苜蓿屬和草木樨屬種質(zhì)資源親緣關(guān)系的過氧化物酶分析
2016-05-12 09:27:02陳立強邊艷霞馬春暉徐利明趙浩波楊浩宏
新疆農(nóng)業(yè)科學 2016年3期
陳立強,邊艷霞,馬春暉,徐利明,趙浩波,楊浩宏
(1.塔里木大學動物科學學院/塔里木畜牧科技兵團重點實驗室,新疆阿拉爾 843300;2.塔里木大學后勤管理處,
新疆阿拉爾 843300;3.石河子大學動物科技學院,新疆石河子 832000)
?
苜蓿屬和草木樨屬種質(zhì)資源親緣關(guān)系的過氧化物酶分析
陳立強1,邊艷霞2,馬春暉3,徐利明1,趙浩波1,楊浩宏1
(1.塔里木大學動物科學學院/塔里木畜牧科技兵團重點實驗室,新疆阿拉爾843300;2.塔里木大學后勤管理處,
新疆阿拉爾843300;3.石河子大學動物科技學院,新疆石河子832000)
摘要:【目的】分析苜蓿屬和草木樨屬種質(zhì)資源的親緣關(guān)系,提高兩屬植物遠緣雜交的可預見性。【方法】以苜蓿屬(Medicago)3個種和草木樨屬(Melilotus)2個種共20份種質(zhì)資源為材料,用過氧化物酶電泳技術(shù)對其親緣關(guān)系進行分析。【結(jié)果】苜蓿屬材料間、草木樨屬材料間、苜蓿屬和草木樨屬材料間的酶帶相似率分別介于30.00%~83.33%、33.33%~100.00%和0%~44.44%,相似系數(shù)分別介于0.417~0.917、0.667~1.000和0.083~0.667。供試材料在相似系數(shù)0.402處可聚為2類,草木樨屬材料為一類,苜蓿屬材料為一類。與草木樨屬相比,苜蓿屬材料在主成分分析圖中的位置相對分散,遺傳變異水平較高。【結(jié)論】部分遠緣種質(zhì)資源的酶帶相似程度較高,親緣關(guān)系較近,雜交成功的可能性較大。
關(guān)鍵詞:苜蓿屬;草木樨屬;過氧化物酶;親緣關(guān)系;遠緣雜交
0 引 言
【研究意義】苜蓿屬(Medicago)和草木樨屬(Melilotus)隸屬于豆科(Leguminosae)、蝶形花亞科(Papilionoideae)。屬內(nèi)植物多為優(yōu)良牧草,長期以來,在我國畜牧業(yè)發(fā)展、生態(tài)治理、種植業(yè)結(jié)構(gòu)調(diào)整及土壤肥力改良中發(fā)揮著重要作用。然而目前苜蓿屬和草木樨屬植物育種工作均相對滯后,現(xiàn)有品種遠不能滿足生產(chǎn)需求[1-2]。隨著苜蓿品種資源的日益枯竭,要想在育種領(lǐng)域取得突破,就必須通過遠緣雜交超越種間界限,創(chuàng)造新物種。育種實踐表明,遠緣雜交具有不可交配性,通常親本間親緣關(guān)系越近,雜交成功的可能性越大[3]。以此為依據(jù),研究苜蓿屬和草木樨屬種間和屬間種質(zhì)資源的親緣關(guān)系,對提高遠緣雜交可預見性、創(chuàng)造新物種具有重要意義。