不同寄主來源腐爛病菌對棗樹致病性研究
2016-04-13 06:32:00李春艷張王斌劉振亞姚永生但紅俠
新疆農(nóng)業(yè)科學(xué) 2016年1期
李春艷,張王斌,2 ,茍 巧 ,劉振亞 ,姚永生 ,但紅俠
(1.塔里木大學(xué)植物科學(xué)學(xué)院,新疆阿拉爾 843300;2.南疆農(nóng)業(yè)有害生物綜合治理重點實驗室,新疆阿拉爾 843300)
不同寄主來源腐爛病菌對棗樹致病性研究
李春艷1,張王斌1,2,茍 巧1,劉振亞1,姚永生1,但紅俠1
(1.塔里木大學(xué)植物科學(xué)學(xué)院,新疆阿拉爾 843300;2.南疆農(nóng)業(yè)有害生物綜合治理重點實驗室,新疆阿拉爾 843300)
【目的】明確棗樹腐爛病菌來源,為生產(chǎn)綜合防治提供理論依據(jù)。【方法】將不同寄主來源腐爛病菌分別接種于30%棗樹樹皮培養(yǎng)基、果實、枝條和田間接種枝條,觀察并計算發(fā)病率、病斑擴(kuò)展速度及產(chǎn)孢情況。【結(jié)果】在30%棗樹樹皮培養(yǎng)基上,不同寄主來源腐爛病菌均能在7 d內(nèi)產(chǎn)孢,來源于柳樹、蘋果、沙棗、棗樹腐爛病菌在30 d內(nèi)先后產(chǎn)生橘黃色分生孢子角;來源于沙棗、棗樹、胡楊及楊樹腐爛病菌均能在棗果上產(chǎn)孢,來源于蘋果和梨樹腐爛病菌未在果實上形成產(chǎn)孢體;不同寄主來源腐爛病菌均能在棗樹離體枝條上產(chǎn)孢并吐出分生孢子角,來源于桃樹和沙棗樹腐爛病菌在枝條上的侵染速度最快。田間枝條接種測定中來源于胡楊腐爛病菌病斑擴(kuò)展速度最快,接種胡楊和梨樹腐爛病菌的枝條在15 d內(nèi)產(chǎn)生小黑點并吐出黃色分生孢子角。【結(jié)論】不同寄主來源腐爛病菌均能在30%棗樹樹皮培養(yǎng)基繁殖并能形成產(chǎn)孢體,并能產(chǎn)生分生孢子角;均能侵染棗樹離體組織和田間健康枝條,但不同寄主來源腐爛病菌致病性存在明顯差異。
棗樹;腐爛病菌;致病力;擴(kuò)展速度;產(chǎn)孢體
0 引 言
【研究意義】隨著農(nóng)業(yè)產(chǎn)業(yè)結(jié)構(gòu)的調(diào)整、退耕還林政策的實施及南疆地理環(huán)境、氣候、土地資源等諸多天然優(yōu)勢,紅棗產(chǎn)業(yè)的發(fā)展已成為南疆地區(qū)又一支柱產(chǎn)業(yè),成為農(nóng)村經(jīng)濟(jì)增長和農(nóng)民增加收入的新亮點[1-3]。南疆紅棗發(fā)展雖已初具規(guī)模,但發(fā)展歷程短,還存在很多問題,尤其是棗樹樹皮腐爛病發(fā)病越發(fā)嚴(yán)重[4],常常造成新栽幼樹大量死亡,棗樹受害后,樹勢衰弱,生長緩慢,結(jié)果晚,產(chǎn)量低,造成巨大的經(jīng)濟(jì)損失[5]。紅棗的種植格局是典型的綠洲農(nóng)業(yè),防護(hù)林成為果業(yè)生產(chǎn)的主要屏障,其中楊樹、胡楊、沙棗和柳樹是新疆防護(hù)林的主要樹種,因此明確棗樹腐爛病菌來源迫在眉睫。【前人研究進(jìn)展】杜琴等從新疆主林果樹木上采集腐爛病樣,經(jīng)鑒定為Cytospermaschulzeri.&P.Syd(有性型Valsamalicola)C.chrysoperma(有性型V.sordida)和C.sacculus(Schwein.)Gvrit(有性型V.ceratosperma.)(C.carphospermaFr.)[6-9]。病原菌由外向里逐漸侵染,使受害部位皮層壞死,切斷樹體營養(yǎng)運輸造成樹木死亡。【本研究切入點】關(guān)于棗樹腐爛病菌和其他林果樹木腐爛病菌有無交互侵染,尚未見報道。目前防護(hù)林腐爛病發(fā)生嚴(yán)重,是否是引起棗樹腐爛病的主要原因尚不明確。