水稻WRKY轉錄因子基因家族響應外源一氧化氮的表達譜分析
2016-05-09 09:41:35孟姣王海華向建華蔣丹彭喜旭3賀歡歡
中國水稻科學 2016年2期
關鍵詞:水稻
孟姣王海華,2,3,*向建華蔣丹彭喜旭,3賀歡歡
(1湖南科技大學生命科學學院,湖南湘潭411201;2重金屬污染土壤生態修復與安全利用湖南省高校重點實驗室,湖南湘潭411201;3園藝作物病蟲害防治湖南省重點實驗室,湖南湘潭411201;*通訊聯系人,E-mail:hhwang@hnust.edu.cn)
?
水稻WRKY轉錄因子基因家族響應外源一氧化氮的表達譜分析
孟姣1王海華1,2,3,*向建華1蔣丹1彭喜旭1,3賀歡歡1
(1湖南科技大學生命科學學院,湖南湘潭411201;2重金屬污染土壤生態修復與安全利用湖南省高校重點實驗室,湖南湘潭411201;3園藝作物病蟲害防治湖南省重點實驗室,湖南湘潭411201;*通訊聯系人,E-mail:hhwang@hnust.edu.cn)
孟姣,王海華,向建華,等.水稻WRKY轉錄因子基因家族響應外源一氧化氮的表達譜分析.中國水稻科學,2016,30(2):111-120.
摘 要:一氧化氮(nitric oxide,NO)信號分子與WRKY轉錄因子均參與植物抗逆、發育與代謝等許多生理過程。采用Agilent水稻全基因組cDNA芯片分析了NO處理后1、6和12h水稻幼苗WRKY轉錄因子基因的表達譜,鑒定出在1個時間點有兩倍或兩倍以上表達變化的WRKY基因32個,主要分布在WRKY的Ⅰ和Ⅱ組,其中75%的Ⅱa和45.6%的Ⅱd亞組成員為差異表達基因;鑒定出至少在2個時間點有兩倍或兩倍以上表達變化的WRKY基因15個,均為早期(1h)應答,且多數(64.2%)持續上調;基因功能預測分析表明,這些基因主要參與生物學過程中的細胞過程、代謝過程和刺激響應,以及分子功能中的轉錄調節活性和結合;代謝通路分析表明,WRKY24涉及植物與病原菌相互作用代謝通路。實時熒光定量PCR驗證結果與芯片雜交結果基本一致,印證了芯片雜交結果的有效性。上述發現提示,NO信號可能參與了WRKY轉錄因子介導的生物學調控功能,并為這些基因的進一步功能分析奠定基礎。
關鍵詞:水稻;WRKY轉錄因子;一氧化氮;基因表達譜
植物在正常生長條件下的發育與代謝以及在環境脅迫條件下的應答均涉及一系列相關基因在空間和時間上的差異表達與調控。轉錄是基因表達的第一步,體現了效率和靈活的原則,因此,轉錄水平的調控在植物基因表達調控中占據中心位置。轉錄因子與靶基因啟動子中的順式元件特異結合,從而激活或抑制基因的表達,在植物的發育、代謝以及環境適應等諸多生理過程中扮演重要的角色。全基因組基因鑒定表明,擬南芥和水稻基因組分別有超過2100和2300個轉錄因子基因,而且相當大比例的轉錄因子基因為植物所特有[1]。轉錄因子基因在全基因組尺度上的cDNA微陣列表達分析和突變體的遺傳分析都證明了它們在植物基因表達調控中的重要性。
WRKY蛋白在高等植物中構成轉錄因子超家族[2]。它們最典型的結構特征是含有1~2個保守的WRKY結構域:由大約60個氨基酸組成,其N端的七肽WRKYGQK十分保守,C端為鋅指結構。根據所含WRKY結構域的數目和鋅指類型,WRKY蛋白可分成三個組別。第Ⅰ組含兩個WRKY結構域,鋅指類型為C-X4-5-C-X22-23-H-X-H;第Ⅱ和Ⅲ組只含1個WRKY結構域,鋅指分別為C-X4-5-C-X22-23-H-X-H和C-X7-C-X23-H-X-C。WRKY蛋白C端的WRKY結構域具DNA結合活性,而N端的WRKY結構域可能介導蛋白質之間的相互作用,或具有輔助DNA結合的活性。WRKY蛋白絕大部分均與W盒[(T)(T)TGAC (C/T)]順式元件結合[2],從側面反映了WRKY域的保守性。WRKY保守結構域外的氨基酸順序分歧較大,這與不同的WRKY轉錄因子具有不同的生物學功能相符。
WRKY轉錄因子最顯著的功能是調節植物抗病反應[3],并在植物抗高溫/低溫、高鹽、干旱、滲透和營養饑餓等非生物逆境以及發育和代謝中具有重要的調節功能[4-5]。WRKY轉錄因子對植物生理過程的調節作用與水楊酸(SA)、茉莉酸(JA)和脫落酸(ABA)等激素介導的信號途徑密切相關。例如,擬南芥WRKY70(AtWRKY70)和水稻WRKY13(Os-WRKY13)位于SA和JA抗病信號途徑的交叉點,通過激活前者而抑制后者,從而調控植物抗病反應[6-7]。我們已證明,OsWRKY4作為JA信號途徑中的正調節子,介導水稻對紋枯病的抗性[8]。突變體的遺傳分析表明,AtWRKY40、AtWRKY18和 AtWRKY60的敲除突變體均在種子萌發過程中表現出ABA超敏表型,三者協同在ABA信號傳遞中起負調控作用[9]。
