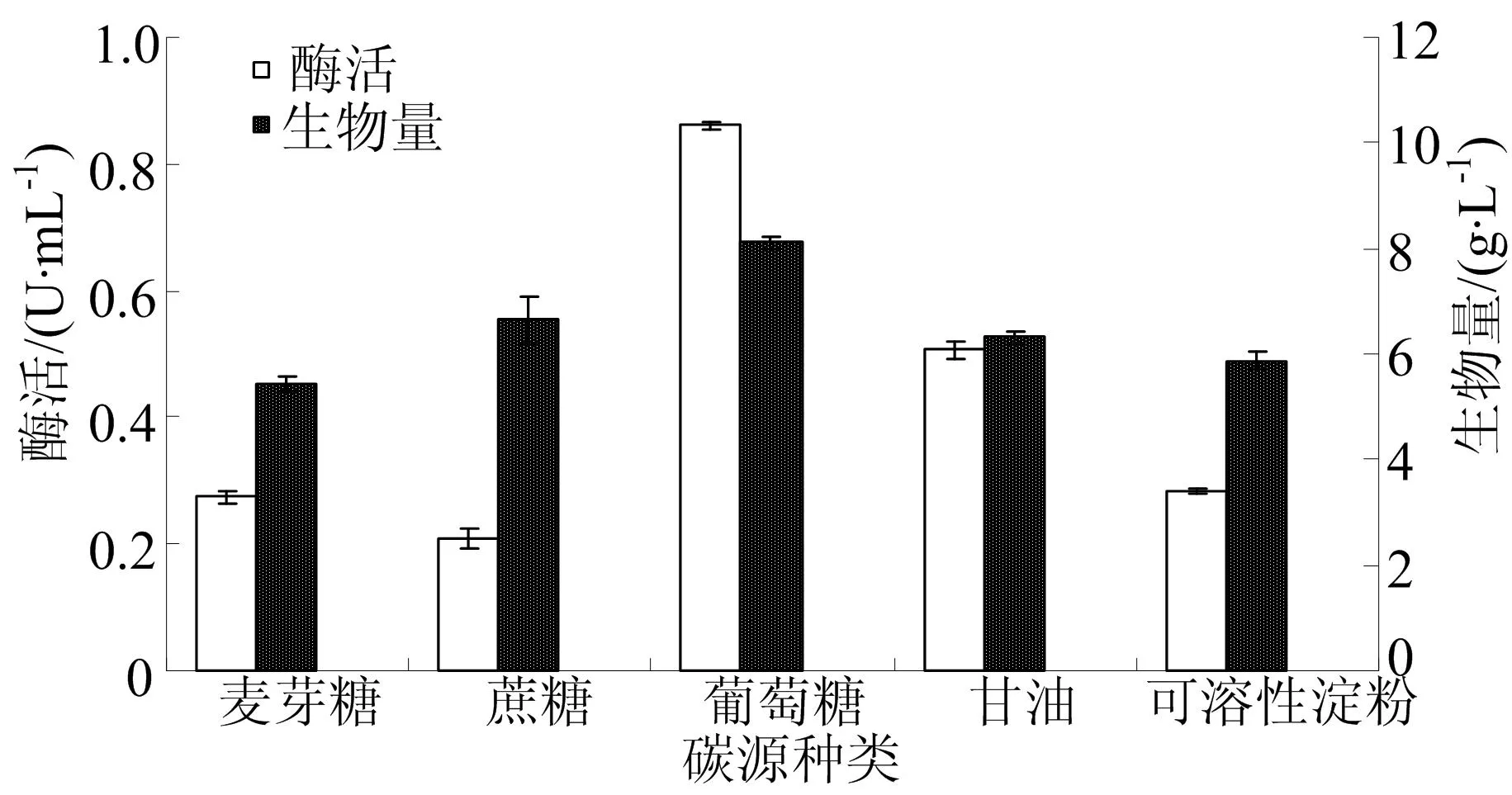
圖1 不同碳源種類對發酵的影響
2.2碳源添加量對發酵的影響
不同葡萄糖質量濃度對發酵的影響如圖2所示。
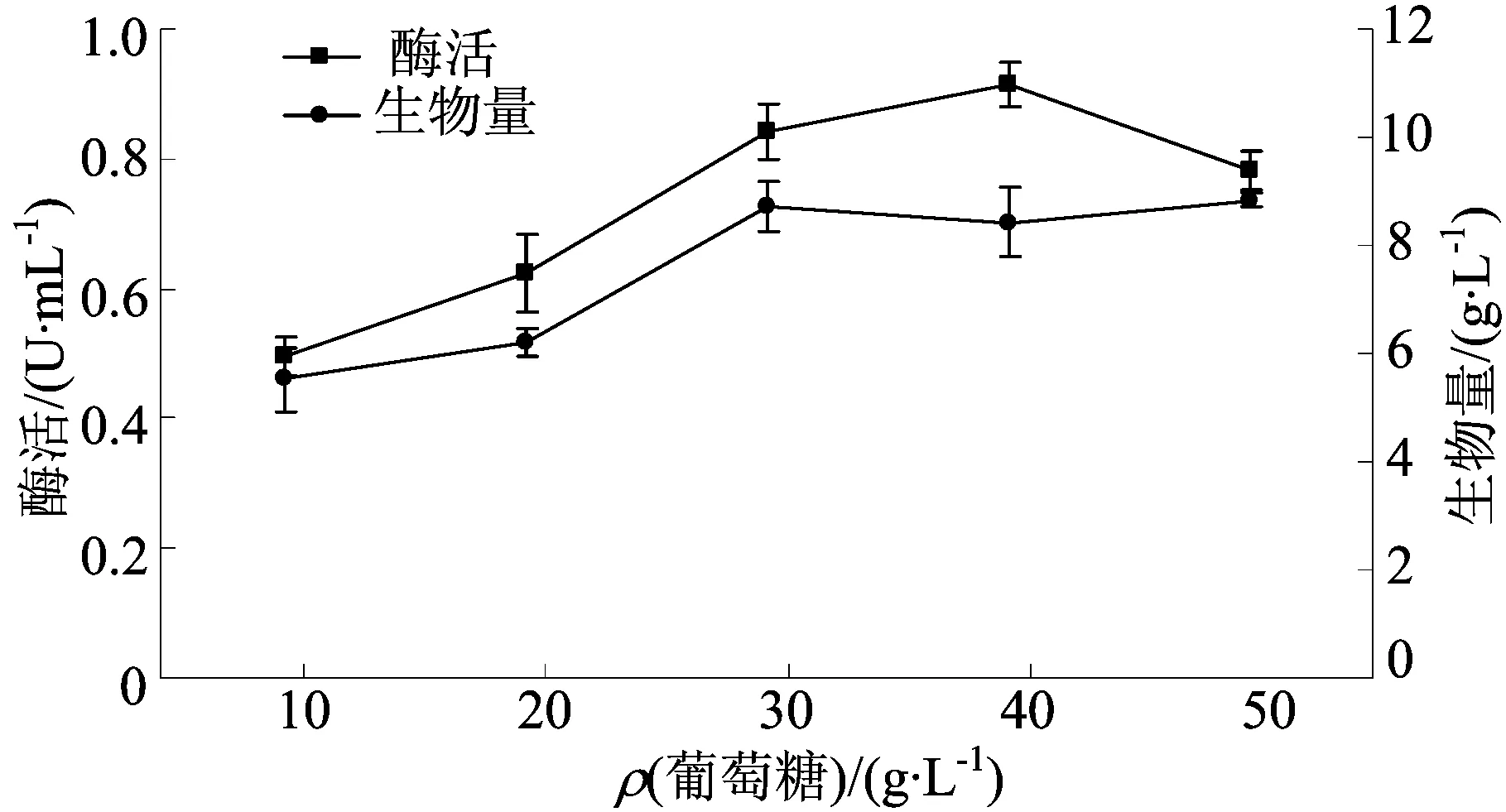
圖2 不同葡萄糖質量濃度對發酵的影響
由圖2可知,TG酶活和生物量隨著葡萄糖質量濃度的增加呈現先增后減的趨勢。當質量濃度為40 g/L時酶活達到最大,為(0.91±0.03)U/mL,此時生物量為(8.72±0.46)g/L。當質量濃度為50 g/L時,酶活急劇下降而生物量也幾乎沒有增加,說明過多的碳源并不利于菌株產酶和生長,也可能是TG酶的合成受碳源分解代謝物阻遏效應的影響[22]。因此,選擇葡萄糖質量濃度為40 g/L。
2.3有機氮源種類對發酵的影響
不同有機氮源種類對發酵的影響如圖3所示。
氮源是微生物生長和發酵的重要營養來源,微生物利用它在細胞內合成氨基酸和堿基,進而合成蛋白質、核酸等細胞成分以及含氮的代謝產物[23]。同時,氮源也是合成TG酶的主要底物。由圖3可知,產酶最適有機氮源為蛋白胨,該條件下,酶活可達(0.96±0.04)U/mL,此時生物量為(10.25±0.20)g/L;其次是玉米漿,為(0.86±0.04)U/mL和(8.93±0.28)g/L。而酪蛋白幾乎不能起到促進產酶的作用,酶活僅為(0.15±0.05)U/mL,生物量為(8.45±0.23)g/L。因此,選擇蛋白胨為有機氮源。