將來源于楊樹(PopulusbolleanaLauche)、沙棗樹(ElaeagnusangustifoliaL)、胡楊(Populuseuphratica)、梨樹(Pyrussorotina)、蘋果樹(M.pumilaMill)、棗樹(Zizyphusjujuba)、桃樹(Prunuspersica)、柳樹(Salixbabylonica)、核桃(J.regiaL)腐爛病菌株分別接種到30%棗樹樹皮培養(yǎng)基、果實、離體枝條以及田間健康枝條上,觀察病狀特點、產(chǎn)孢情況并計算發(fā)病率及病斑擴(kuò)展速率,明確棗樹腐爛病菌來源。【擬解決的關(guān)鍵問題】通過不同寄主來源腐爛病菌分離株接種在室內(nèi)棗樹離體組織和田間健康組織觀察其致病性,明確引起棗樹腐爛病病菌來源及不同寄主來源腐爛病菌致病力差異性,為棗園林木腐爛病綜合防治提供理論依據(jù)。
1 材料與方法
1.1 材 料
1.1.1 病原菌
腐爛病菌菌株,分離于阿克蘇地區(qū)的楊樹、沙棗、胡楊、梨樹、蘋果、棗樹、桃樹、柳樹、核桃樹發(fā)病枝條上。實驗前在PDA平板上25℃全光照培養(yǎng)3~4 d活化以備用[10]。表1
表1 不同寄主來源腐爛病菌分離株
Table 1 Source rot bacteria isolated from different host strains
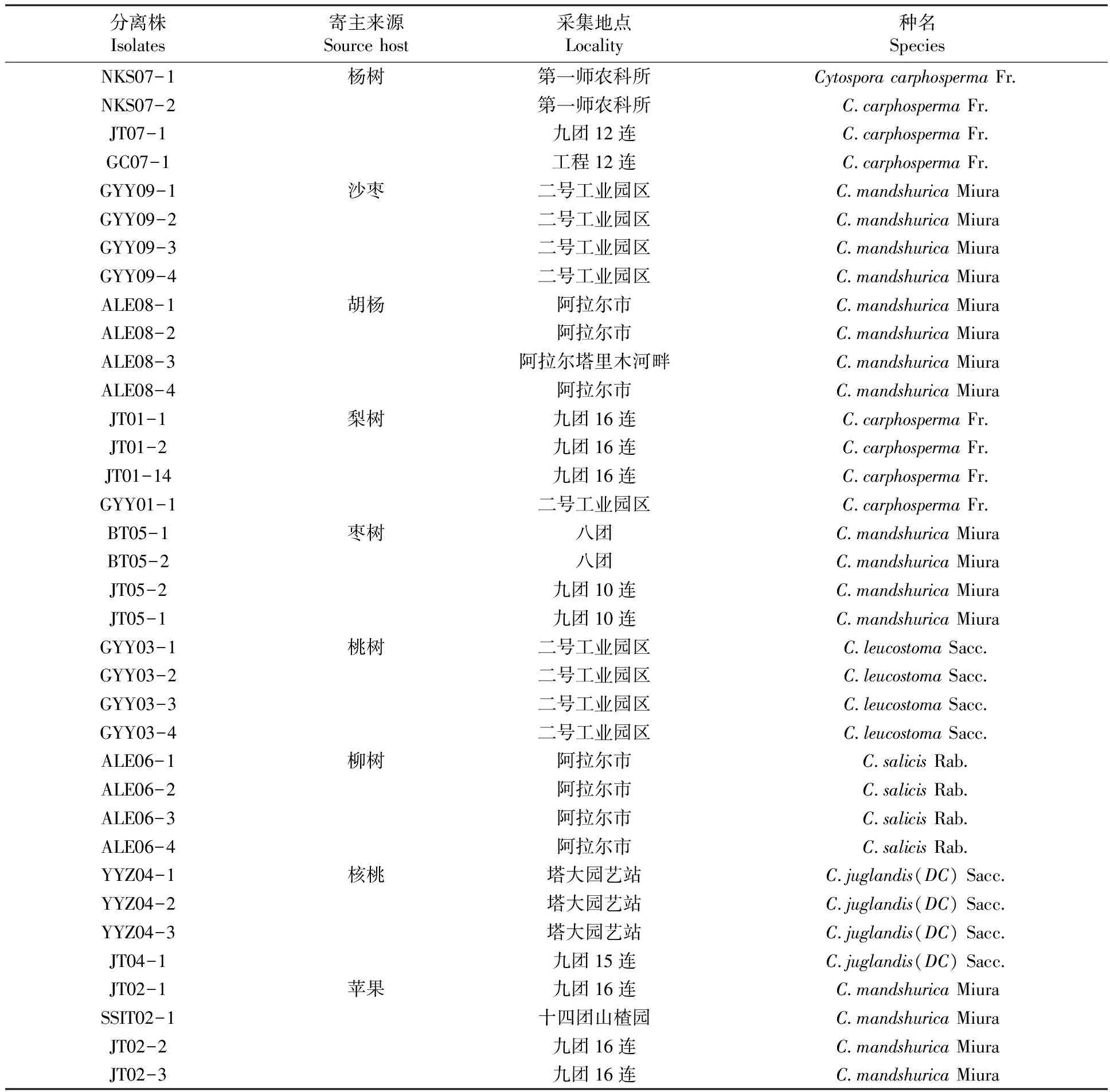
分離株Isolates寄主來源Sourcehost采集地點Locality種名SpeciesNKS07-1楊樹第一師農(nóng)科所CytosporacarphospermaFr NKS07-2第一師農(nóng)科所C carphospermaFr JT07-1九團(tuán)12連C carphospermaFr GC07-1工程12連C carphospermaFr GYY09-1沙棗二號工業(yè)園區(qū)C mandshuricaMiuraGYY09-2二號工業(yè)園區(qū)C mandshuricaMiuraGYY09-3二號工業(yè)園區(qū)C mandshuricaMiuraGYY09-4二號工業(yè)園區(qū)C mandshuricaMiuraALE08-1胡楊阿拉爾市C mandshuricaMiuraALE08-2阿拉爾市C mandshuricaMiuraALE08-3阿拉爾塔里木河畔C mandshuricaMiuraALE08-4阿拉爾市C mandshuricaMiuraJT01-1梨樹九團(tuán)16連C carphospermaFr JT01-2九團(tuán)16連C carphospermaFr JT01-14九團(tuán)16連C carphospermaFr GYY01-1二號工業(yè)園區(qū)C carphospermaFr BT05-1棗樹八團(tuán)C mandshuricaMiuraBT05-2八團(tuán)C mandshuricaMiuraJT05-2九團(tuán)10連C mandshuricaMiuraJT05-1九團(tuán)10連C mandshuricaMiuraGYY03-1桃樹二號工業(yè)園區(qū)C leucostomaSacc GYY03-2二號工業(yè)園區(qū)C leucostomaSacc GYY03-3二號工業(yè)園區(qū)C leucostomaSacc GYY03-4二號工業(yè)園區(qū)C leucostomaSacc ALE06-1柳樹阿拉爾市C salicisRab ALE06-2阿拉爾市C salicisRab ALE06-3阿拉爾市C salicisRab ALE06-4阿拉爾市C salicisRab YYZ04-1核桃塔大園藝站C juglandis(DC)Sacc YYZ04-2塔大園藝站C juglandis(DC)Sacc YYZ04-3塔大園藝站C juglandis(DC)Sacc JT04-1九團(tuán)15連C juglandis(DC)Sacc JT02-1蘋果九團(tuán)16連C mandshuricaMiuraSSIT02-1十四團(tuán)山楂園C mandshuricaMiuraJT02-2九團(tuán)16連C mandshuricaMiuraJT02-3九團(tuán)16連C mandshuricaMiura
