‘豐水’梨二次開花過程中枝皮和花芽有機營養變化特征
2016-05-06 03:48:08張全軍鐘必鳳李文貴鄧家林張紹鈴
西北植物學報 2016年3期
張全軍,鐘必鳳,李文貴,鄧家林,張紹鈴
(1 四川省農業科學院園藝研究所/農業部西南地區園藝作物生物學及種質創制重點實驗室,成都 610066;2 南京農業大學 梨工程技術研究中心,南京210095)
?
‘豐水’梨二次開花過程中枝皮和花芽有機營養變化特征
張全軍1,鐘必鳳1,李文貴1,鄧家林1,張紹鈴2
(1 四川省農業科學院園藝研究所/農業部西南地區園藝作物生物學及種質創制重點實驗室,成都 610066;2 南京農業大學 梨工程技術研究中心,南京210095)
摘要:為了探討砂梨二次開花與枝皮和花芽營養之間的關系,分析了‘豐水’梨去葉后二次開花過程中枝皮和花芽有機營養的變化。結果顯示:(1)‘豐水’梨從去葉后第6天出現萌芽現象,在第2周萌芽率迅猛增長,至第42天萌芽率增至91%,滿樹呈春天開花現象。(2)去葉后初期‘豐水’梨枝皮中可溶性糖含量逐漸增加,并于第12天時達到高峰后迅速下降,且枝皮中可溶性糖和淀粉含量迅速降低的時期與花芽萌芽率快速提高期同步;去葉后枝皮中可溶性蛋白質含量急劇降低,而花芽中游離氨基酸的含量迅速增加。(3) ‘豐水’梨花芽萌芽率與枝皮、花芽中的淀粉和可溶性蛋白質含量呈極顯著或顯著負相關關系,而與花芽中可溶性糖含量呈顯著正相關關系。研究表明,砂梨枝皮、花芽中有機營養物質及其活躍狀態是影響其花芽萌動的重要原因。
關鍵詞:砂梨;二次花;營養條件;可溶性糖;淀粉;可溶性蛋白
按照正常的物候期,在中國梨開花期應該是3~5月份,但在南方地區當年形成的花芽在8~10月會出現異常開花的現象[1-2],生產上稱為‘二次開花’,該現象在新西蘭、南非、巴西被稱為‘芽敗育’(bud abortion)[3]或‘跳芽’(bud jump)[4-5]。砂梨二次開花會消耗樹體大量營養,導致花量少、花質差、著果少、果實小,雖然也能形成少量果實,但隨著氣溫的下降,果實逐漸停止生長,最后脫落,失去經濟價值。