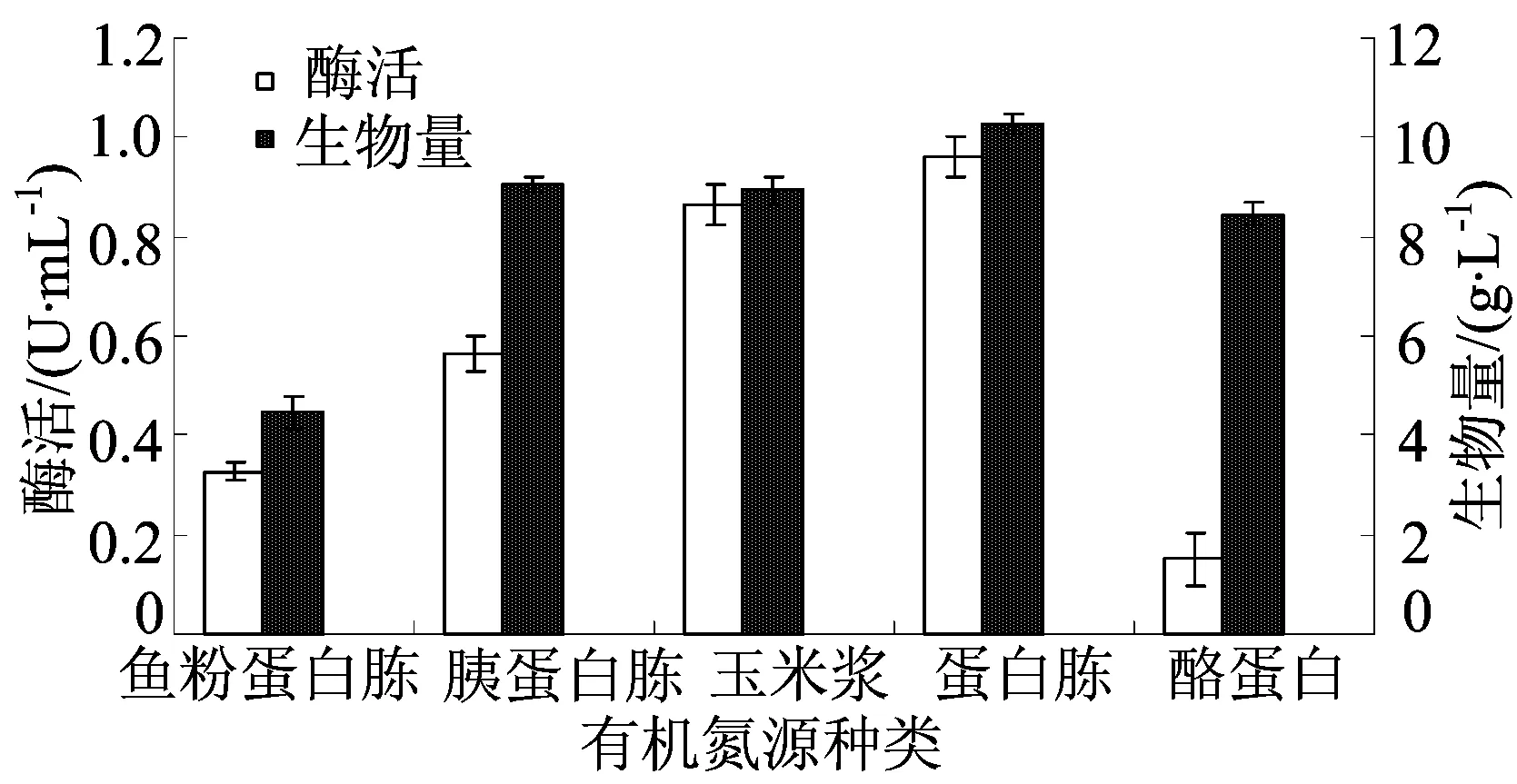
圖3 不同有機氮源種類對發酵的影響
2.4有機氮源添加量對發酵的影響
有機氮源添加量對發酵的影響如圖4所示。
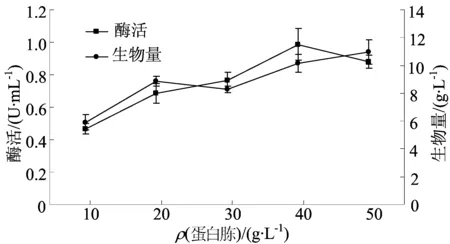
圖4 不同蛋白胨質量濃度對發酵的影響
由圖4可知,蛋白胨質量濃度從 10 g/L增加到40 g/L時,酶活約增大了1倍。當質量濃度為40 g/L時酶活達到最大值,為(0.99±0.10)U/mL,生物量為(10.12±0.65)g/L。質量濃度再增加到 50 g/L時,生長量與酶活均有明顯的下降。這可能是因為一方面氮源偏多,會使微生物細胞早期生長過于旺盛,導致菌體提前衰老自溶[24];另一方面是培養基中過量存在的蛋白胨類或游離氨基酸、小肽等在該酶的作用下共價交聯形成了聚合體,從而對菌體的物質吸收利用和代謝生長產生有害作用,影響了酶的生產[18]。因此,選擇蛋白胨質量濃度為40 g/L。
2.5無機氮源種類對發酵的影響
無機氮源種類對發酵的影響如圖5所示。
無機氮源是速效氮源,可以被細胞吸收后直接利用,有利于菌株前期的生長,而較高的酶活是建立在菌株生長良好的基礎之上。由圖5可知,添加硝酸銨的培養基,酶活有顯著增加,效果優于硝酸鈉和硫酸銨,說明硝酸銨能被菌株很好地利用。在該條件下,酶活可達(1.02±0.03)U/mL,此時生物量為(9.66±0.09)g/L。因此,選擇硝酸銨為適宜的無機氮源。