1.1.2 接種材料
1.1.2.1 30%棗樹樹皮培養(yǎng)基
稱取棗樹樹皮300 g,用清水洗凈晾干,加蒸餾水1 000 mL,沸水煮30 min后過濾取上清液,加入蒸餾水補(bǔ)至1 000 mL,加瓊脂30 g,煮至瓊脂融化,pH自然,分裝至150 mL三角瓶中121℃,30 min高溫滅菌備用。
1.1.2.2 果實
摘取大小一致的未著色棗樹青果,經(jīng)清水沖洗3遍,用濾紙擦干,75%酒精進(jìn)行表面消毒,再用無菌水沖洗3遍后備用。
1.1.2.3 枝條
從田間采集2~3年生健康棗樹枝條,截成10 cm長的小段,先用洗潔精清洗,再用清水沖洗枝條3遍,晾干后75%酒精表面消毒,用規(guī)格5 mm打孔器滅菌后在枝條上進(jìn)行組織燙傷,之后用75%酒精擦拭一遍,再用無菌水沖洗3遍備用。
1.1.2.4 田間枝條選取
田間2~3年生的健康棗樹枝條,用規(guī)格5 mm打孔器消毒后在枝條上組織燙傷出圓形傷口,75%酒精擦拭一遍,再用無菌水沖洗三遍備用。
1.2 方 法
1.2.1 接 種
1.2.1.1 30%樹皮培養(yǎng)基
將不同寄主來源腐爛病菌株在PDA平板上25°C全光照條件下培養(yǎng)3 d后打成直徑5 mm的菌餅,分別接種于30%紅棗樹皮培養(yǎng)基上,每組3個重復(fù)。
1.2.1.2 果實組織燙傷法
用5 mm打孔器消毒滅菌后在果實上燙傷打孔,并將不同寄主來源腐爛病5 mm菌餅接種在傷口處,每個菌株接種6個棗果,接種好的果實放在滅菌的托盤中,托盤底部鋪有兩層無菌水潤濕的紗布用來保濕,最后用保鮮膜將托盤密封。
1.2.1.3 枝條組織燙傷法
用5 mm打孔器消毒滅菌后在酒精燈外焰灼熱后在枝條上打孔、接菌餅,接種好的枝條每兩支放在一個150 mL的三角瓶中,每個三角瓶內(nèi)加3 mL無菌水用來保濕,每種菌株接6個枝條,分裝3個三角瓶中并標(biāo)記。
1.2.1.4 田間枝條燙傷法
同室內(nèi)離體枝條實驗一樣用枝條組織燙傷法在田間健康枝條上打孔,接菌餅,滅菌的脫脂棉在無菌水中潤濕后取一小塊蓋在菌餅上,并用保鮮膜纏緊,以保證菌餅不會在侵染發(fā)病前干掉或脫落,每種菌6次重復(fù)。
1.2.2 培養(yǎng)和記錄
將接種好的30%棗樹樹皮培養(yǎng)基、果實、室內(nèi)離體枝條放在25°C全光照培養(yǎng)箱中培養(yǎng),每24 h觀察一次,并記錄菌絲生長速度、菌落形態(tài)、產(chǎn)孢時間及產(chǎn)孢量。同時對病狀特征進(jìn)行拍照。
接種好的田間枝條每隔24 h觀察記錄一次,計算發(fā)病率、病斑擴(kuò)展速度、病斑顏色、產(chǎn)孢情況等,對病狀特點進(jìn)行拍照。
1.2.3 測 定
1.2.3.1 采用十字交叉法測定病斑大小計算擴(kuò)展速度。
1.2.3.2 產(chǎn)孢量計算方法,參照郭仲眾等[11]采用區(qū)域分割法計數(shù)累計產(chǎn)孢量。
2 結(jié)果與分析
2.1 不同寄主來源腐爛病菌在30%棗樹樹皮培養(yǎng)基培養(yǎng)
培養(yǎng)3 d后,測量菌絲生長速率,來源棗樹和梨樹腐爛病菌生長速度最快大于2.30 cm/d,來源沙棗和柳樹腐爛病菌生長速度最慢小于2.00 cm/d,來源楊樹、蘋果和胡楊的病菌生長速度相差不大,7種不同寄主來源腐爛病菌在17 d內(nèi)均產(chǎn)孢,來源沙棗、胡楊、梨樹、蘋果、棗樹和柳樹腐爛病菌均在4~6 d內(nèi)形成產(chǎn)孢體,楊樹最晚在第17 d產(chǎn)孢,且形成產(chǎn)孢體數(shù)量和產(chǎn)孢大小存在差異。在第22 d觀察時,來源蘋果、沙棗樹腐爛病菌在樹皮培養(yǎng)基上吐出橘黃色分生孢子角,30 d時來源棗樹腐爛病菌在樹皮培養(yǎng)基上吐出橘黃色分生孢子角,來源其他樹木腐爛病菌始終未產(chǎn)生孢子角。表2