目前中國南方地區幾乎所有低海拔早熟梨產區都存在著秋季二次開花的現象,造成的損失在15%~30%,個別果園在發生較重的年份達70%以上。黃新忠等[6]認為砂梨二次開花與早期落葉緊密相關;鐘必鳳等[1]的試驗結果表明,早期落葉降低了砂梨花芽中ABA的含量,從而使激素比值發生改變,導致生長促進型激素占主導地位,阻止自然休眠的花芽進入休眠或促使進入淺休眠部分花芽解除休眠,所以出現砂梨二次開花現象。趙志國等[7]認為砂梨二次花的發生與該現象發生前2~4個月內樹體無機營養元素水平無直接關系。但迄今關于樹體有機營養與砂梨二次開花關系的研究未見報道。
果樹萌發期間需用的養料與能量主要來自它的貯藏物質的轉化和利用;其貯藏物質主要有蛋白質、淀粉和脂肪。在完成花芽分化之后的休眠期內,樹體內的營養物質發生轉變,如細胞液濃度提高,原生質粘性增強,原生質膜形成擬脂層使透性減弱等[8]。這些生理變化一方面有利于果樹防止失水和提高抗逆性,另一方面積極為儲備再生長準備物質基礎[9]。鑒于砂梨二次開花與早期落葉的緊密關系[10],本研究采取100%去葉處理‘豐水’梨促花措施,探討其二次開花過程中相關營養生理指標的變化及其與砂梨花芽萌動、二次開花的關系,期望對砂梨二次開花的調控提供重要的借鑒。
1材料和方法
1.1材料和試驗地
本試驗于2012年在南京農業大學江浦農場進行,試驗品種為8年生‘豐水’梨,試驗樹生長結果正常,長勢和立地條件基本一致。根據南京氣象局的資料,南京屬亞熱帶季風氣候,雨量充沛,年降水1 200 mm,四季分明,年平均溫度15.4 ℃,年極端氣溫最高39.7 ℃,最低-13.1 ℃,年平均降水量1 106 mm。
1.2方法
1.2.1花芽分化時期的觀察于6月10日開始,每隔5~10 d取均勻一致的花芽,每次取樣均為20個芽。對部分芽樣進行剝芽并統計鱗片數,采用半薄切片觀察,并制作石蠟切片。