全基因組表達譜分析有助于初步推測基因家族成員的生物學功能。Ryu等[10]運用RT-PCR和Northern雜交研究了水稻WRKY基因家族在稻瘟病菌接種以及SA和JA處理下的表達譜,發現在所檢測的45個WRKY基因中有15個表達上調,Os-WRKY45和OsWRKY62的表達受SA誘導,Os-WRKY10、OsWRKY82和OsWRKY85的表達受JA誘導,而OsWRKY30和OsWRKY83既受SA又受JA誘導。Ramamoorthy等[11]運用RT-PCR在全基因組范圍研究了水稻WRKY基因在寒冷、干旱和鹽脅迫以及SA、JA、ABA等5種激素處理下的表達譜,結果表明,共有54個WRKY基因的表達發生了顯著差異,其中13個基因的表達僅受非生物脅迫的影響,13個基因的表達僅受激素的影響,而剩余的28個基因的表達同時受非生物脅迫和激素的調節,提示WRKY調節非生物脅迫反應和激素介導的信號傳遞之間存在交互作用。
一氧化氮(nitric oxide,NO)是一種具有高度反應活性、膜透性的氣態小分子。作為信號激素廣泛參與植物發育過程,包括促進種子萌發、氣孔關閉、不定根的發生和打破種子休眠等[12]。同時,NO具有緩解植物生物與非生物脅迫的作用,如病原、鹽、滲透、熱和金屬脅迫等[13-14]。NO如何影響植物WRKY的表達以及是否參與該家族對生理的調節作用鮮有報道,然而仍有線索表明水稻WRKY基因受NO的調控。cDNA微陣列雜交分析表明,At-WRKY15、AtWRKY25、AtWRKY40和At-WRKY53在NO處理下顯著上調表達[15]。At-WRKY27在擬南芥對細菌萎蔫病菌(Ralstonia solanacearum)抗性反應中發揮負調節作用。Atwrky27突變體對NO供體硝普鈉(含有活性亞硝基配體,溶液狀態下可釋放有活性的NO)的處理十分敏感。表達譜分析表明,WRKY27負調控氮代謝相關基因和NO合成相關基因的表達[16]。然而,從全基因組范圍探討NO對水稻WRKY基因表達的影響未見報道。
為此,我們采用水稻全基因組cDNA微陣列(Agilent,4×44k)對硝普鈉處理下水稻WRKY基因進行了轉錄組學研究,鑒定了受NO調節的WRKY基因(包括差異表達基因和響應基因),運用生物信息學方法對NO響應的WRKY基因進行了GO分析和代謝通路分析,部分WRKY的表達得到了qRT-PCR的驗證。
1 材料與方法
1.1 水稻材料培養與硝普鈉處理
將水稻日本晴(Oryza sativa L.japonica cv.Nipponbare)播種在添加木村B營養液的蛭石基質上,在人工氣候室中培養至3葉1心期,培養條件為光強100μmol/(m2·s),晝夜周期14h/10h,28℃/22℃,相對濕度約85%。用100μmol/L硝普鈉溶液(含0.1%吐溫80)對水稻進行葉面噴施;為了保證處理充分,另在培養液中添加相同濃度的硝普鈉(不含吐溫80),分別在處理0(T0,對照)、1(T1)、6 (T6)和12h(T12)后取葉片,用錫箔紙包裹置于液氮中,-80℃下保存。
1.2 葉綠素含量測定
葉綠素提取與含量測定采用分光光度法和Arnon公式計算[17]。
1.3 RNA提取和純化
總RNA抽提采用TRIZOL試劑,根據廠商提供的標準操作流程進行,經Agilent Bioanalyzer 2100電泳質檢合格后,用RNeasy mini kit和RNase-Free DNase Set純化。
1.4 芯片雜交和數據標準化
采用Agilent rice oligo microarray(4×44k)芯片。該芯片代表了42 489個水稻基因或轉錄本,所有注解均可在RAP-DB數據庫進行查詢。芯片雜交和雜交信號處理委托上海伯豪生物技術公司完成,實驗方法參照Agilent單色微陣列雜交操作手冊。每個時間點的樣品進行兩次獨立的微陣列雜交。從Feature Extraction Software獲得的微陣列雜交信號導入GeneSpring GX軟件(Agilent Technologies)。數據用P、A、M(分別表示正常、缺失和邊緣化的雜交信號)標記,隨后用Quantile算法進行標準化,并基于所有樣本的信號中值進行基線處理。