表2 不同寄主來源腐爛病菌在30%棗樹樹皮培養(yǎng)基上生長狀況
Table 2 The observation results of canker pathogens in the medium with 30% dates bark from different hosts

分離株Isolates寄主來源Sourcehost侵染速度Infectionrate(cm/d)產(chǎn)孢時間Sporulationtime(d)累積產(chǎn)孢CumulativeSpore(unit)產(chǎn)生分生孢子角時間Producesporescornertime(d)分生孢子大小Conidiasize(mm)菌落形態(tài)ColonymorphologyNKS07-1楊樹2 1017377—0 74~2 89菌絲乳白色、緊密、平伏、散射狀GYY09~1沙棗1 886321220 60~2 49菌絲乳白色、疏松、平伏、散射狀A(yù)LE08~1胡楊2 124413—0 60~3 18菌絲乳白色、疏松JT01-1梨樹2 334404—0 45~1 75菌絲緊密呈淡白色,散射狀生長JT02-1蘋果2 096442220 83~4 78菌絲生長疏松呈乳白色BT05-1棗樹2 306292300 73~2 25菌絲生長稀疏松散呈乳白色ALE06-1柳樹1 806216220 85~2 64菌絲生長稀疏CK——————無菌絲生長
注:—表示未產(chǎn)生分生孢子角
Note:—Indicates that no sporulation, did not produce spores angle
2.2 不同寄主來源腐爛病菌株侵染棗果實
在接種第3 d觀察9種腐爛病菌均能侵染棗果,其中來源胡楊和梨樹腐爛病菌在24 h內(nèi)顯癥,接種楊樹、梨樹和核桃腐爛病菌的果實腐爛最為嚴(yán)重,病斑呈褐色或深紅褐色占全果面積的2/3以上,病斑上有菌絲著生且病果向內(nèi)凹陷。接種沙棗、胡楊、棗樹和柳樹腐爛病菌的棗果病斑均呈褐色且腐爛面積較小占全果1/3以下,接種蘋果和桃樹腐爛病菌的棗果病斑面積占全果1/3~2/3;來源楊樹、沙棗、胡楊、棗樹腐爛病菌在棗果上形成產(chǎn)孢體,其他5種寄主來源腐爛病菌未形成產(chǎn)孢體。表3
表3 不同寄主來源腐爛病菌侵染棗果實觀察結(jié)果
Table 3 Different sources of host fruit rot pathogen infection observation dates
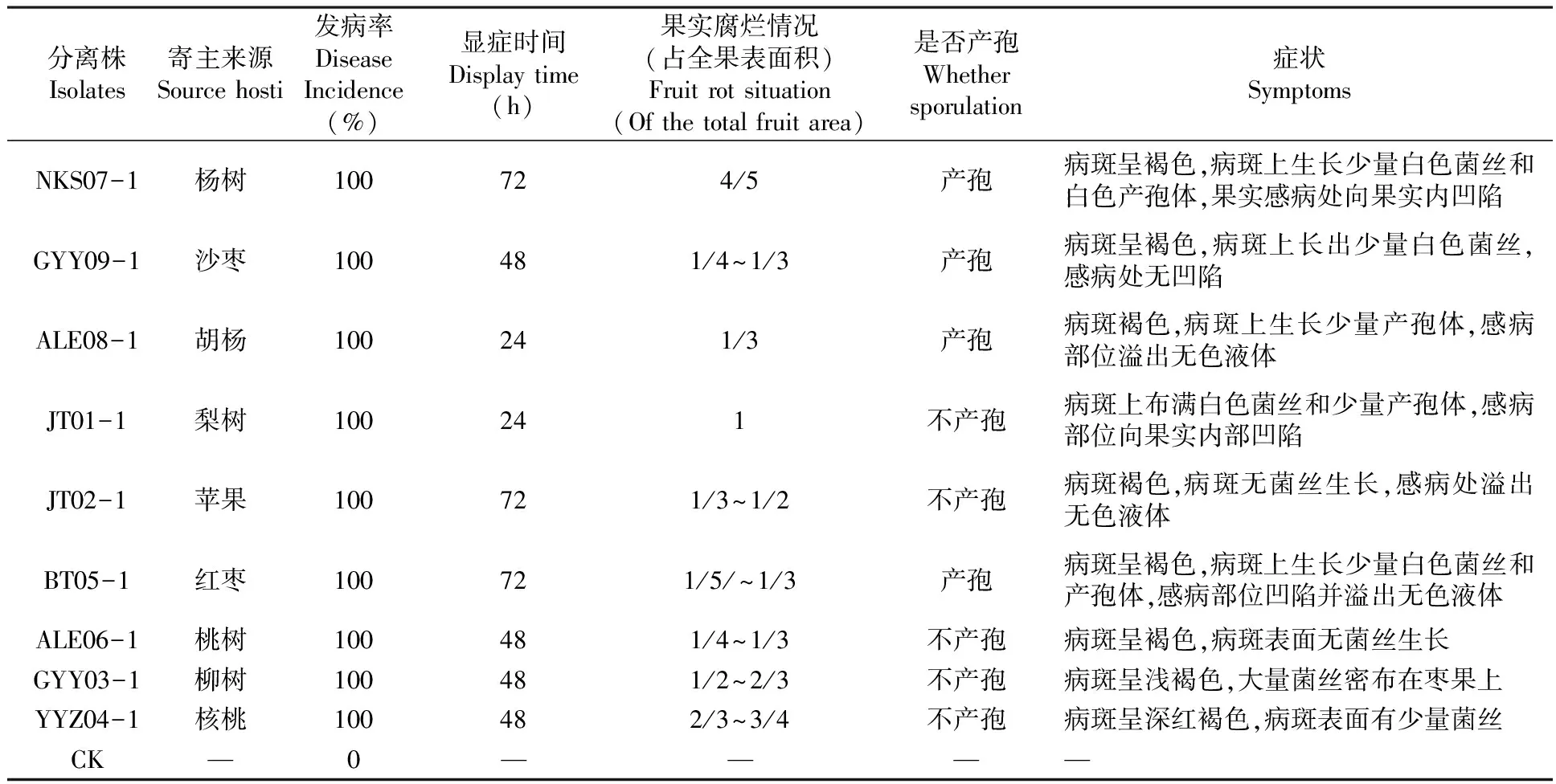
分離株Isolates寄主來源Sourcehosti發(fā)病率DiseaseIncidence(%)顯癥時間Displaytime(h)果實腐爛情況(占全果表面積)Fruitrotsituation(Ofthetotalfruitarea)是否產(chǎn)孢Whethersporulation癥狀SymptomsNKS07-1楊樹100724/5產(chǎn)孢病斑呈褐色,病斑上生長少量白色菌絲和白色產(chǎn)孢體,果實感病處向果實內(nèi)凹陷GYY09-1沙棗100481/4~1/3產(chǎn)孢病斑呈褐色,病斑上長出少量白色菌絲,感病處無凹陷ALE08-1胡楊100241/3產(chǎn)孢病斑褐色,病斑上生長少量產(chǎn)孢體,感病部位溢出無色液體JT01-1梨樹100241不產(chǎn)孢病斑上布滿白色菌絲和少量產(chǎn)孢體,感病部位向果實內(nèi)部凹陷JT02-1蘋果100721/3~1/2不產(chǎn)孢病斑褐色,病斑無菌絲生長,感病處溢出無色液體BT05-1紅棗100721/5/~1/3產(chǎn)孢病斑呈褐色,病斑上生長少量白色菌絲和產(chǎn)孢體,感病部位凹陷并溢出無色液體ALE06-1桃樹100481/4~1/3不產(chǎn)孢病斑呈褐色,病斑表面無菌絲生長GYY03-1柳樹100481/2~2/3不產(chǎn)孢病斑呈淺褐色,大量菌絲密布在棗果上YYZ04-1核桃100482/3~3/4不產(chǎn)孢病斑呈深紅褐色,病斑表面有少量菌絲CK—0————