1.2.2試驗處理在‘豐水’梨花芽分化完成后,即‘豐水’梨采果后的8月20日,進行全樹人工100%去葉處理,以不去葉為對照,5株小區,4次重復。每6 d取1次混合芽和一年生枝皮進行測試分析。
1.2.3測定項目及方法枝條萌芽率從開始處理后的42 d內每隔6 d統計1次;萌芽率=萌芽數/枝條總芽數×100%。參考《植物生理實驗指導》[11],淀粉含量采用蒽酮比色法測定;淀粉酶活性的測定 采用3,5-二硝基水楊酸比色法;可溶性糖含量采用蒽酮比色法測定;氨基酸含量采用茚三酮比色法測定;可溶性蛋白質含量采用考馬斯亮藍法測定。
1.3數據處理
實驗數據處理采用Excel軟件完成,差異性顯著性檢驗采用DPS軟件完成。
2結果與分析
2.1‘豐水’梨花芽分化時期和二次開花現象觀察
2.1.1花芽分化從6月上旬開始,通過半薄切片和石蠟切片的觀察,明確了‘豐水’梨花芽形態分化始于6月中、下旬,生長點于7月上旬開始膨大隆起,接著花序的小花原基開始分化,為花芽分化發端期;7月下旬萼片原基形成開始,為原基分化期;8月上旬花芽分化基本完成(圖1)。
2.1.2枝條萌芽率如圖2所示,在南京試驗點自然氣候條件下,‘豐水’梨從去葉后第6天開始出現萌芽現象,并在隨后2周內萌芽率迅猛增長,出現二次開花現象;從第24天起萌芽率增速減緩,到第42天萌芽率增至91%,滿樹呈春天開花現象。同時,相對來講,未去葉處理的‘豐水’梨在第12天出現零星萌芽,隨著秋季氣溫的延續,到第42天萌芽率僅12.3%,雖然其萌芽率動態變化與去葉處理‘豐水’梨保持相似的趨勢,但萌芽率一直保持在相對較低的水平。這可能與未去葉‘豐水’梨保持正常的營養生長狀態,抑制了生殖生長有關。
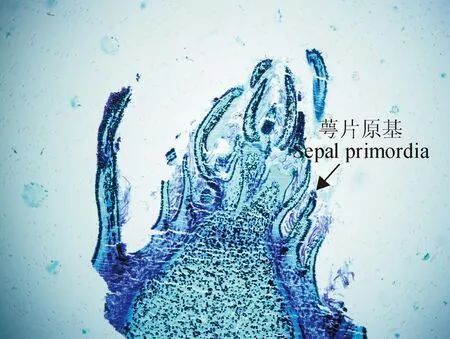
圖1 8月15日‘豐水’梨花芽的石蠟切片(×40)
2.2‘豐水’梨二次開花過程中枝皮和花芽生理特征變化