圖5 不同無機氮源種類對發酵的影響
2.6無機氮源添加量對發酵的影響
硝酸銨質量濃度對發酵的影響如圖6所示。由圖6可知,隨著硝酸銨質量濃度的增大,酶活反而呈下降的趨勢。當質量濃度為1 g/L時,酶活最大,可達(1.06±0.06)U/mL,此時生物量為(10.44±0.06)g/L。當質量濃度為5 g/L時,酶活僅為最大值的62%,生物量也有所下降。因此,選擇硝酸銨質量濃度為1 g/L。

圖6 不同硝酸銨質量濃度對發酵的影響
2.7培養基成分正交試驗
根據以上不同碳源和氮源單因素試驗結果和分析,選取葡萄糖、蛋白胨和硝酸銨進行三因素三水平的正交試驗,并以酶活作為考察指標,獲得有利于茂原鏈霉菌產TG酶的發酵培養基。選取的因素水平和試驗結果見表2所列。方差分析結果見表3所列。
由表2和表3可知,各因素影響酶活的主次順序為B>A>C,即蛋白胨>葡萄糖>硝酸銨。根據極差分析得最佳組合條件為:A2B2C1。其中B因素高度顯著,A和C因素顯著。
直接分析較好的條件是第5組,即A2B2C3,由于計算分析結果與直觀分析結果不一致,因此將2種組合同時進行驗證,每組重復3次。驗證方案與結果見表4所列。
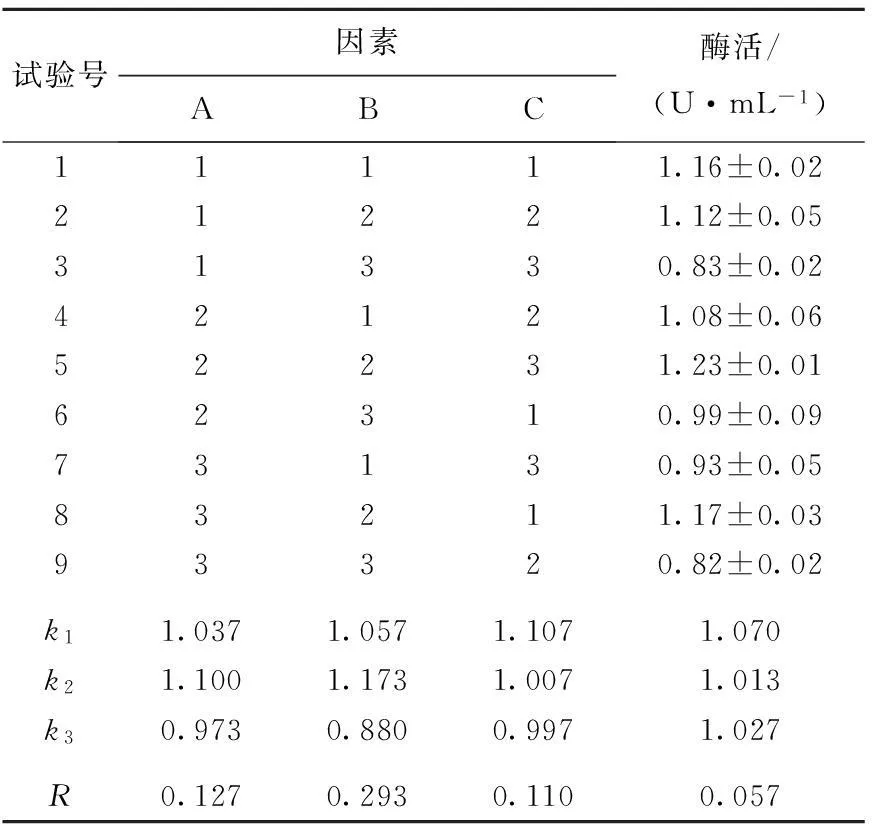