【前人研究進展】目前,已有研究對苜蓿及其近緣植物的親緣關(guān)系、草木樨資源的親緣關(guān)系進行了研究。劉磊等[4]用ISSR標記研究了紫花苜蓿(M.sativa)、黃花苜蓿(M.falcata)和胡盧巴屬(Trigonella)種質(zhì)資源的親緣關(guān)系,結(jié)果表明,供試種質(zhì)資源的相似系數(shù)介于0.850~0.940,胡盧巴(T.foenum-graecum)與黃花苜蓿的親緣關(guān)系較近。張宇等[5]用SRAP標記對扁蓿豆(M.ruthenica)-紫花苜蓿雜交種、紫花苜蓿和黃花苜蓿種質(zhì)資源的遺傳多樣性進行了研究,結(jié)果表明,供試苜蓿材料具有較高的異質(zhì)性。李飛飛等[6]用SSR和ISSR標記對黃花苜蓿、多變苜蓿(M.varia)及紫花苜蓿居群的遺傳多樣性及親緣關(guān)系進行了研究,結(jié)果表明,居群間的遺傳距離介于0.050~0.230,平均為0.120,多變苜蓿居群VG和黃花苜蓿居群FH表現(xiàn)出了較豐富的遺傳多樣性。Bagavathiannan等[7]用SSR標記對12個野生紫花苜蓿群體、10個紫花苜蓿栽培品種和1份黃花苜蓿種質(zhì)資源的遺傳多樣性進行了研究,結(jié)果表明,野生群體的遺傳變異水平較高。狄紅艷等[8]用ITS序列和trnL-trnF序列對18個白花草木樨(M.alba)和黃花草木樨(M.officinalis)種群的遺傳多樣性進行了研究,結(jié)果表明,白花草木樨和黃花草木樨種群間遺傳距離較小,黃花草木樨種群的遺傳多樣性水平較高。【本研究切入點】育種實踐表明,苜蓿屬種間雜交能取得成功[9],且苜蓿和草木樨的種子和實生苗均非常相似[10],并可通過體細胞融合產(chǎn)生雜種愈傷組織[11]。據(jù)此推斷,若親本選配恰當,苜蓿屬和草木樨屬種間甚至屬間雜交前景廣闊。遠緣雜交親本選配,親緣關(guān)系的遠近是關(guān)鍵,然而目前還沒有紫花苜蓿、黃花苜蓿、雜花苜蓿、白花草木樨、黃花草木樨間親緣關(guān)系的研究報道。研究種質(zhì)資源的親緣關(guān)系,提高遠緣雜交的可預見性。【擬解決的關(guān)鍵問題】研究選用過氧化物酶電泳技術(shù),對新疆野生紫花苜蓿、野生黃花苜蓿、野生雜花苜蓿、野生白花草木樨、野生黃花草木樨和紫花苜蓿栽培品種間的親緣關(guān)系進行分析,為進一步開展苜蓿屬和草木樨屬資源遺傳改良、雜交親本選配及野生種質(zhì)資源的開發(fā)利用提供依據(jù)。
1 材料與方法
1.1 材 料