2.2.1枝皮和花芽淀粉含量圖3顯示,在去葉后的整個萌芽二次開花過程中(0~42 d),‘豐水’梨去葉處理的枝皮淀粉含量呈遞減趨勢,并且一直低于未去葉對照,淀粉含量最低值僅僅為0.03 mg·g-1;而未處理枝皮淀粉含量在第0~12天呈緩慢遞增趨勢,在第12天后緩慢減少,最后淀粉含量穩定在0.08 mg·g-1左右。在去葉后24 d,經去葉處理的‘豐水’梨大部分花芽已經呈萌動開花狀態,所以花芽中淀粉含量停止取樣分析。圖3顯示,未去葉對照花芽中淀粉含量在第6天后呈緩慢遞增趨勢,由0.072 mg·g-1增至0.083 mg·g-1,但于0~24 d維持在0.05~0.08 mg·g-1范圍內,明顯低于同期對照的枝皮淀粉含量(0.09~0.14 mg·g-1);與枝皮中淀粉含量變化相似,去葉處理的花芽淀粉含量也表現出逐漸降低趨勢,且始終低于同期的對照處理。可見,去葉處理不同程度地降低了‘豐水’梨枝皮和花芽淀粉含量,且去葉時間越長降幅越大。
2.2.2枝皮淀粉酶活性如圖4 所示,在去葉處理后的第6天,去葉處理‘豐水’梨枝皮α-淀粉酶活性先由1.1 mg·g-1·min-1增至1.6 mg·g-1·min-1,而后呈遞減狀態,于第18天降至1.2 mg·g-1·min-1,而后略微升高,此后持續降低,又在第42天降低至0.6 mg·g-1·min-1;同時,在整個去葉處理后階段(0~42 d),對照枝皮α-淀粉酶活性低于同期的去葉處理,且一直穩定在0.8~1.1 mg·g-1·min-1之間,起伏波動不大。在‘豐水’梨去葉后的過程中,去葉處理枝皮β-淀粉酶活性大體上均呈現先增加再降低、略呈平穩狀態后又升高的趨勢(圖4);而對照枝皮β-淀粉酶活性在0~42 d內呈遞減趨勢,由3.4 mg·g-1·min-1緩慢降至2.0 mg·g-1·min-1,變化較平穩。總之,在整個觀察過程中,‘豐水’梨枝皮中2種淀粉酶活性在未去葉處理下波動變化的幅度較小,而在去葉處理下變化較明顯。