表2 正交試驗設計與結果
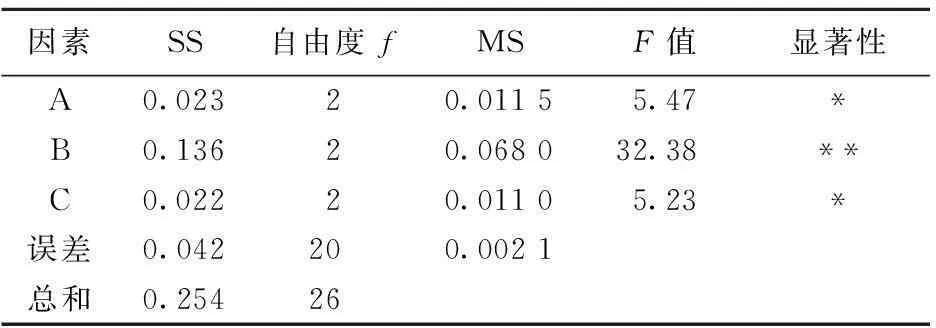
表3 正交試驗方差分析表
注:F0.05(2,20)=3.49,F0.01(2,20)=5.85。

表4 正交驗證試驗方案與結果
根據試驗結果,組合A2B2C1優于A2B2C3,因此選擇培養基的優化組合為A2B2C1,即蛋白胨40 g/L,葡萄糖40 g/L,硝酸銨1 g/L。
3結束語
單因素試驗表明,葡萄糖是適宜的碳源,最佳質量濃度為40 g/L,蛋白胨和硝酸銨是適宜的氮源,最佳質量濃度分別為40 g/L和1 g/L。通過正交試驗數據可知,對茂原鏈霉菌產TG酶的影響主次順序為蛋白胨>葡萄糖>硝酸銨。優化后的發酵培養基組成為:葡萄糖40 g/L,蛋白胨40 g/L,硝酸銨1 g/L,酵母膏 2 g/L,硫酸鎂2 g/L,磷酸氫二鉀 2 g/L。最終酶活可達(1.30±0.07) U/mL,此時生物量為(9.69±0.23) g/L。
[參考文獻]
[1]趙新淮,徐紅華,姜毓君.食品蛋白質:結構、性質與功能[M].北京: 科學出版社,2009:473.
[2]李寶臻,李海賓,劉爾卓,等.谷氨酰胺轉氨酶及其對肉制品凝膠特性的影響[J].農產品加工:學刊,2014 (1): 60-63.