野生種質(zhì)資源由塔里木大學草業(yè)科學學科組提供,均采自新疆,包括3份野生紫花苜蓿、2份黃花苜蓿、1份雜花苜蓿、5份白花草木樨和5份黃花草木樨。三得利、阿爾岡金和苜蓿王為新疆引進栽培品種,新疆大葉為新疆地方品種,種子由甘肅農(nóng)業(yè)大學草業(yè)學院提供。列出試驗材料的名稱、種類、類型、花色、原產(chǎn)地和其他特性。表1
表1供試材料的名稱、種類、類型、花色、原產(chǎn)地和性狀
Table 1List of all the accessions used in this study along with the name, specie, type, flower color, origin and characters description

序號No.材料Accession種類Specie類型Type花色Flowercolor原產(chǎn)地Origin性狀Charactersdescription1紫花苜蓿ⅠM.sativa野生材料紫色新疆拜城植株高大2紫花苜蓿ⅡM.sativa野生材料紫色新疆阿拉爾半匍匐3黃花苜蓿ⅠM.falcata野生材料黃色新疆巴里坤抗寒4雜花苜蓿ⅠM.varia野生材料雜花新疆巴里坤抗寒5紫花苜蓿ⅢM.sativa野生材料紫色新疆阿拉爾抗旱6三得利M.sativa栽培品種紫色法國抗倒伏7新疆大葉M.sativa栽培品種紫色新疆和田葉片特大8阿爾岡金M.sativa栽培品種紫色加拿大抗寒、抗旱9苜蓿王M.sativa栽培品種紫色美國抗逆性強10黃花苜蓿ⅡM.falcata野生材料黃色新疆吉木乃縣抗旱11白花草木樨ⅠM.alba野生材料白色新疆巴里坤抗寒12黃花草木樨ⅠM.officinalis野生材料黃色新疆巴里坤抗寒13白花草木樨ⅡM.alba野生材料白色新疆阿拉爾植株矮小14黃花草木樨ⅡM.officinalis野生材料黃色新疆阿拉爾不分枝15白花草木樨ⅢM.alba野生材料白色新疆阿拉爾植株高大16黃花草木樨ⅢM.officinalis野生材料黃色新疆拜城植株高大17白花草木樨ⅣM.alba野生材料白色新疆拜城植株高大18白花草木樨ⅤM.alba野生材料白色新疆阿拉爾半匍匐19黃花草木樨ⅣM.officinalis野生材料黃色新疆吉木乃縣抗旱20黃花草木樨ⅤM.officinalis野生材料黃色新疆阿拉爾有分枝
1.2 方 法
1.2.1 酶液提取
過氧化物酶從幼苗中提取,每份材料隨機選取100粒種子,在培養(yǎng)箱中發(fā)芽,至第8 d,提取酶液。稱取幼苗,置于研缽中,按幼苗重(g)∶緩沖液體積(mL)=1∶4的比例,加入0.1 mol/L的Tric-HCl緩沖液(pH=8.0)[12],研磨至勻漿,8 000 r/min離心10 min,取上清液分裝于1.5 mL離心管中,置于-80℃冰箱保存?zhèn)溆谩?/p>
1.2.2 電泳及染色