2.2.3枝皮和花芽可溶性糖含量在‘豐水’梨去葉處理后二次開花過程中,去葉處理枝皮中可溶性糖含量呈現先略微增加又持續降低的變化趨勢,并在去葉處理第42天降至0.1 mg·g-1,而對照枝皮中可溶性糖含量雖也呈緩慢降低趨勢,但一直保持在0.22~0.25 mg·g-1,呈較穩定狀態(圖5)。同時,在整個去葉后二次開花過程中,對照花芽中可溶性糖含量顯著低于同期的去葉處理,且穩定保持在0.10~0.12 mg·g-1之間,而去葉處理可溶性糖含量先在第6天緩慢遞增,于第12天增至0.16 mg·g-1,隨后保持在平穩狀態(圖5)。可見,去葉處理引起‘豐水’梨枝皮和花芽中可溶性糖含量明顯波動,但枝皮中有降低趨勢,而花芽中則呈上升趨勢,且枝皮中波動幅度更大。
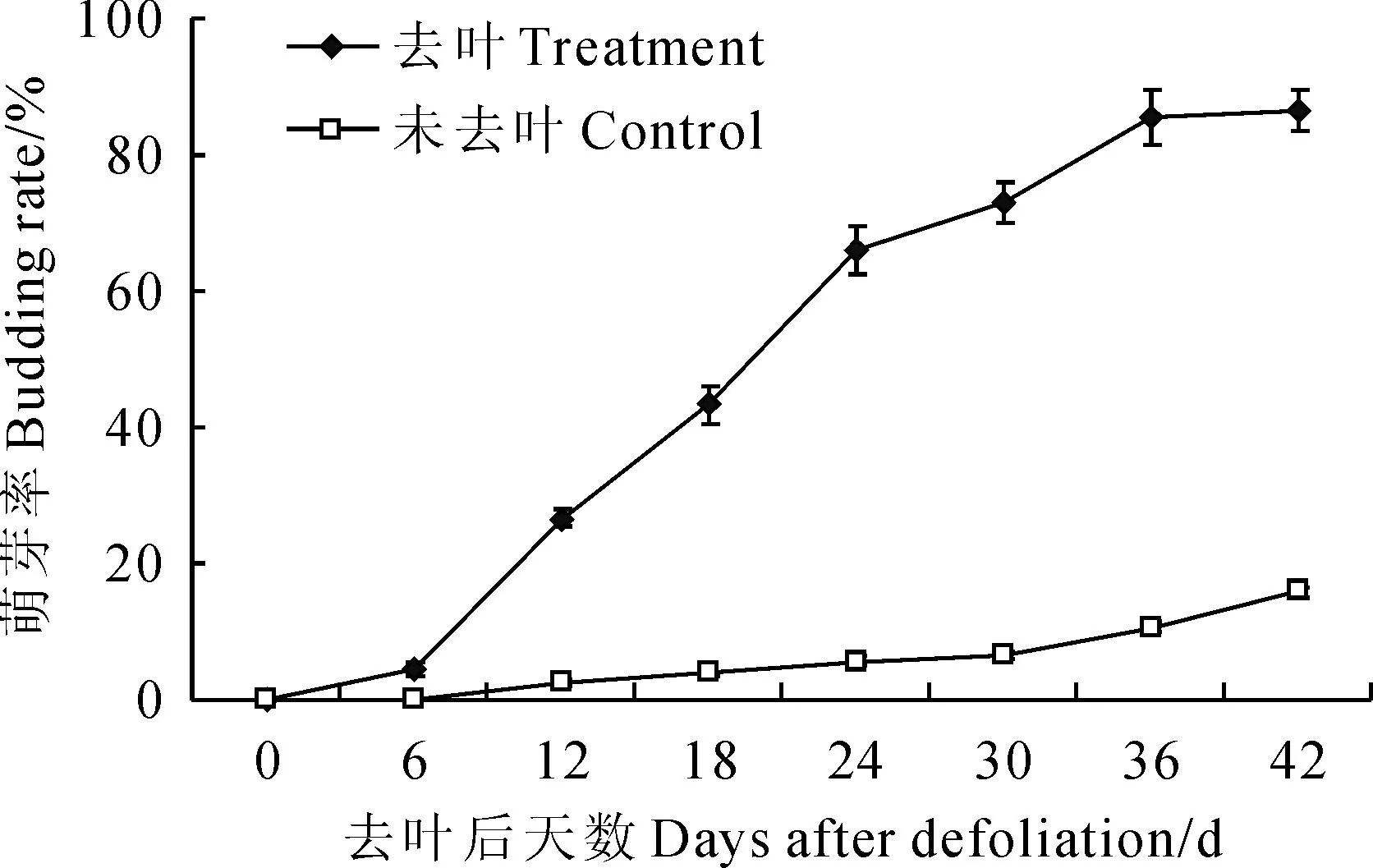
圖2 去葉后‘豐水’梨萌芽率動態變化

圖3 去葉后‘豐水’梨枝皮和花芽內淀粉含量變化
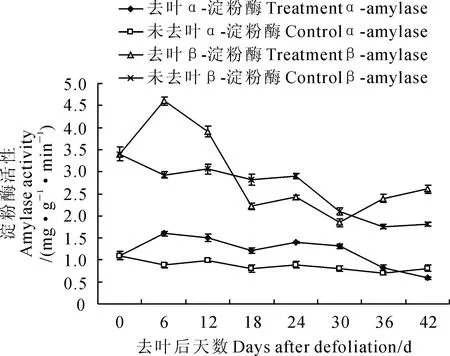
圖4 去葉處理后‘豐水’梨枝皮α-淀粉酶和
2.2.4枝皮和花芽游離氨基酸含量由圖6 可以看出:在去葉后‘豐水’梨萌芽二次開花過程中,去葉處理‘豐水’梨枝皮游離氨基酸含量始終低于同期的對照,且呈現先明顯降低后緩慢增加趨勢,并處理后第12天降低到最低值(1.60 mg·g-1),于第42天又緩慢增至2.21 mg·g-1。從整體來看(圖6),‘豐水’梨花芽中游離氨基酸含量低于枝皮;去葉處理花芽中游離氨基酸含量始終高于同期的對照,其中的對照花芽游離氨基酸含量波動在2.30~2.45 mg·g-1之間,而去葉處理花芽游離氨基酸含量從第6天開始緩慢上升,至第42天達最高值(2.68 mg·g-1)。可見,去葉處理明顯降低了‘豐水’梨枝皮氨基酸含量,而花芽中氨基酸含量增高。
2.2.5枝皮和花芽可溶性蛋白含量在去葉后萌芽二次開花整個過程中,去葉處理‘豐水’梨枝皮可溶性蛋白質含量始終低于對照,且差距逐漸增大(圖7)。其中,對照枝皮可溶性蛋白質含量呈緩慢遞減趨勢,并在第18天降至谷值(0.13 mg·g-1),隨后緩慢上升,到第42天增至0.19 mg·g-1;而去葉處理枝皮可溶性蛋白質含量一直呈遞減趨勢,至第42天時減至0.07 mg·g-1。