[3]Yu X,Chen C G,Cai K Z,et al.Combined effect of blood plasma powder,agar,and microbial transglutaminase on physicochemical and textural properties of pork music gels[J].Food Science and Biotechnology,2012,21(4):941-950.
[4]張莉麗,韓雪,張蘭威,等.茂原鏈霉菌谷氨酰胺轉氨酶在酸奶中的應用研究[J].食品工業科技,2014,35(12):210-212.
[5]Sanli T,Sezgin E,Deveci O,et al.Effect of using transglutaminase on physical,chemical and sensory properties of set-type yoghurt [J].Food Hydrocolloids,2011,25(6):1477-1481.
[6]吳進菊,于博,豁銀強,等.轉谷氨酰胺酶改性可食用膜的研究進展[J].食品研究與開發,2013,34(4): 114-117.
[7]滑艷穩,陳志周.谷氨酰胺轉氨酶對大豆蛋白/PVA復合薄膜性能的影響[J].包裝與食品機械,2014(2):19-23.
[8]楊慧林,包瑩玲,潘力.片段化全基因組體外誘變選育轉谷氨酰胺酶高產菌株的研究[J].現代食品科技,2009,25(1):34-37.
[9]劉穎,田沛霖,陳佳,等.產谷氨酰胺轉胺酶菌株的高通量篩選[J].西北農林科技大學學報:自然科學報,2013,41(6): 167-172.
[11]Guerra-Rodríguez E,Vázquez M.Evaluation of a novel low-cost culture medium containing exclusively milk,potato and glycerol for microbial transglutaminase production bySreptomycesmobataensis[J].Chemical Engineering Research and Design,2014,92:784-791.
[12]郝偉,王璋,蔡慧農.微生物谷氨酰胺轉胺酶沉淀分離方法的研究[J].食品科學,2006,27(10):331-334.
[13]Macedo J A,Lara D S,Sato H H.Purification and characterization of a new transglutaminase fromStreptomycessp.isolated in Brazilian soil[J].Journal of Food Biochemistry,2011,35(4):1361-1372.