1.5 數據篩選、聚類和功能分類
將標準化的數據導入SAS系統,獲得數據的相關系數。標準化信號值經Log2處理,計算基因在處理后不同時間點(Tn)與T0間的差異表達倍數(FC):FC=MeanTn/MeanT0。將在某一時間點上具有兩倍或兩倍以上表達變化的基因定義為差異表達基因(DEGs);將在3個處理時間點上至少在2個時間點有兩倍或兩倍以上表達變化的基因定義為NO響應基因。
用維恩圖鑒定出NO響應基因后,用MeV 4.0軟件獲得各組重復樣本的聚類熱圖。根據RAP-DB數據庫提供的注釋文檔轉換格式后,利用agriGO對NO響應基因進行GO分析,用SAS的分子注釋功能得到通路分析圖,最后對NO響應的WRKY基因進行表達趨勢聚類分析。
1.6 qRT-PCR分析
隨機挑選8個WRKY基因,利用qRT-PCR對其表達水平進行驗證。用Primer Premier 5.0設計基因特異性引物(表1),擴增產物大小在150bp左右。水稻Actin 1(Os03g0718100)作為內參基因。
cDNA合成之前用DNaseⅠ消除基因組DNA污染。用FastQuant RT Kit(with gDNase)合成cDNA第一鏈。qRT-PCR在Bio-Rad CFX96PCR System上進行,反應體系包括10μmol/L的正反引物各0.6μL,2×SuperReal PreMix Plus 10μL,cDNA模板6μL,RNase-free ddH2O補至20μL。
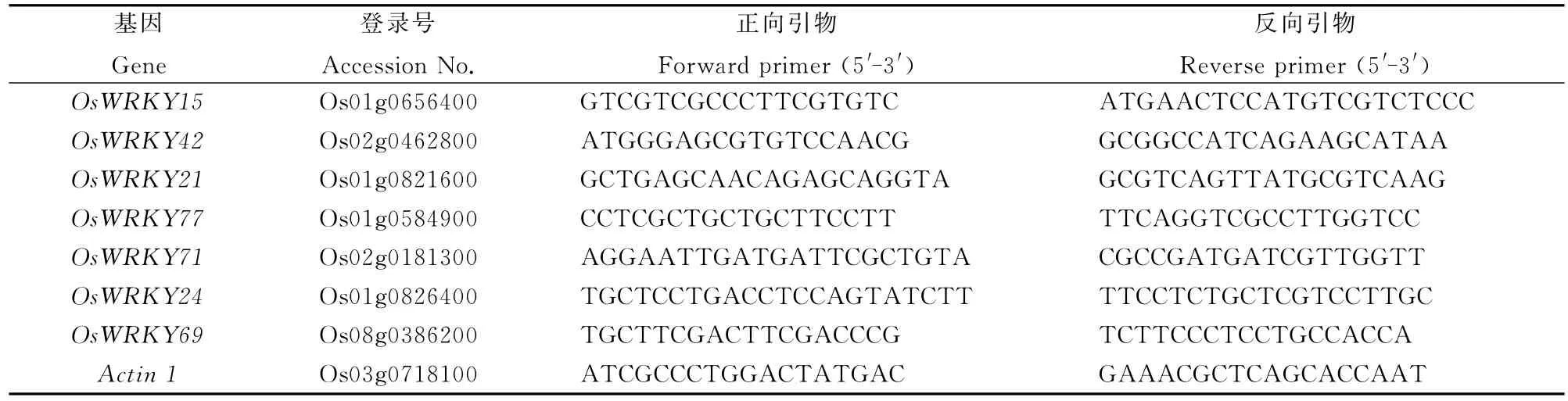
表1 候選基因的qRT-PCR引物Table 1.Primers of the candidate genes for qRT-PCR.
反應程序如下:95℃下15 min;95℃下10s,60℃下30s,40個循環。設陰性對照,三次技術重復和兩次生物學重復。結果釆用相對定量的方法(2-△△CT)計算基因的表達變化倍數。
2 結果與分析
2.1 微陣列數據相關性分析
將標準化的基因表達各組數據導入SAS系統,用Correlation Analysis分析微陣列雜交實驗的相關性,重復樣本數據間的相關性系數均大于0.98(結果未顯示),說明實驗具有較好的重復性,可用于后續分析。
2.2 NO處理下各時間點水稻葉片葉綠素含量測定
為了保證基因轉錄只發生中等程度的變化和獲得特異的基因表達譜,我們對NO供體硝普鈉的使用濃度進行了前期的摸索,發現100μmol/L硝普鈉處理未對水稻幼苗葉綠素含量(圖1)和形態產生明顯的影響。
2.3 NO處理下WRKY基因的差異表達分析
手工搜索發現芯片共有WRKY探針112個,代表71個WRKY基因,它們在各時間點的差異表達倍數見在線輔助性表S1(http://www.ricesci.cn/CN/article/showSupportInfo.do?id=2580)。國際上多個獨立的研究小組致力于水稻WRKY功能的研究,先后出現了幾種不同的WRKY命名系統,給水稻WRKY的研究與交流帶來了一些混亂。為此,國際水稻WRKY工作組提出了統一的WRKY命名規則[18]。我們對芯片中71個WRKY基因中不符合規則的編號進行修正。數據篩選結果表明,在3個NO處理時間點的某一時間點發生兩倍或兩倍以上表達變化的WRKY探針42個,基因32個(表2)。在處理1h時上調表達WRKY基因數為24個,下調9個;在6h時上調表達WRKY基因數為18個,下調表達15個;在12h時上調表達的WRKY基因數為26個,下調表達7個(圖2)。
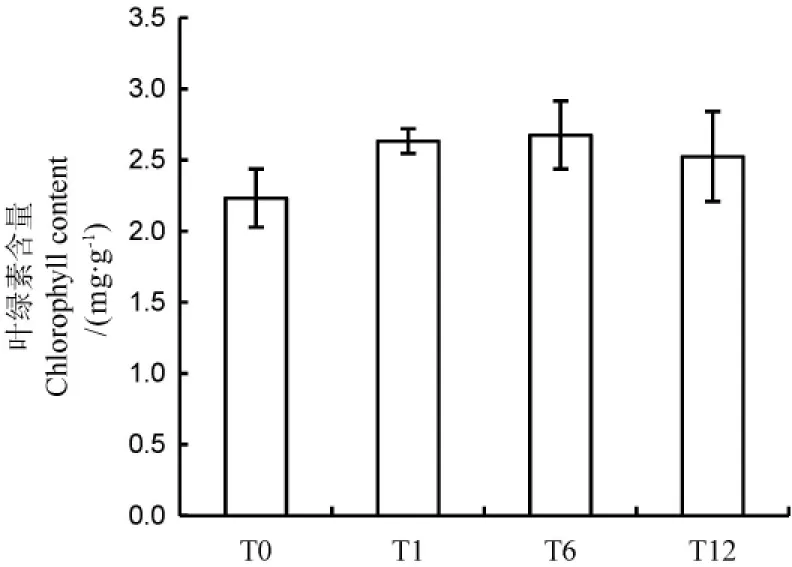
T0、T1、T6、T12分別表示NO處理0、1、6和12h。下同。NO treatments for 0,1,6and 12hare indicated as T0,T1,T6and T12,respectively.The same as below.圖1 NO處理不同時間后水稻葉片葉綠素含量Fig.1.Chlorophyll contents in rice leaves after NO treatment for different time.