同時,在花芽萌動過程中(圖7),無論是對照還是去葉處理,花芽中可溶性蛋白質含量都呈緩慢遞減趨勢,去葉處理可溶性蛋白質含量遞減速度高于同期的對照,其值始終低于對照,到去葉第24天降至最低值(0.075 mg·g-1)。可見,去葉處理明顯降低了‘豐水’梨枝皮和花芽中可溶性蛋白含量,且枝皮中下降的幅度更大。
2.3二次開花過程中萌芽率與枝皮、花芽營養相關指標的關系
對‘豐水’梨二次開花過程中萌芽率與枝皮和花芽營養指標進行相關性分析的結果(表1、2)表明,‘豐水’梨萌芽率與枝皮淀粉含量和可溶性蛋白含量呈極顯著的負相關關系(r分別為-0.931和-0.934),與可溶性糖含量、β-淀粉酶活性呈顯著的負相關關系,與花芽中淀粉、可溶性蛋白含量亦呈顯著的負相關,而與花芽中可溶性糖呈正相關關系。說明枝皮營養物質及其活躍狀態是影響砂梨花芽萌動的重要原因。

圖5 去葉處理后‘豐水’梨枝皮和花芽可溶性糖含量變化
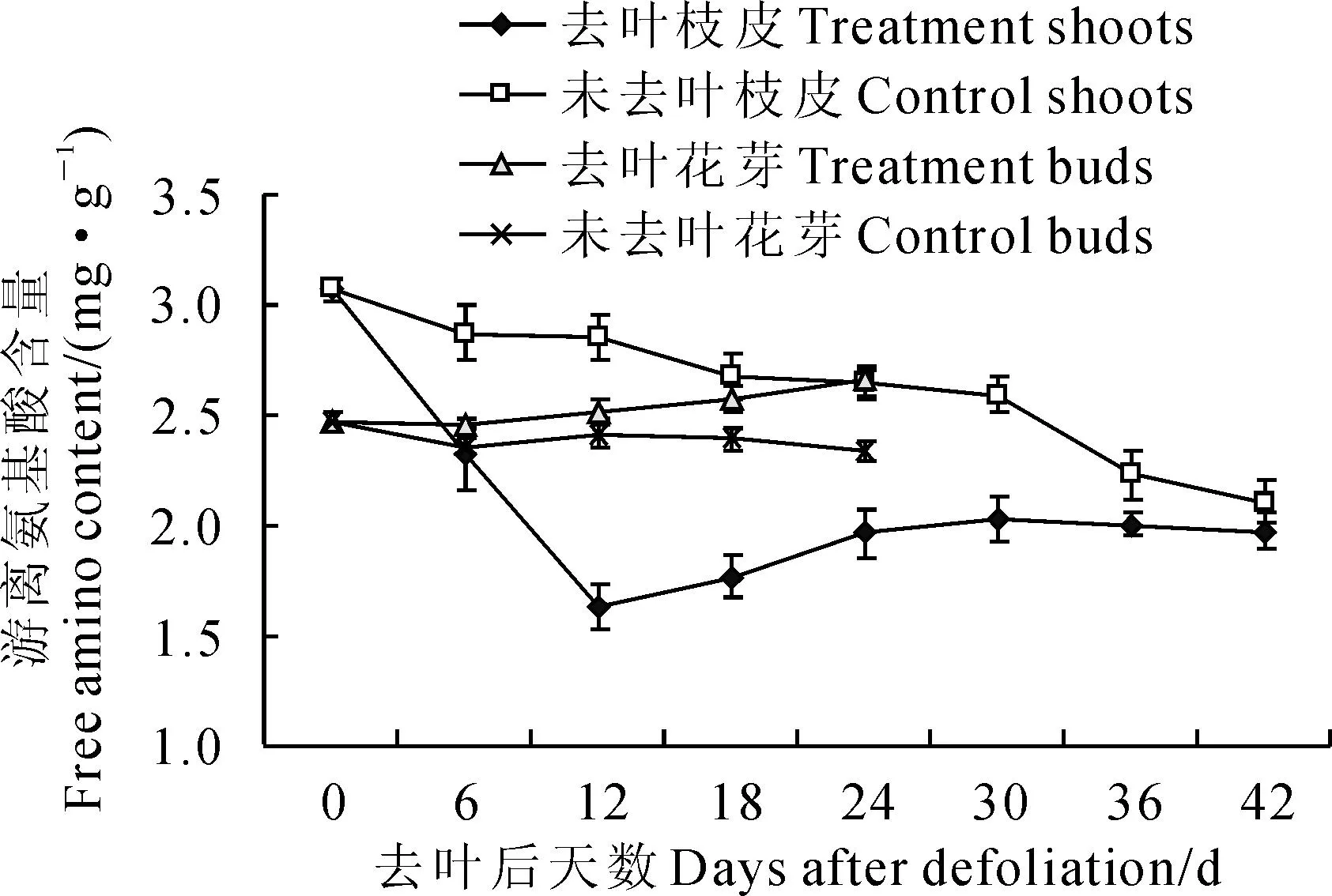
圖6 去葉處理后‘豐水’梨枝皮和花芽游離氨基酸含量變化
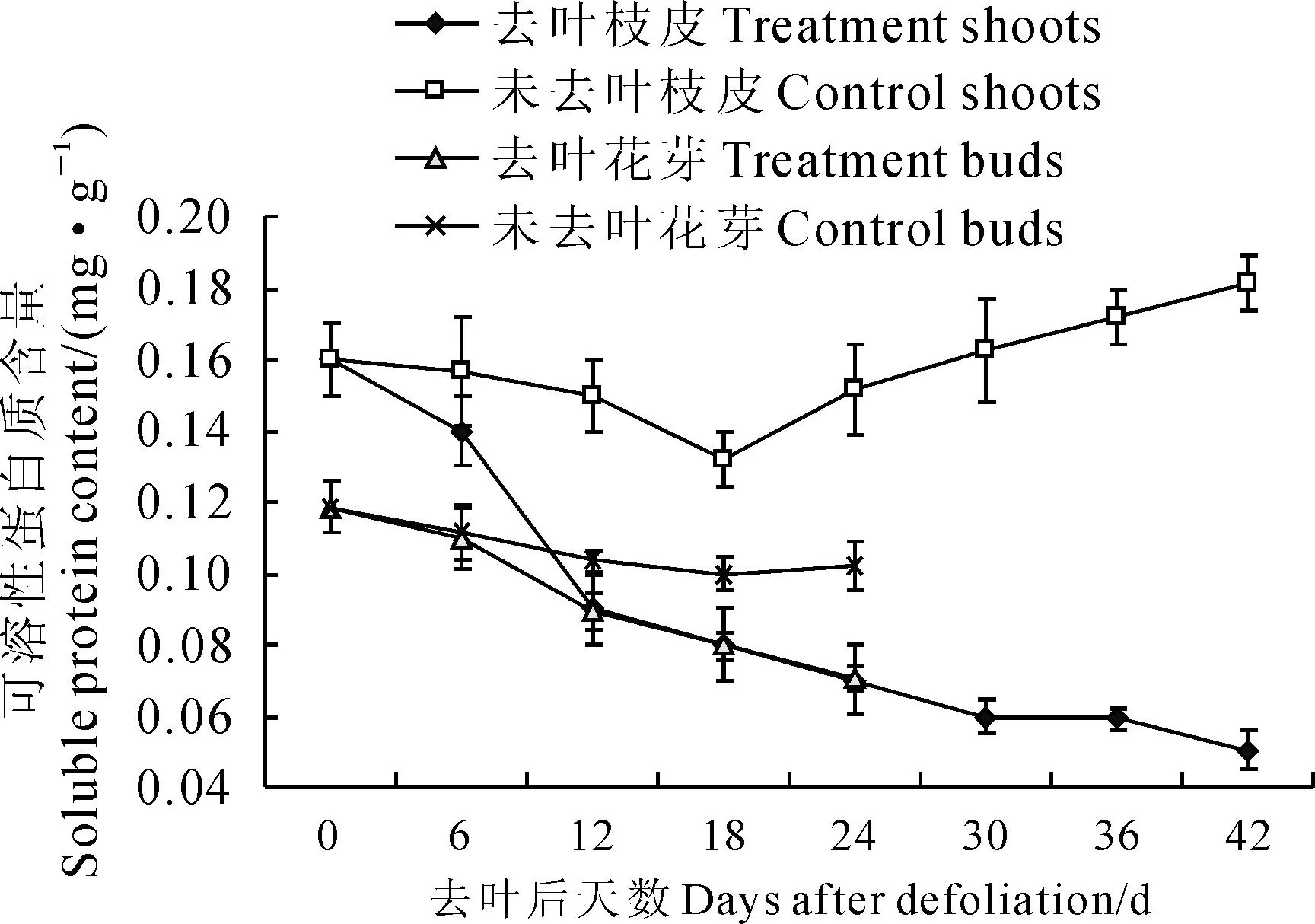
圖7 去葉處理后‘豐水’梨枝皮和花芽中蛋白質含量變化
同時,砂梨枝皮β-淀粉酶活性與淀粉含量、可溶性蛋白和游離氨基酸含量之間的相關系數均達到極顯著水平,這些證明在砂梨花芽萌動過程中,枝皮和花芽中貯藏營養成分之間存在的轉換關系。
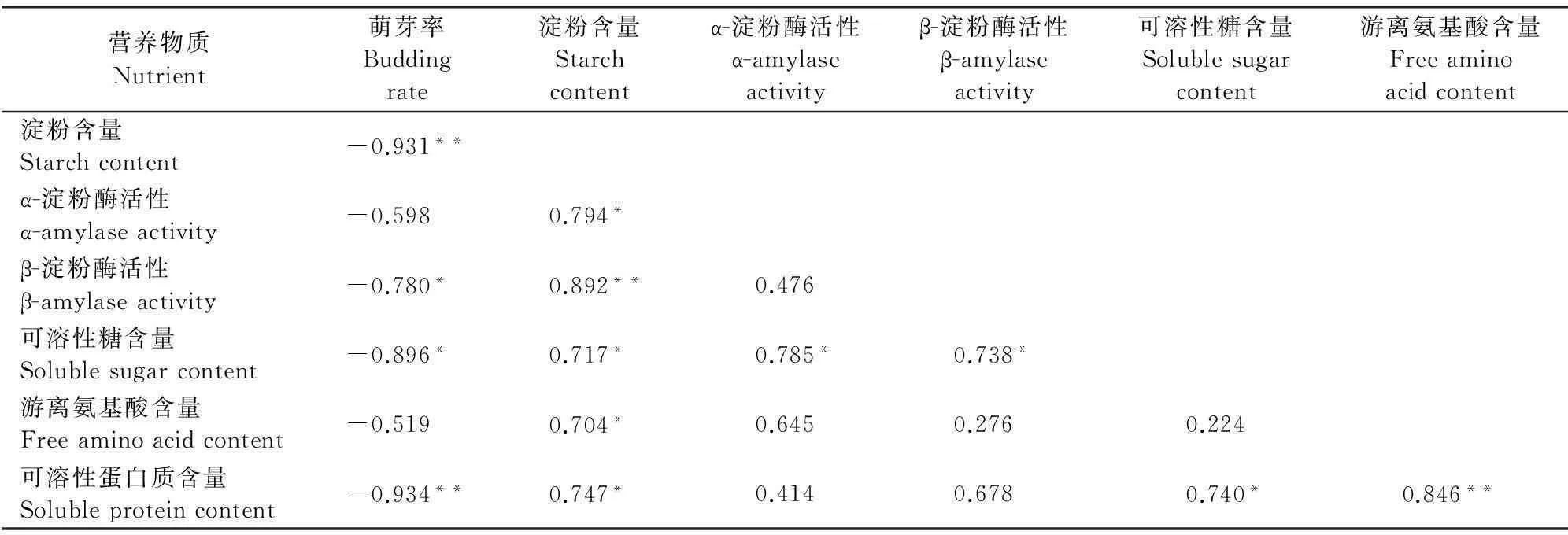
表1 ‘豐水’梨萌芽率與枝皮有機營養物質含量的相關性系數
注:*和**分別表示相關性在0.05和0.01水平達到顯著水平;下同。
Note:*and ** stand for the correlation is significant at 0.05 and 0.01 level,respectively;the same as below.