[14]儀朝印,張東杰,王穎.茂原鏈霉菌轉谷氨酰胺酶基因工程菌的構建及表達[J].中國食品學報,2014,14(2):42-46.
[15]Du K,Liu Z M,Cui W J,et al.pH-dependent activation ofStreptomyceshygropicustransglutaminase mediated by intein[J].Applied and Environmental Microbiology,2014,80(2):723-729.
[16]Yan G L,Du G C,Li Y,et al.Enhancement of microbial transglutaminase production byStreptoverticilliummobaraense:application of a two-stage agitation speed control strategy[J].Process Biochemistry,2005,40(2):963-968.
[17]Zheng M Y,Du G C,Guo W F,et al.A temperature-shift strategy in batch microbial transglutaminase fermentation[J].Process Biochemistry,2001,36(6):525-530.
[18]楊雪霞,常忠義,曹丹玥,等.谷氨酰胺轉胺酶發酵培養基的優化[J].東華大學學報:自然科學版,2008,34(1):77-80.
[19]曹丹玥,高紅亮,常忠義,等.谷氨酰胺轉胺酶發酵培養基的響應面分析優化[J].華東師范大學學報:自然科學版,2007(2): 93-97.
[20]Grossowicz N,Wainfan E,Borek E,et al.The enzymatic formation of hydroxamic acids from glutamine and asparagine[J].Journal of Biological Chemistry,1950,187(1):111—125.
[21]黃亞杰,王子輝,趙彥偉,等.重組大腸桿菌產谷氨酰胺轉氨酶培養基及發酵條件優化[J].中國釀造,2012,31(4):21-24.
[22]常中義,江波,王璋.培養基組成對輪枝鏈霉菌合成谷氨酰胺轉胺酶的影響[J].無錫輕工大學學報,2001,20(1):51-54.
[23]何國慶,賈英民,丁立孝.食品微生物學[M].第2版.北京:中國農業大學出版社,2009:78-79.
[24]周德慶.微生物學教程[M].北京: 高等教育出版社,1997:111.
(責任編輯閆杏麗)
Optimization of medium components for the production of transglutaminase byStreptomycesmobaraensis
WANG Yan1,2,LUO Shui-zhong1,2,CAI Jing1,2,ZHONG Xi-yang1,2,JIANG Shao-tong1,2,ZHENG Zhi1,2
(1.School of Biotechnology and Food Engineering, Hefei University of Technology, Hefei 230009, China; 2.Key Laboratory for Agricultural Products Processing of Anhui Province, Hefei 230009, China)
Abstract:The transglutaminase(TG) produced by Streptomyces mobaraensis was investigated with different culture media. TG enzyme activity and DCW were taken as indexes to screen for the optimal components and concentrations of medium by using single factors and orthogonal experiments. The optimized medium was as follows: the mass concentration of glucose was 40 g/L, peptone 40 g/L, ammonium nitrate 1 g/L, yeast extract 2 g/L, MgSO4 2 g/L and K2HPO4 2 g/L. Under this experimental condition, TG activity and the biomass were up to (1.30±0.07)U/mL and (9.69±0.23)g/L, respectively. This study provides a reference for amplification cultivation and fermentation of bacterial strain in the future.
Key words:Streptomyces mobaraensis; transglutaminase(TG); fermentation medium; orthogonal experiment
中圖分類號:TS201.3
文獻標識碼:A
文章編號:1003-5060(2016)02-0270-05
Doi:10.3969/j.issn.1003-5060.2016.02.024
作者簡介:汪燕(1990-),女,安徽黃山人,合肥工業大學碩士生;姜紹通(1954-),男,江蘇鹽城人,合肥工業大學教授,博士生導師;
基金項目:國家高技術研究發展計劃(863計劃)資助項目(2013AA102201);安徽省科技攻關計劃重大資助項目(1301031031)
收稿日期:2015-01-04;修回日期:2015-04-02
鄭志(1971-),男,安徽和縣人,博士,合肥工業大學教授,碩士生導師.