2.3 不同寄主來源腐爛病菌在棗樹離體枝條上致病性
接種后的棗樹枝條在25°C全光照條件下培養(yǎng)觀察,不同寄主來源腐爛病菌株均能使棗樹枝條在72 h內(nèi)發(fā)病,來源同種寄主不同株系的腐爛病菌在顯癥時間、病斑擴(kuò)展速度、產(chǎn)孢時間等方面存在差異,其中來源楊樹腐爛病菌的4個分離株病斑擴(kuò)展速度最快,JT07-1為0.22 cm/d,最慢的是NKS07-1和NKS07-2為0.12 cn/d,且在第15 d時觀察到所產(chǎn)生的分生孢子角顏色有差異,來源NKS07-1和NKS-2產(chǎn)生淺黃的分生孢子角,來源JT07-1和GC07-1產(chǎn)生橘黃色分生孢子角。其中來源沙棗、胡楊、棗樹和柳樹腐爛病菌各個分離株病斑擴(kuò)展速度最快,均在0.2 cm/d以上。9種不同寄主來源腐爛病菌接種棗樹枝條產(chǎn)孢時間相差不大都在7~9 d,且在第15 d觀察其吐出淡黃色、黃色、橘黃色分生孢子角。表4
2.4 不同寄主來源腐爛病菌在田間棗樹枝條的致病性
不同寄主來源腐爛病菌株均能侵染田間健康的棗樹枝條,并在5 d內(nèi)使之發(fā)病。接種后第15 d測量病斑擴(kuò)展大小并計算病斑擴(kuò)展速度,其中來源胡楊的腐爛病菌病斑擴(kuò)展速度最快為0.20 cm/d,來源梨樹、楊樹和沙棗腐爛病菌株病斑擴(kuò)展速度較快,而來源紅棗腐爛病菌株病斑擴(kuò)展速度最慢為0.11 cm/d。來源梨樹和胡楊的腐爛病菌株侵染枝條的病斑處產(chǎn)生大量小黑點,并有黃色和橘黃色分生孢子角溢出。在第20 d觀察時發(fā)現(xiàn),胡楊和沙棗病斑明顯向內(nèi)凹陷。表5
表4 不同寄主來源腐爛病菌在棗樹枝條上的致病性測定
Table 4 Different host plants rot bacteria pathogenic measured on dates branches
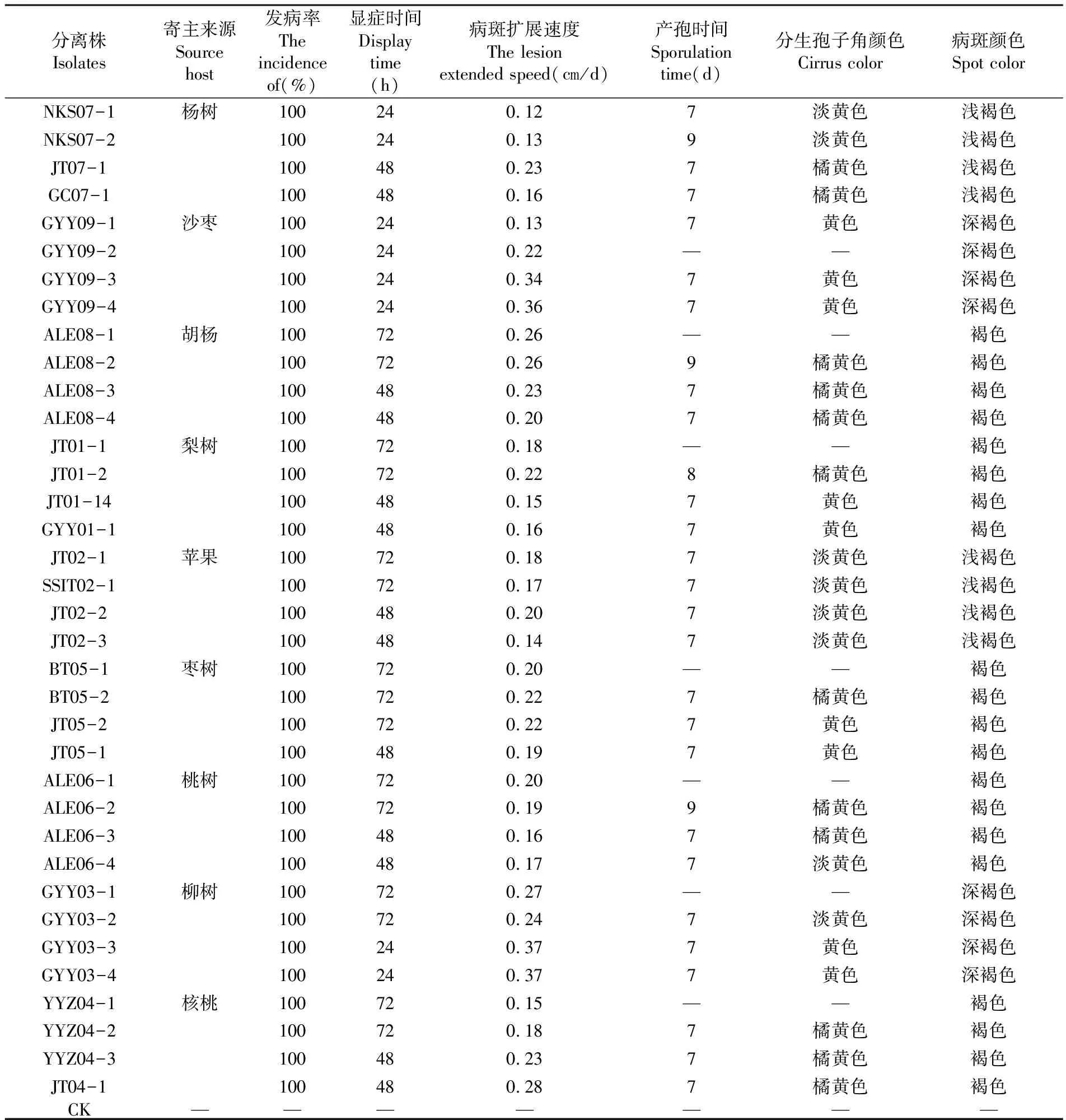
分離株Isolates寄主來源Sourcehost發(fā)病率Theincidenceof(%)顯癥時間Displaytime(h)病斑擴(kuò)展速度Thelesionextendedspeed(cm/d)產(chǎn)孢時間Sporulationtime(d)分生孢子角顏色Cirruscolor病斑顏色SpotcolorNKS07-1楊樹100240 127淡黃色淺褐色NKS07-2100240 139淡黃色淺褐色JT07-1100480 237橘黃色淺褐色GC07-1100480 167橘黃色淺褐色GYY09-1沙棗100240 137黃色深褐色GYY09-2100240 22——深褐色GYY09-3100240 347黃色深褐色GYY09-4100240 367黃色深褐色ALE08-1胡楊100720 26——褐色ALE08-2100720 269橘黃色褐色ALE08-3100480 237橘黃色褐色ALE08-4100480 207橘黃色褐色JT01-1梨樹100720 18——褐色JT01-2100720 228橘黃色褐色JT01-14100480 157黃色褐色GYY01-1100480 167黃色褐色JT02-1蘋果100720 187淡黃色淺褐色SSIT02-1100720 177淡黃色淺褐色JT02-2100480 207淡黃色淺褐色JT02-3100480 147淡黃色淺褐色BT05-1棗樹100720 20——褐色BT05-2100720 227橘黃色褐色JT05-2100720 227黃色褐色JT05-1100480 197黃色褐色ALE06-1桃樹100720 20——褐色ALE06-2100720 199橘黃色褐色ALE06-3100480 167橘黃色褐色ALE06-4100480 177淡黃色褐色GYY03-1柳樹100720 27——深褐色GYY03-2100720 247淡黃色深褐色GYY03-3100240 377黃色深褐色GYY03-4100240 377黃色深褐色YYZ04-1核桃100720 15——褐色YYZ04-2100720 187橘黃色褐色YYZ04-3100480 237橘黃色褐色JT04-1100480 287橘黃色褐色CK———————