表2 ‘豐水’梨萌芽率與花芽有機營養物質含量的相關性系數
3討論
3.1二次開花過程中枝條營養運輸特征
南方砂梨二次開花的本質就是花芽分化完成后避開休眠或者在淺休眠階段解除休眠,誘導樹體連續生長發育的結果。黃新忠等[6]認為砂梨二次開花與早期落葉緊密相關。日本的林真二[12]指出,多雨的梨產區,葉片生產的同化養分較少,樹體內碳水化合物合成減少;此時新梢伸展迅速,對氮和碳水化合物的消耗極多,同時樹體吸收的大量氮素難以消化轉變為蛋白質,樹體內硝態氮或非蛋白質氮暫時增多,導致落葉從而引發二次開花的發生。趙志國等[7]認為砂梨二次花的發生與該現象發生前2~4個月內樹體無機營養元素水平無直接關系。關于樹體有機營養與砂梨二次開花的研究還未見報道。果樹內貯藏的碳水化合物是樹體抵御冬季嚴寒、冬季休眠消耗、萌芽和春季起始生長的物質基礎[13]。Amold[14]對蘋果根與梢的皮部全蛋白質含量作了測定,發現在低溫累積過程中,全蛋白質含量通常呈增加趨勢,直到葉芽低溫需求量被滿足,然后蛋白質開始降解。本試驗中去葉處理‘豐水’梨枝皮淀粉和可溶性糖含量均下降,而花芽糖含量卻略有升高;說明去葉處理后花芽萌動需要消耗大量的營養物質,去葉促進了枝皮淀粉向花芽可溶性糖的轉化,為代謝變化提供物質及能量。隨著去葉時間的增加,淀粉酶將不易溶解的大分子淀粉分解為易溶解吸收的小分子可溶性糖,所以,花芽中可溶性糖含量是一個逐漸增加的過程。本試驗還發現,枝皮蛋白質含量急劇降低,花芽中游離氨基酸的含量迅速增加也進一步證實了枝皮貯藏營養的供給關系。
本試驗還表明,‘豐水’梨萌芽率與枝皮淀粉和可溶性蛋白含量呈極顯著的負相關關系,與可溶性糖含量、β-淀粉酶活性呈顯著的負相關,與花芽中淀粉和可溶性蛋白含量亦呈顯著的負相關,而與中花芽可溶性糖呈正相關關系,表明了可溶性糖和二次花芽萌動的緊密關系;Mason[15]認為除了生長素及激素的平衡能夠調節頂花芽的萌動之外,糖的積累也能夠促進花芽的萌動并限制BRANCHED1(BRC1)基因的表達,這與本研究的結果一致。但是枝條和花芽的營養狀態是如何影響二次花發生的還有待于進一步研究。
3.2去葉對‘豐水’梨二次開花的影響和開花逆轉現象
去葉能促使成熟的花芽萌動、開花等[16]。去葉一般在花芽分化完成后自然休眠發生前進行,分為人工去葉和化學去葉,這樣可以使樹體避開休眠,進入下一生長周期。因此,花芽形成的時間和休眠發展的進程對于去葉而言至關重要[17],去葉時間過早影響花芽分化,過晚則休眠深度過深降低萌芽率,這就需要根據不同樹種的花芽分化進程和休眠發展進程的界定選擇各自合適的去葉時間。本研究選擇在梨果實采收后花芽分化完成以后全部去葉,滿足了成花的條件,試驗處理達到了預期的花芽萌動和開花的目的。關于去葉使樹體避開休眠的原理,研究認為去葉限制了芽內ABA的累積,并且使赤霉素含量升高[16]。
另外,本研究在去葉的過程中還發現,花芽在‘豐水’梨全部去葉后生長過程中可以直接抽出枝條。開花逆轉現象[18]的發生,說明分生組織的活動方式不僅可以從營養生長狀態轉向生殖發育狀態,而且可以從生殖發育狀態逆轉回營養生長狀態[19]。本研究同時驗證了在砂梨開花逆轉過程中伴隨著枝皮和花芽有機營養的轉換。
參考文獻:
[1]鐘必鳳,張全軍,李文貴,等.南方砂梨返花過程中內源激素含量變化及其與秋季返花的關系[J].西北植物學報,2014,34(12):2 453-2 458.
ZHONG B F,ZHANG Q J,LI W G,etal. Variation of endogenous hormone ofPyruspyrifoliaNakai in South China during returning bloom and its relationship with returning bloom[J].ActaBot.Boreal.-Occident.Sin,2014,34(12):2 453-2 458.
[2]LIU G Q,LI W S,ZHENG P H,etal.Transcriptomic analysis of ‘Suli’ pear (Pyruspyrifoliawhite pear group) buds during the dormancy by RNA-Seq [J].BMCGenomics,2012,13:700.
[3]DU PLOOY P,JACOBS G,COOK NC.Quantification of bearing habit on the basis of lateral bud growth of seven pear cultivars grown under conditions of inadequate winter chilling in South Africa [J].Sci.Hortic-Amsterdam,2002,95:185-192.
[4]OH S,KLINAC D.Relationship between incidence of floral bud death and temperature fluctuation during winter in Japanese Pear(Pyruspyrifoliacv.Hosui)under New Zealand climate conditions [J].Kor.J.HortSci.Technol.,2003,44:162-166.
[5]ROBSON R Y,AKEMI K H,etal. "Floral Primordia Necrosis" incidence in mixed buds of Japanese pear (Pyruspyrifolia(Burm.) Nakai var.culta) ‘Housui’ Grown under mild winter conditions and the possible relation with water dynamics [J].J.Japan.SocietyHort.Sci.,2010,79(3):246-257.
[6]黃新忠,陳義挺,雷龑,等.福建梨早期大量落葉誘因與防控策略[J].中國農學通報,2010,26(2):91-95.
HUANG X Z,CHEN Y T,LEI Y,etal.Causes and control strategies of a large number of early falling leaves of pear in Fujian [J].ChineseAgriculturalScienceBulletin,2010,26(2):91-95.
[7]趙志國,黃寧珍,唐鳳鸞,等.桂北南方優質早熟梨生理異常現象成因研究[J].廣西科學,2011,18(3):298-303.
ZHAO Z G,HUANG N Z,TANG F L,etal.Research on the cause of physiologic abnormity of south-early-ripening-pear in northern Guangxi[J].GuangxiSciences,2011,18(3):298-303.