試驗選用聚丙烯酰胺垂直板凝膠電泳技術(shù)對酶液進行分離,膠板厚1.5 mm。分離膠濃度為7%,濃縮膠濃度為4%。選用高離子強度的電極緩沖液(pH=8.3)。以6×DNA loading buffer(溴酚藍,二甲苯青)為指示劑,上樣量20 μL,恒壓電泳,初始電壓100 V,待指示劑凝成藍線后,電壓升至200 V,指示劑移至玻璃板末端時終止電泳,采用醋酸聯(lián)苯胺法[13]進行染色。
1.3 數(shù)據(jù)統(tǒng)計
用遷移率定義不同酶帶,遷移率=酶帶中部的遷移距離/指示劑的遷移距離。以酶帶在相對遷移位置的有無,賦值為“1”或“0”,多態(tài)位點百分率=(多態(tài)性酶帶數(shù)/總酶帶數(shù))×100%,酶帶相似率=(共有酶帶數(shù)/總酶帶數(shù))×100%。用NTSYSpc 2.1軟件計算相似系數(shù)(genetic similarity,GS),進行聚類分析和主成分分析,構(gòu)建聚類圖和主成分二維圖。
2 結(jié)果與分析
2.1 酶帶的分布特征及相似程度
過氧化物酶在供試材料中共檢測到了12條酶帶,Rf依次為0.015、0.080、0.125、0.165、0.215、0.250、0.285、0.320、0.345、0.380、0.443和0.500,多態(tài)位點百分率為100.00%。紫花苜蓿酶譜共表現(xiàn)出了11條酶帶,Rf為0.080、0.285和0.320的酶帶為共有條帶,Rf為0.125、0.165、0.215、0.250、0.345、0.380、0.443和0.500的酶帶為多態(tài)性條帶,多態(tài)位點百分率為72.73%,Rf為0.215的酶帶僅出現(xiàn)在黃花苜蓿Ⅱ的酶譜中。草木樨酶譜共表現(xiàn)出了6條酶帶,Rf為0.015、0.038和0.443的酶帶為共有條帶,Rf為0.285、0.345和0.500的酶帶為多態(tài)性條帶,多態(tài)位點百分率為50.00%。圖1
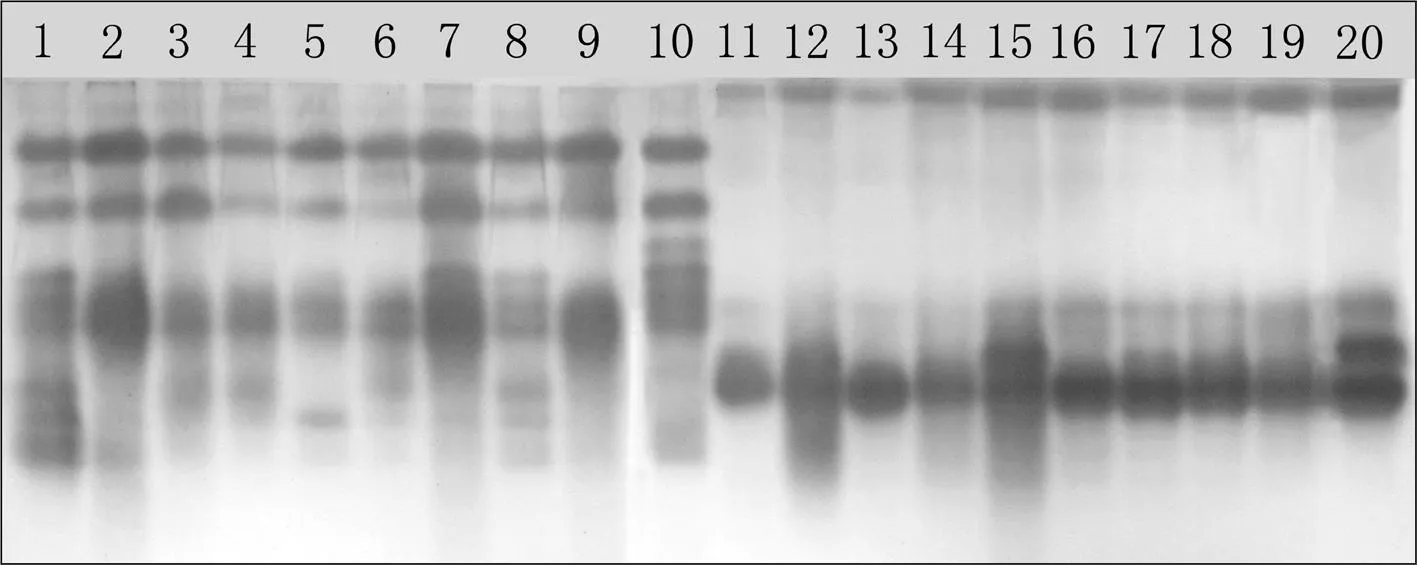
注:圖中序號與表1同
Note:the accession numbers were the same as those in Table 1
圖120份苜蓿屬和草木樨屬材料過氧化物酶的電泳結(jié)果
Fig. 1Electrophoresis results of 20MedicagoandMelilotusaccessions generated by peroxidase electrophoresis
苜蓿屬材料間的酶帶相似率介于30.00%~85.71%,黃花苜蓿Ⅰ和阿爾岡金、新疆大葉和苜蓿王的酶帶相似率最高,為85.71%,黃花苜蓿Ⅰ和雜花苜蓿Ⅰ次之,為83.33%,然后是紫花苜蓿Ⅰ和紫花苜蓿Ⅱ,為80.00%。三得利和黃花苜蓿Ⅱ的酶帶相似率最小,為30.00%。