注:—表示未形成產(chǎn)孢體
Note:—Stands for unformed
3 討 論
研究表明,不同寄主來源腐爛病菌分離株均能使棗樹不同部位感病與郭眾仲等[11]研究的6種不同寄主來源腐爛病菌株均能侵染庫爾勒香梨結(jié)果一致。依據(jù)柯赫氏法則鑒定獲得同種不同株系腐爛病菌純培養(yǎng)在顏色和菌落形態(tài)上有差異,與王旭麗等[12]梨樹腐爛病菌各分離株與蘋果腐爛病菌在菌落形態(tài)和產(chǎn)孢特點有一定差別,結(jié)果一致。梨樹和蘋果腐爛病在棗樹皮培養(yǎng)基上均形成產(chǎn)孢體與王旭麗等[13]研究結(jié)果基本一致,但產(chǎn)孢量和產(chǎn)孢大小存在差異。結(jié)果研究來源同種寄主的不同腐爛病菌分離株在致病性上存在差異與臧瑞等[14]研究結(jié)果一致。韋潔玲等[15]用不同致病力的蘋果腐爛病菌,通過燙傷法接種蘋果的不同組織,發(fā)現(xiàn)以嫩梢、果實作為接種材料時,測定結(jié)果與枝條做接種材料時結(jié)果一致,而研究通過燙傷法將不同寄主來源腐爛病菌各菌株接種棗果、離體枝條和田間枝條,測定結(jié)果不一致。在田間接種試驗中蘋果腐爛病菌接種紅棗枝條上30 d內(nèi)未發(fā)現(xiàn)肉眼可見的分生孢子器與陳曲[16]蘋果腐爛病接種一個月左右分生孢子器就可以形成研究結(jié)果不同,可能是由于寄主改變,菌株的致病力也發(fā)生改變或是由于菌株之間存在差異性。
表5 不同寄主來源腐爛病菌在田間棗樹枝條的致病性測定
Table 5 Different host plants rot bacteria pathogenicity test of dates branches in the field
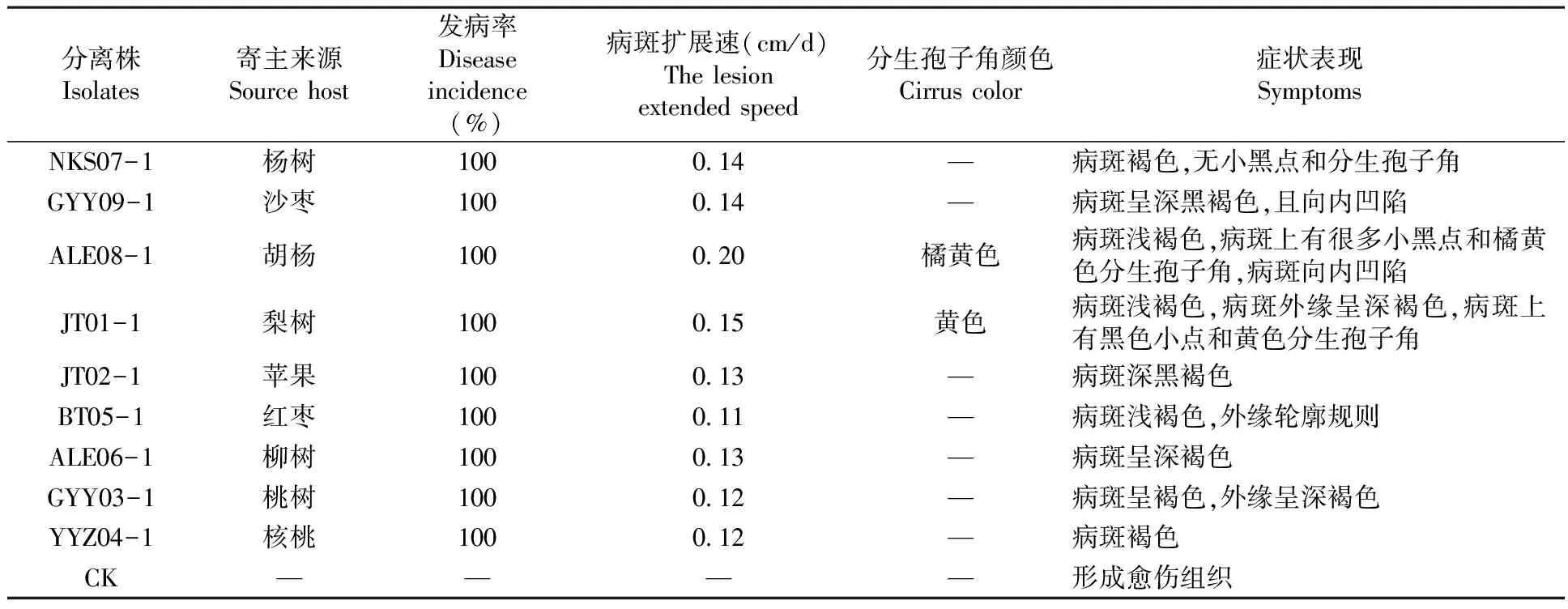
分離株Isolates寄主來源Sourcehost發(fā)病率Diseaseincidence(%)病斑擴(kuò)展速(cm/d)Thelesionextendedspeed分生孢子角顏色Cirruscolor癥狀表現(xiàn)SymptomsNKS07-1楊樹1000 14—病斑褐色,無小黑點和分生孢子角GYY09-1沙棗1000 14—病斑呈深黑褐色,且向內(nèi)凹陷ALE08-1胡楊1000 20橘黃色病斑淺褐色,病斑上有很多小黑點和橘黃色分生孢子角,病斑向內(nèi)凹陷JT01-1梨樹1000 15黃色病斑淺褐色,病斑外緣呈深褐色,病斑上有黑色小點和黃色分生孢子角JT02-1蘋果1000 13—病斑深黑褐色BT05-1紅棗1000 11—病斑淺褐色,外緣輪廓規(guī)則ALE06-1柳樹1000 13—病斑呈深褐色GYY03-1桃樹1000 12—病斑呈褐色,外緣呈深褐色YYZ04-1核桃1000 12—病斑褐色CK————形成愈傷組織
注:—表示未產(chǎn)生分生孢子角
Note:—Indicates that no produce spores angle
4 結(jié) 論
室內(nèi)離體測定中,不同寄主來源腐爛病菌均能在30%棗樹樹皮培養(yǎng)上擴(kuò)展并形成產(chǎn)孢體、使果實和枝條發(fā)病。9種不同寄主來源腐爛病的致病性存在差異,來源梨樹、棗樹、胡楊和楊樹腐爛病菌在30%棗樹樹皮培養(yǎng)基上菌絲生長速度較快,且形成產(chǎn)孢體較多,來源梨樹、楊樹、核桃腐爛病菌對棗果致病力較強(qiáng),來源棗樹、胡楊、沙棗和核桃腐爛病菌對室內(nèi)離體棗樹枝條致病力較強(qiáng),來源胡楊和梨樹腐爛病菌對田間健康枝條致病力較強(qiáng),且病斑上產(chǎn)生大量小黑點并吐出橘黃色分生孢子角。來源于梨樹、胡楊和棗樹腐爛病菌在室內(nèi)離體測定和田間健康枝條的測定中發(fā)現(xiàn),較之其他6種腐爛病菌致病力強(qiáng)。