水稻粳稻基因組中鑒定的WRKY基因家族成員超過100個,分成Ⅰ、Ⅱ和Ⅲ組,其中第Ⅱ組又細分為a~d四個亞組[19]。芯片雜交結果表明,差異表達的WRKY基因在Ⅰ和Ⅱ組中的比例接近,分別為35.3%和36.7%,而占水稻WRKY基因家族最多的Ⅲ組的比例較低(25.0%)。Ⅱa亞組包含4個基因,其中3個為差異表達基因;將近一半(5/11)的Ⅱd亞組為差異表達基因(表3)。
用Venny分析鑒定了在3個NO處理時間點上至少在2個時間點有兩倍及兩倍以上表達變化的WRKY基因,即NO響應的WRKY基因。結果表明,NO響應的WRKY探針20個,代表15個基因(圖3-A)。將基因的差異表達倍數取Log2后導入MEV軟件,得到聚類熱圖(圖3-B)。
2.4 NO響應的WRKY基因的GO分析和通路分析
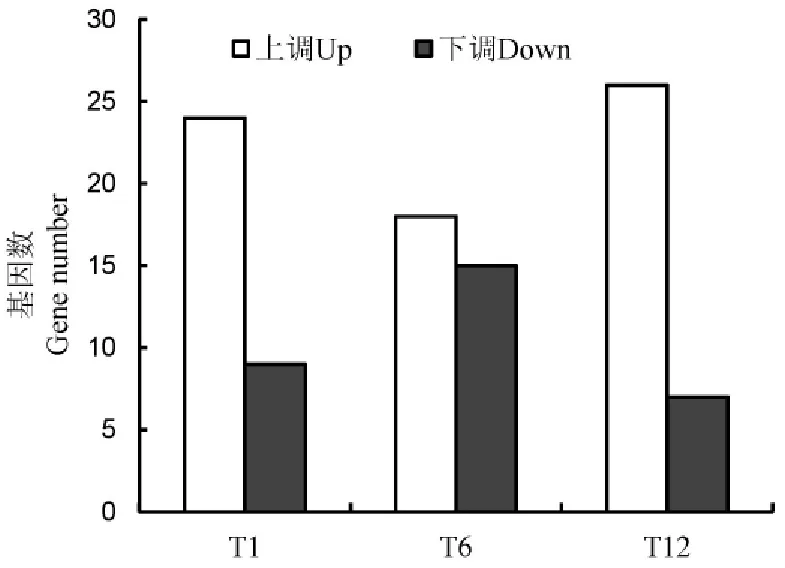
圖2 NO處理后不同時間后差異表達的水稻WRKY基因Fig.2.Differentially expressed rice WRKY genes after NO treatments for different time.
Gene Ontology是一種生物本體語言,提供了三層結構的系統定義方式,即分子功能(molecular function)、生物過程(biological process)和細胞組成(cellular component)三個部分。根據RAP-DB數據庫所提供的注釋文檔將基因名稱轉換格式后,借助在線工具agriGO(http://bioinfo.cau.edu.cn/agriGO/)進行GO富集分析,結果表明,注釋的15個NO響應的WRKY基因主要富集在生物過程中的細胞過程(cellular process,GO:0009987)、代謝過程(metabolic process,GO:0008152)和刺激應答(response to stimulus,GO:0050896),以及分子功能中的轉錄調節子活性(transcription regulator activity,GO:0030528)和結合(binding,GO:0005488)(圖4)。
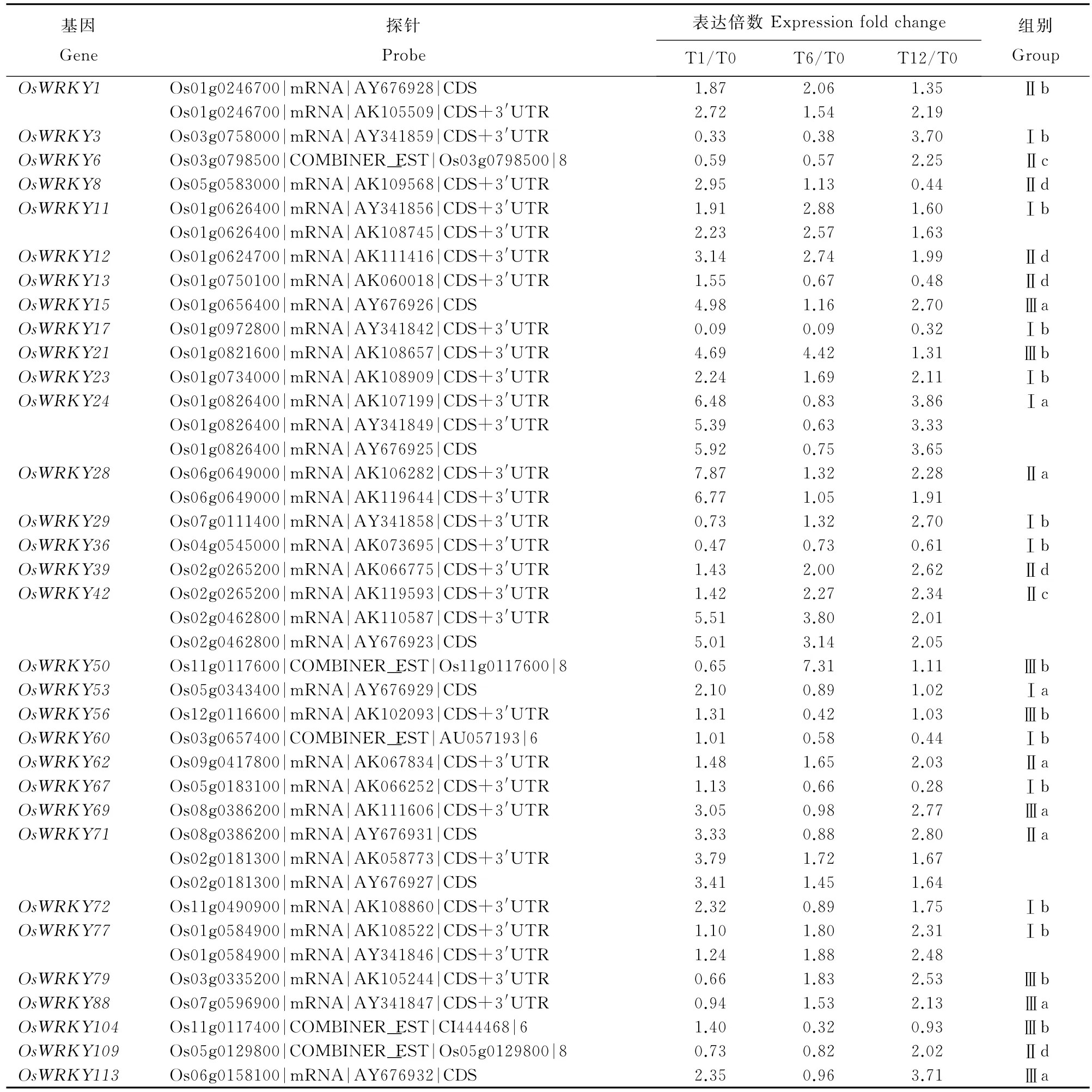
表2 NO處理下水稻WRKY差異表達基因的芯片分析Table 2.Microarray analysis of differentially expressed WRKYgenes in rice under NO treatment.
將NO響應的WRKY基因導入SAS系統,通過分子注釋發現OsWRKY24(Os01g0826400)參與植物與病原互作通路(詳見輔助性圖S1,http://www.ricesci.cn/CN/article/showSupportInfo.do?id=2580)。
2.5 NO響應的WRKY基因的表達聚類分析