[8]SIMINOVITCH D.Evidence from increase in ribonucleic acid and protein synthesis in autumn for increase in protoplasm during the frost-hardening of block locust bark cells[J].Can.J.Bot.,1963,41:1 301-1 308.
[9]YOUNG E,CYTOKININ,JOLUBJE.Carbohydrcte concentrations in xylen sap of apple during dormancy and bud bread [J].J.Am.Soc.HortSci,1989,114(2):297-300.
[10]張全軍,鐘必鳳,李文貴,等.南方砂梨返花返青發生特性及其與早期落葉的關系[J].西南農業學報,2013,26(3):1 160-1 163.
ZHANG Q J,ZHONG B F,LI W G,etal.Reflorescence and refoliation characteristics ofPyruspyrifoliaNakai in south China and its relationship with early defoliation[J].SouthwestChinaJ.Agric.Sci,2013,26(3):1 160-1 163.
[11]李合生,植物生理實驗指導[M].北京:高等教育出版社,2003
[12]林真二.梨[M]//吳耕民,譯.北京:中國農業出版社,1981:34.
[13]KAZUO IWASAKI,ROBERT J.WEAVER.Effects of chilling,calcium cyanamide,and bud scale removal on bud break,rooting,and inhibitor content of buds of‘Zinfandelgrape (VitisviniferaL.)[J].J.Am.Soc.Sci.,1977,102(5):584-587.
[14]AMOLD M A,YOUNG E.Gronth and protein content of apple in response to root and shoot temperature following chilling[J].Hortscience,1990,25(12):1 583-1 588.
[15]MASON M G,ROSS J J,BABST B A,etal.Sugar demand,not auxin,is the initial regulator of apical dominance [J].P.Natl.Acad.Sci.USA,2014,111:6 092-6 097.
[16]王海波,王孝梯,劉鳳之,等.落葉果樹無休眠栽培的原理與技術體系[J].果樹學報,2007,24(2):210-214.
WANG H B,WANG X T,LIU F Z,etal.Principles and technical systems of non-dormancy culture on deciduous fruit tree[J].JournalofFruitScience,2007,24(2):210-214.
[17]彭抒昂,羅充,章文才.鈣和鈣調素在梨花芽分化中的動態研究[J].武漢植物學研究,1999,17(2):178-180.
PENG S A,LUO C,ZHANG W C.Studies on the changes of calcium and Calmodulin during flower differentiation of pear[J].JournalofWuhanBotanicalResearch,1999,17(2):178-180.
[18]吳存祥,韓天富.植物開花逆轉研究進展[J].植物學通報,2002,19(5):523-529.
WU C X,HAN T F.Progress in study of flowering reversion in plants [J].ChineseBulletinofBotany,2002,19(5):523-529.
[19 ]BATTEY N H,LYNDON R F .Reversion of flowering[J].Bot.Rev.,1990,56:162-189.
(編輯:裴阿衛)
Variation of Organic Nutrition on Shoot and Flower Bud of ‘Hosui’Pear (PyruspyrifoliaNakai) during Reflorescence Period
ZHANG Quanjun1,ZHONG Bifeng1,LI Wengui1,DENG Jialin1,ZHANG Shaoling2
(1 Horticultural Research Institute of Sichuan Academy of Agricultural Sciences/Key Laboratory of Horticultural Crops,Biology and Germplasm Enhancement in Southwest Regions,Ministry of Agriculture,Chengdu 610066,China;2 Center of Engineering and Technology Research,Nanjing Agricultural University,Nanjing 210095,China)
Abstract:To elucidate the relationship between reflorescence and nutrition conditions,were investigated the organic nutrition changes in shoots and flower buds from the Pyrus pyrifolia cv.Hosui during reflorescence.The results showed that:(1)along with the bud break at 6 d defoliated by hand,the budding rate was keeping low rising,and flowers opened fully at 42 d.(2)The soluble sugar was increased firstly when the tree defoliated by hand.However,the soluble sugar content was decreased at the 12 d of rapid increasing stage for budding rate.The budding rate increasing stage was consistent with starch contents reducing periods in shoots.In addition,protein content in shoots was decreased to promote rapid increasing for the free amino acid content in the flower buds.(3)The related positive or negative correlation for budding rate and starch content,soluble protein content in flower bud and shoot reached at the 1% significant level,respectively.The positive correlation for budding rate and soluble sugar content in flower bud were 0.794,reached at the 5% significant level.It was indicated the nutrition conditions played important roles in the reflorescence occurrence for Pyrus pyrifolia cv.Hosui.
Key words:Pyrus pyrifolia Nakai;reflorescence;nutrition condition;soluble sugar;starch;soluble protein
中圖分類號:Q945.6
文獻標志碼:A
作者簡介:張全軍(1975-),男,博士,副研究員,從事果樹栽培生理與育種工作。E-mail:124317090@qq.com
基金項目:國家梨產業技術體系建設項目(NYCYTX29-37);四川省應用基礎研究項目(15JC0096)
收稿日期:2015-10-29;修改稿收到日期:2016-03-11
文章編號:1000-4025(2016)03-0493-06
doi:10.7606/j.issn.1000-4025.2016.03.0493