References)
[1]石國強(qiáng). 新疆兵團(tuán)紅棗產(chǎn)業(yè)發(fā)展戰(zhàn)略研究[D]. 石河子:石河子大學(xué)碩士論文,2013.
SHI Guo-qiang. (2013).ResearchonJujubeIndustryDevelopmentStrategyofXinjiangProduction&ConstructionCorps[D].Master Dissertation. Shihezi University,Shehezi. (in Chinese)
[2]金新文,賈文婷,鄭霞,等. 新疆南疆地區(qū)紅棗產(chǎn)業(yè)營銷現(xiàn)狀與戰(zhàn)略思考[J]. 新疆農(nóng)墾經(jīng)濟(jì),2014,(1):37-39.
JIN Xin-wen, JA Wen-ting, ZHENG Xia, et al. (2014). Red jujube industry in Southern Xinjiang area of marketing present situation and the strategic thinking [J].XinjiangStateFarmsEconomy, (1):37-39. (in Chinese)
[3]姜羽晗,王志彬. 新疆南疆紅棗產(chǎn)業(yè)發(fā)展存在的問題及對策-以阿克蘇為例[J]. 新疆農(nóng)墾經(jīng)濟(jì),2012,12:36-38.
JIANG Yu-han,WANG Zhi-bin. (2012). Southern Xinjiang red jujube industry development problems and countermeasures, -take Aksu for example [J].XinjiangStateFarmsEconomy, (12):36-38. (in Chinese)
[4]海力帕爾·吾麥爾. 新疆果樹腐爛病的發(fā)生及防治[J]. 河北農(nóng)業(yè)科技,2008,13:30.
Hailipaer Wuermai. (2008). The occurrence and prevention of fruit rot disease in Xinjiang [J].HebeiAgriculturalScienceandTechnology, (13):30. (in Chinese)
[5]杜琴. 新疆主要林木腐爛病菌種類鑒定及其防治方法研究[D]. 石河子:石河子大學(xué)碩士論文,2013.
DU Qin. (2013).IdentificationtoPathogenofMainWoodsCankerinXinjianganditsControlTechnologyStudy[D]. Master Dissertation. Shihezi University,Shehezi. (in Chinese)
[6]唐俊煜,張王斌,李亞鵬, 等.庫爾勒香梨樹腐爛病菌生物學(xué)特性與致病性的初步研究[J]. 新疆農(nóng)業(yè)科學(xué),2015,52(2):252-257.
TANG Jun-yu, ZHANG Wang-bin, LI Ya-peng, et al. (2015). Korla Pear Tree Biological Characteristics of the Valsa Canker Growth Pathogen and Its Pathogenicity [J].XinjiangAgriculturalSciences,52(2):252-257. (in Chinese)
[7]陸燕君. 梨樹腐爛病病原菌的研究[J].植物病理學(xué)報. 1992, 22(3): 197-204.