基于NO處理過程中全基因組水平的基因表達變化的復雜性,對基因差異表達倍數取Log2后,對15個NO響應的WRKY基因的表達規律進行聚類。結果表明,這些基因分為5個具有各自表達規律的類別(圖5)。類別1表示持續上調,有9個基因(OsWRKY1、OsWRKY11、OsWRKY12、Os-WRKY15、OsWRKY21、OsWRKY23、OsWRKY28、OsWRKY39和OsWRKY42);類別2表示持續下調,有1個基因(OsWRKY17);類別3表示先上調后下調,有1個基因(OsWRKY8);類別4表示上調-下調-上調,有3個基因(OsWRKY113、OsWRKY69 和OsWRKY24);類別5表示先下調后上調,有1個基因(OsWRKY3)。

表3 NO處理下水稻WRKY差異表達基因在各組中的分布Table 3.Group distribution of differentially expressed WRKYgenes of rice responsive to nitric oxide.
2.6 基因表達水平的qRT-PCR驗證
為了驗證微陣列數據的可靠性,利用qRTPCR檢測了同—批材料中隨機挑選的6個WRKY基因的表達水平。結果表明,除個別WRKY基因的個別處理外(如OsWRKY24和OsWRKY69的T6),qRT-PCR所獲得的基因表達水平的變化與微陣列數據基本一致(圖6),反映了微陣列數據的可靠性。
3 討論
面對各種環境脅迫,植物進化出復雜而精巧的適應機制,涉及信號感知、信號轉導和相關基因表達的再編程。其中,較為普遍的策略是通過轉錄因子來調節下游靶蛋白的表達,進而在生長發育與逆境適應之間建立暫時的新的代謝平衡。因此,轉錄因子作為信號轉導途徑的終端事件——基因表達的調控者,在植物的適應機制中扮演非常重要的角色。在高等植物中,越來越多的證據表明,WRKY參與了各種脅迫反應、發育和代謝等生理過程[3-5],而WRKY對植物某些生理過程的調節作用往往與SA、JA、ABA和生長素介導的信號途徑相互聯系在一起。全基因組表達譜分析證實了上述觀點。例如,Dong等[20]用Northern雜交研究了擬南芥WRKY基因家族在植物抗病反應中的表達情況,發現7 2個WRKY基因中有4 3個受SA或病原細菌的誘導,6個受抑制。Kalde等[21]報道,第Ⅲ組的大多數成員既受SA又受病原誘導。同樣,在水稻中,Ryu等[10]和Ramamoorthy等[11]采用RT-PCR和Northern雜交對WRKY基因的表達譜進行了研究,發現了一些響應生物或非生物逆境、SA、JA和ABA等激素信號的WRKY基因。

圖4 NO響應的水稻WRKY基因的GO分析Fig.4.GO analysis on the NO-responsive rice WRKYgenes.

Log2FC-Log2處理后的基因表達變化倍數。Log2FC indicates Log2transformed expression fold change.圖5 NO響應的水稻WRKY基因的表達模式Fig.5.Expression patterns of the NO-responsive rice WRKYgenes.