LU Yan-jun. (1992). Studies on the pathogenic fungus of pear canker disease [J].JournalofPlantPathology, 22(3): 197-204. (in Chinese)
[8]郭靖,張王斌,張琦,等. 五種殺菌劑對庫爾勒香梨腐爛病的防效[J]. 北方園藝,2014,(11):103-105.
GUO Jing, ZHANG Wang-bin, ZHANG Qi, et al. (2014). Control effect of five fungicides rot disease prevention on Korla pear [J].NorthernHorticulture, (11):103-105. (in Chinese)
[10]方中達(dá).植病研究方法[M].北京:農(nóng)業(yè)出版社,1979:330-344.
FANG Zhong-da. ((1979).Researchmethodsofplantpathology[M]. Beijing: China Agriculture Pres:330-344. (in Chinese)
[11]郭眾仲,張王斌,李亞鵬, 等. 不同寄主來源腐爛病菌對庫爾勒香梨樹致病性研究[J]. 新疆農(nóng)業(yè)科學(xué),2014,51(12):2 245-2 250.
GUO Zhong-zhong, ZHANG Wang-bin, LI Ya-peng, et al. (2014). Study on the Pathogenicity of Canker Pathogen from Different Hosts to Korla fragrant pear Tree [J].XinjiangAgriculturalSciences, 51(12):2,245-2,250. (in Chinese)
[12]王旭麗,康振生,黃麗麗, 等.ITS序列結(jié)合培養(yǎng)特征鑒定梨樹腐爛病菌[J].菌物學(xué)報,2007,26 (4):517-527.
WANG Xu-li, KANG Zhen-sheng, HUANG Li-li, et al. (2007). Cultivating ITS sequence characterization pear tree Valsa Mali [J].JournalofFungi, 26(4): 517-527. (in Chinese)
[13]王旭麗,臧睿,王磊, 等. 蘋果樹Valsa malicola的發(fā)現(xiàn)及其致病性研究[J]. 林業(yè)科學(xué),2007,(9):23-26,156.
WANG Xu-li, ZANG Rui, WANG Lei,et al. (2007). Study on the pathogenicity and Valsa malicola found that apple tree [J].ForestryScience, (9): 23-26,156. (in Chinese)
[14]臧睿,黃麗麗,康振生,等.陜西蘋果樹腐爛病菌(Cytospora spp.)不同分離株的生物學(xué)特性與致病性研究[J].植物病理學(xué)報,2007,(4):344-351.
ZANG Rui, ,HUANG Li-li, KANG Zheng-sheng, et al. (2007).Biological characteristics and pathogenicity of different isolates of Cytospora spp.isolated from apple terees in Shannxi Province (China) [J].JournalofPlantPathology, (4):344-351. (in Chinese)
[15]韋潔玲. 蘋果樹腐爛病菌不同分離株致病性差異研究[D].楊凌:西北農(nóng)林科技大學(xué)碩士論文,2009.
WEI Jie-ling. (2009).Studyonthepathogenictyofdifferentisolatesofappletreevalsacankerpathogen[D].Master Dissertation.North West Agriculture and Forestry University, Yangling. (in Chinese)
[16]陳曲. 蘋果樹腐爛病病斑擴(kuò)展及分生孢子形成和釋放規(guī)律研究[D].保定:河北農(nóng)業(yè)大學(xué)碩士論文,2011.
CHEN QU. (2011).StudyonthesporeformationandreleasepatternofValsaceratosperma,andthelesionexpansiondynamicsofappletreecanker[D]. Master Dissertation. Agricultural University of Hebei Province, Baoding.(in Chinese)
Fund project:Supported by Regional Science Funds of NSFC (31460471), 2013 National College and University Students Innovational and Entrepreneurial Training Program(TDGCX201215;201310757002)
Study on the Pathogenicity ofCytosporaspp. from Different Hosts to Jujube
LI Chun-yan1, ZHANG Wang-bin1,2,GOU Qiao1, LIU Zhen-ya1,YAO Yong-sheng1, DAN Hong-xia1
(1.CollegeofPlantScience,TarimUniversity,AlarXinjiang843300,China; 2.KeyLaboratoryofComprehensiveManagementofAgriculturalPestsinSouthernXinjiang,TarimUniversity,AlarXinjiang843300,China)
【Objective】 To clarify the source of Chinese jujube rot pathogen in order to provide the main basis for the comprehensive prevention and control of production.【Method】 Different source host rot pathogens were inoculated in 30% jujube bark media, and fruit, branches and field inoculated branches were observed and the incidence, disease spot extending speed and spore production, etc. were calculated.【Result】 In 30% jujube tree bark medium, different source host rot pathogens produced spore production within 7 days; Rot pathogen from willow, apple, elaeagnus angustifolia, jujube rot pathogen within 30 d successively produced orange conidium angle; Rot pathogen from elaeagnus angustifolia, jujube, peuphratica and poplar could spread fast spores on the jujube fruit; Rot pathogen from branches in the field derived from Euphrates poplar inoculation disease spot expanded the fastest. Rot pathogen on branches inoculated with euphratica and pear trees produced small black spots in 15 days and spitted out yellow conidium angles.【Conclusion】 Different source host rot pathogens can be bred in 30% jujube tree bark medium and form spores, and produce conidium angle; they fully infect jujube with the in vitro and the healthy branches, but great difference exist in different source host rot pathogens.
jujube;Cytosporaspp.; virulence; spread velocity; gleba
10.6048/j.issn.1001-4330.2016.01.007
2015-05-30
國家自然基金地區(qū)科學(xué)基金“香梨優(yōu)斑螟與腐爛病協(xié)同為害的化學(xué)生態(tài)機(jī)制”(31460471);國家級大學(xué)生創(chuàng)新創(chuàng)業(yè)訓(xùn)練計劃(TDGCX201215;201310757002)
李春艷(1992-),女,新疆石河子人,本科生,研究方向為植物病理學(xué),(E-mail)387863197@qq.com
張王斌(1974-),男,陜西澄城人,副教授,研究方向為林果樹木腐爛病,(E-mail)zwbzky@163.com
S436.65
A
1001-4330(2016)01-0051-08