作為一種高通量、靈敏的基因表達檢測手段,基于mRNA表達的基因芯片技術目前廣泛應用于植物在正常生長條件下的組織特異性、發育階段特異性以及在各種逆境下的基因表達譜研究;結合生物信息學方法,可以對差異表達基因進行GO、代謝通路、表達聚類、共表達(coexpression)以及啟動子順式元件分析,相對于RT-PCR、Northern雜交、cDNA-AFLP等傳統的基因表達分析手段具有明顯的優點。基因表達分析為揭示基因功能提供重要信息。基因在哪種條件下表達,預示該基因可能在該條件下發揮作用。NO作為植物體內重要的信使分子,被證明廣泛參與植物的生長發育、代謝以及環境脅迫適應等生理過程[12-14]。本研究用Agilent水稻芯片從全基因組角度系統分析了水稻WRKY基因家族在NO處理下的表達譜。結果表明,NO處理下水稻WRKY基因家族中45.1%(32/71)為差異表達基因,21.1%(15/71)為NO響應基因。注釋的15個NO響應的WRKY基因主要富集在生物過程中的細胞過程、代謝過程和刺激應答,以及分子功能中的轉錄調節子活性和結合,這與WRKY蛋白作為轉錄因子相互印證。基于cDNA微陣列分析,Palmieri等[15]發現,至少有4個擬南芥WRKY基因的表達受NO調控,其中3個直系同源(ortholog)基因的表達在本研究中亦受NO調控(At-WRKY15與OsWRKY119;AtWRKY40與Os-WRKY28、OsWRKY71;AtWRKY53與OsWRKY113),證明了本研究的可靠性。啟動子分析表明,在NO響應基因的啟動子區域中有8個類別的轉錄因子結合位點出現的頻率比芯片中所含的28 447個對照基因至少高出15%,其中包含WRKY轉錄因子的結合位點[15]。這些發現提示NO信號途徑可能參與了WRKY調節的生理過程。NO處理下水稻WRKY的差異表達基因比例比擬南芥高,可能與植物種類和NO供體及其處理方式不同有關。在Palmieri等[15]對擬南芥響應NO的基因表達譜研究中,使用的是氣態NO,而本實驗采用是硝普鈉,且同時對水稻幼苗進行了葉面噴施和灌根處理。

Log2FC-Log2處理后的基因表達變化倍數。Log2FC indicates Log2transformed expression fold change.圖6 NO響應的水稻WRKY基因表達水平的qRT-PCR驗證Fig.6.Expression verification of NO-responsive rice WRKYgenes by qRT-PCR.
我們的表達譜分析結果中也存在著在進化樹中兩兩相聚的成員(旁系同源,paralog)基因都趨向響應NO信號的現象(OsWRKY56與OsWRKY104; OsWRKY28與OsWRKY71;OsWRKY11與Os-WRKY8;OsWRKY23與OsWRKY72),這就暗示它們在功能上也可能具有相似性,為進一步探究水稻WRKY基因家族成員的功能提供了便利。有趣的是,水稻WRKYⅡa亞組包含4個成員,其中3個(OsWRKY28、OsWRKY62和OsWRKY71)屬于差異表達基因。WRKYⅡa亞組的4個基因均受SA、JA和稻瘟病菌侵染誘導,OsWRKY28和Os-WRKY71屬于稻瘟病菌侵染早期誘導型,Os-WRKY62和OsWRKY76屬于晚期誘導型[22]。然而,它們在水稻抗病反應中的作用完全不同。Os-WRKY28、WRKY62和WRKY76是水稻對稻瘟病或白葉枯病的負調因子;而OsWRKY71是水稻對白葉枯病的正調因子[22-25]。WRKYⅡd與鈣調蛋白(CaM)相互作用[26],NO與Ca2+信號系統交聯,調控植物細胞諸多生理過程[27]。將近一半(45.6%,5/11)的水稻WRKYⅡd亞組基因為NO差異表達基因。因此,今后在進一步研究水稻WRKYⅡa和Ⅱd亞組成員的生物學功能時,要關注NO在其中發揮的作用,及其與SA、JA和Ca等其他信號之間的交互影響。
NO為植物應激反應的早期信號分子[12]。水稻WRKY基因家族中的15個NO響應基因的表達聚為5個不同的類別,這些聚類均在NO處理早期(1h)表現出顯著的表達差異,其中類別1、類別3和類別4的OsWRKY1、OsWRKY8、OsWRKY11、OsWRKY12、OsWRKY15、OsWRKY21、Os-WRKY23、OsWRKY24、OsWRKY28、OsWRKY39、OsWRKY42、OsWRKY69和OsWRKY113表現為早期上調;類別2的OsWRKY17表現為早期下調。WRKY基因被NO早期誘導或抑制符合WRKY作為轉錄因子的分子特征。在這些NO響應的WRKY基因中,部分基因的生物學功能得以不同程度的解析,它們主要參與抗病或對非生物逆境的響應。例如,OsWRKY12為Liu等[28]報道的Os-WRKY03,作為轉錄激活因子,調節水稻對白葉枯病的抗性。OsWRKY23是植物抗病和衰老過程的調節子,超表達該基因的擬南芥植株增強了對丁香假單胞桿菌(Pseudomonas syringae)的抗性,同時加速了黑暗誘導葉片衰老進程[29]。OsWRKY8參與抵抗滲透脅迫[30]。OsWRKY42能通過抑制JA信號相關基因負調控水稻對稻瘟病菌的響應,與OsWRKY45-2、OsWRKY13組成一個順次轉錄調控級聯,WRKY45-2轉錄激活WRKY13,WRKY13能直接抑制WRKY42[31]。最近,我們的研究表明,OsWRKY1和OsWRKY17的同源基因OsWRKY4正調控水稻對紋枯病的抗性[8]。水稻糊粉層細胞中的OsWRKY24的表達受脫落酸(ABA)誘導,但在赤霉素(GA)處理下穩定表達;瞬時表達分析表明OsWRKY24能有效抑制ABA和GA信號途徑,提示它作為兩條信號通路的共同抑制子,參與調控種子萌發及其萌發后的生長過程[32]。然而,本研究對NO響應的水稻WRKY基因的代謝通路分析表明,OsWRKY24參與植物與病原互作通路,提示它可能在抗病防御中起作用。另外,cDNA代表性差異分析表明,水稻根部組織中OsWRKY17受缺鐵脅迫誘導[33]。OsWRKY21受模擬酸雨(pH5.6)誘導,基因芯片雜交結合qRT-PCR驗證的結果表明,超表達水稻植株中1個過氧化物酶基因、2個谷胱甘肽轉移酶基因和4個細胞色素P450基因表達上調,提示OsWRKY21可能通過調節細胞內氧化還原環境以及降解有害物質來增強水稻對酸雨的抗性[34]。雖然上述研究未曾探究NO信號在這些WRKY基因功能中的作用,考慮到NO在植物抗逆和發育中的廣泛功能以及NO與SA、活性氧、ABA等其他信號分子之間的交互作用,結合本研究的結果,可以推測NO信號可能參與上述WRKY基因在植物抗病、抗非生物脅迫、衰老和代謝中的調節效應;同時提示,要進一步深掘這些基因的功能,必須考慮NO信號的潛在作用。據我們所知,仍未有從全基因組角度對水稻WRKY基因家族在NO處理下的表達譜分析。有關NO響應的水稻WRKY的功能有待下一步研究,它們的共表達基因及其啟動子區域順式元件的富集分析也值得期待。
在線輔助性信息:在線輔助性表S1和圖S1見中國水稻科學網站(http://www.ricesci.cn/CN/artide/showSupportInfo.do?id=2580)。
參考文獻:
[1] Ria?o-Pachón D M,Ruzicic S,Dreyer I,et al.PlnTFDB:An integrative plant transcription factor database.BMC Bioinform,2007,8:42.
[2] Eulgem T,Rushton P J,Robatzek S,et al.The WRKY superfamily of plant transcription factors.Trends Plant Sci,2000,5:199-206.
[3] Pandey S P,Somssich I E.The role of WRKY transcription factors in plant immunity.Plant Physiol,2009,150:1648-1655.
[4] Agarwal P,Reddy M P,Chikara J.WRKY:Its structure,evolutionary relationship,DNA-binding selectivity,role in stress tolerance and development of plants.Mol Biol Rep,2011,38:3883-3896.
[5] Chen L,Song Y,Li S,et al.The role of WRKY transcription factors in plant abiotic stresses.Biochim Biophys Acta,2012,1819:120-128.
[6] Li J,Brader G,Palva E T.The WRKY70transcription factor:A node of convergence for jasmonate-mediated and salicylatemediated signals in plant defense.Plant Cell,2004,16:319-331.
[7] Qiu D,Xiao J,Ding X,et al.OsWRKY13 mediates rice disease resistance by regulating defense-related genes in salicylate-and jasmonate-dependent signaling.Mol Plant Microbe Interact,2007,20:492-499.
[8] Wang H,Meng J,Peng X,et al.Rice WRKY4acts as a transcriptional activator mediating defense responses toward Rhizoctonia solani,the causing agent of rice sheath blight.Plant Mol Biol,2005,89(1/2):157-171.
[9] Shang Y,Yan L,Liu Z Q,et al.The Mg-chelatase H subunit of Arabidopsis antagonizes a group of WRKY transcription repressors to relieve ABA-responsive genes of inhibition.Plant Cell,2010,22,1909-1935.
[10]Ryu H S,Han M,Lee S K,et al.A comprehensive expression analysis of the WRKYgene superfamily in rice plants during defense response.Plant Cell Rep,2006,25:836-847.
[11]Ramamoorthy R,Jiang S Y,Kumar N,et al.A comprehensive transcriptional profiling of the WRKYgene family in rice under various abiotic and phytohormone treatments.Plant Cell Physiol,2008,49:865-879.
[12]Lamotte O,Courtois C,Pugin L B A,et al.Nitric oxide in plants:The biosynthesis and cell signalling properties of a fascinating molecule.Planta,2005,221:1-4.
[13]Gould K,Lamotte O,Klinguer A,et al.Nitric oxide production in tobacco leaf cells:A generalized stress response?Plant Cell Environ,2003,26:1851-1862.
[14]Delledonne M.NO news is good news for plants.Curr Opin Plant Biol,2005,8:390-396.
[15]Palmieri M C,Sell S,Huang X,et al.Nitric oxide-responsive genes and promoters in Arabidopsis thaliana:A bioinformatics approach.J Exp Bot,2008,59:177-186.
[16]Mukhtar M S,Deslandes L,Auriac M-C,et al.The Arabidopsis transcription factor WRKY27influences wilt disease symptom development caused by Ralstonia solanacearum.Plant J,2008,56:935-947.
[17]Arnon D I.Copper enzymes in isolated chloroplasts:Polyphenoloxidase in Beta vulgaris.Plant Physiol,1949,24:1-15.
[18]Rice WRKY Working Group.Nomenclature report on rice WRKY′s-Conflict regarding gene names and its solution.Rice,2012,5:3.
[19]Wu K L,Guo Z J,Wang H H,et al.The WRKY family of transcription factors in rice and Arabidopsis and their origins.DNA Res,2005,12:9-26.
[20]Dong J,Chen C,Chen Z.Expression profiles of the Arabidopsis WRKY gene superfamily during plant defense response.Plant Mol Biol,2003,51:21-37.
[21]Kalde M,Barth M,Somssich I E,et al.Members of the Arabidopsis WRKY groupⅢtranscription factors are part of different plant defense signaling pathways.Mol Plant Microbe Interact,2003,16:295-305.
[22]Chujo T,Miyamoto K,Shimogawa T,et al.OsWRKY28,a PAMP-responsive transrepressor,negatively regulates innate immune responses in rice against rice blast fungus.Plant Mol Biol,2013,82:23-37.
[23]Peng Y,Bartley L E,Chen X,et al.OsWRKY62is a negative regulator of basal and Xa21-mediated defense against Xanthomonas oryzae pv.oryzae in rice.Mol Plant,2008,1:446-458.
[24]Yokotani N,Sato Y,Tanabe S,et al.WRKY76is a rice transcriptional repressor playing opposite roles in blast disease resistance and cold stress tolerance.J Exp Bot,2013,64(16):5085-5097.
[25]Liu X,Bai X,Wang X,et al.OsWRKY71,a rice transcription factor,is involved in rice defense response.J Plant Physiol,2007,164:969-979.
[26]Park C Y,Lee J H,Yoo J H,et al.WRKY group IId transcription factors interact with calmodulin.FEBS Lett,2005,579:1545-1550.
[27]Durner J,Wendehenne D,Klessig D F.Defense gene induction in tobacco by nitric oxide,cyclic GMP,and cyclic ADP-ribose.Proc Natl Acad Sci USA,1998,95:10328-10333.
[28]Liu X Q,Bai X Q,Qian Q,et al.OsWRKY03,a rice transcriptional activator that functions in defense signaling pathway upstream of OsNPR1.Cell Res,2005,15(8):593-603.
[29]Jing S,Zhou X,Song Y,et al.Heterologous expression of OsWRKY23 gene enhances pathogen defense and dark-induced leaf senescence in Arabidopsis.Plant Growth Regul,2009,58:181-190.
[30]Song Y,Jing S,Yu D.Overexpression of the stress-induced OsWRKY08improves osmotic stress tolerance in Arabidopsis.Chin Sci Bull,2009,54:4671-4678.
[31]Cheng H,Liu H,Deng Y,et al.The WRKY45-2 WRKY13 WRKY42transcriptional regulatory cascade is required for rice resistance to fungal pathogen.Plant Physiol,2015,167(3):1087-1099.
[32]Zhang Z L,Shin M,Zou X,et al.A negative regulator encoded by a rice WRKYgene represses both abscisic acid and gibberellins signaling in aleurone cells.Plant Mol Biol,2009,70:139-151.
[33]Sperotto R A,Boff T,Duarte G L,et al.Increased senescence-associated gene expression and lipid peroxidation induced by iron deficiency in rice roots.Plant Cell Rep,2008,27:183-195.
[34]黃盼盼.18個水稻OsWRKY基因的克隆和OsWRKY21功能分析.廣州:中山大學,2010.Huang P P.Cloning 18 OsWRKYgenes and functional analysis of OsWRKY21.GuangZhou:Sun Yat-sen University,2010.(in Chinese with English abstract)
Expression Profiles of Rice WRKY Transcription Factor Gene Family Responsive to Exogenous Nitric Oxide Application
MENGJiao1,WANG Hai-hua1,2,3,*,XIANGJian-hua1,JIANGDan1,PENGXi-xu1,3,HE Huan-huan1
(1School of Life Science,Hunan University of Science and Technology,Xiangtan 411201,China;2 Key Laboratory of Ecological Remediation and Safe Utilization of Heavy Metal-polluted Soils,College of Hunan Province,Xiangtan 411201,China;3 Key Laboratory of Integrated Management of the Pests and Diseases on Horticultural Crops in Hunan Province,Xiangtan 411201,China;*Corresponding author,E-mail:hhwang@hnust.edu.cn)
MENG Jiao,WANG Haihua,XIANG Jianhua,et al.Expression profiles of rice WRKY transcription factor gene family responsive to exogenous nitric oxide application.Chin J Rice Sci,2016,30(2):111-120.
Abstract:Signal molecule nitric oxide(NO)and WRKY transcription factors are involved in many physiological processes,such as defense against stresses,development and metabolism in plants.In the present study,we investigated the expression patterns of WRKY transcription factor gene family in rice seedlings at 1,6and 12after exogenous NO treatments using Agilent rice cDNA oligo microarray.Totally,32differently expressed WRKYgenes,whose expression levels increased or decreased by at least two-fold at one time point compared with the control(T0),were identified.These WRKYgenes were mainly distributed over GroupsⅠandⅡ,among which 75%ofⅡa and 45.6%ofⅡd subgroup members were differently expressed upon the NO treatments.Moreover,15identified NO-responsive WRKYgenes,whose expression level changed by more than two-fold at least two time points among the three time points compared with the control,exhibited an earlier(1h)response to the NO treatments,and most of them(64.2%)were continually up-regulated.Prediction of gene function revealed that the NO-responsive WRKY genes were mainly involved in cellular process,metabolic process and response to stimulus of biological process,and transcription regulator activity and binding of molecular function.The analysis of metabolic pathways showed that WRKY24was involved in plant-pathogen interaction pathway.The results of microarray hybridization were largely consistent with those of quantitative real-time PCR,verifying the validity of microarray hybridization.These findings suggest that NO signaling might be involved in the regulatory functions of WRKY transcription factors,and provide a basis for further functional research for these differentially expressed WRKYgenes.
Key words:rice(Oryza sativa);WRKY transcription factor;nitric oxide;gene expression profile
中圖分類號:Q755;S511.01
文獻標識碼:A
文章編號:1001-7216(2016)02-0111-10
基金項目:國家自然科學基金資助項目(31171803,31301617);湖南省教育廳一般資助項目(14C0453);湖南科技大學創新基金資助項目(S140031)。
收稿日期:2015-09-16;修改稿收到日期:2015-11-30。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