草木樨屬材料間的酶帶相似率介于0%~100.00%,白花草木樨Ⅰ、白花草木樨Ⅱ、黃花草木樨Ⅱ之間,黃花草木樨Ⅲ、白花草木樨Ⅳ、白花草木樨Ⅴ、黃花草木樨Ⅳ之間的酶帶相似率最高,均為100.00%。黃花苜蓿Ⅱ和白花草木樨Ⅰ、黃花苜蓿Ⅱ和白花草木樨Ⅱ、黃花苜蓿Ⅱ和黃花草木樨Ⅱ的酶帶相似率最小,為0%。
苜蓿屬和草木樨屬材料的酶帶相似率介于0%~44.44%。阿爾岡金和白花草木樨Ⅲ的酶帶相似率最大,為44.44%,阿爾岡金和黃花草木樨Ⅰ次之,為37.50%,然后是紫花苜蓿Ⅰ和白花草木樨Ⅲ、紫花苜蓿Ⅱ和白花草木樨Ⅲ,為36.36%。紫花苜蓿Ⅱ和白花草木樨Ⅰ、紫花苜蓿Ⅱ和白花草木樨Ⅱ、紫花苜蓿Ⅱ和黃花草木樨Ⅱ、紫花苜蓿Ⅲ和白花草木樨Ⅰ、紫花苜蓿Ⅲ和白花草木樨Ⅱ、紫花苜蓿Ⅲ和黃花草木樨Ⅱ、新疆大葉和白花草木樨Ⅰ、新疆大葉和黃花草木樨Ⅰ、新疆大葉和白花草木樨Ⅱ、新疆大葉和黃花草木樨Ⅱ、苜蓿王和白花草木樨Ⅰ、苜蓿王和黃花草木樨Ⅰ、苜蓿王和白花草木樨Ⅱ、苜蓿王和黃花草木樨Ⅱ之間的酶帶相似率最小,為0。表2
2.2相似系數(shù)
供試材料的相似系數(shù)介于0.083~1.000,平均為0.582。苜蓿屬材料的相似系數(shù)介于0.417~0.917,平均為0.698,黃花苜蓿Ⅰ和雜花苜蓿、黃花苜蓿Ⅰ和阿爾岡金、新疆大葉和苜蓿王之間的相似系數(shù)最大,親緣關(guān)系最近;三得利和黃花苜蓿Ⅱ之間的相似系數(shù)最小,親緣關(guān)系最遠。
草木樨屬材料間的相似系數(shù)介于0.667~1.000,平均為0.867,白花草木樨Ⅰ、白花草木樨Ⅱ和黃花草木樨Ⅱ之間,黃花草木樨Ⅲ、白花草木樨Ⅳ、白花草木樨Ⅴ和黃花草木樨Ⅳ之間的相似系數(shù)最大。白花草木樨Ⅰ和白花草木樨Ⅲ、白花草木樨Ⅱ和白花草木樨Ⅲ、黃花草木樨Ⅱ和白花草木樨Ⅲ、黃花草木樨Ⅰ和黃花草木樨Ⅴ之間的相似系數(shù)最小。
苜蓿屬和草木樨屬材料間的相似系數(shù)介于0.083~0.667,平均為0.402,雜花苜蓿和黃花草木樨Ⅲ、三得利和黃花草木樨Ⅲ、雜花苜蓿和白花草木樨Ⅳ、三得利和白花草木樨Ⅳ、雜花苜蓿和白花草木樨Ⅴ、三得利和白花草木樨Ⅴ、雜花苜蓿和黃花草木樨Ⅳ、三得利和黃花草木樨Ⅳ間的相似系數(shù)最大,親緣關(guān)系最近。紫花苜蓿Ⅱ和白花草木樨Ⅰ、新疆大葉和黃花草木樨Ⅰ、紫花苜蓿Ⅱ和白花草木樨Ⅱ、紫花苜蓿Ⅱ和黃花草木樨Ⅱ間的相似系數(shù)最小,親緣關(guān)系最遠。表3
2.3聚類分析
對供試材料進行聚類分析,構(gòu)建聚類圖。結(jié)果表明,20份材料在相似系數(shù)0.402處可聚為2類,第Ⅰ類由苜蓿屬材料組成,在相似系數(shù)0.726處可聚為3個亞類,第Ⅰ亞類包括紫花苜蓿Ⅰ、紫花苜蓿Ⅱ和黃花苜蓿Ⅱ;第Ⅱ亞類包括黃花苜蓿Ⅰ、雜花苜蓿、阿爾岡金、紫花苜蓿Ⅲ和三得利;第Ⅲ亞類包括新疆大葉和苜蓿王。第Ⅱ類由10份草木樨屬材料組成,在相似系數(shù)0.831處可聚為2個亞類,第Ⅰ亞類包括白花草木樨Ⅰ、白花草木樨Ⅱ、黃花草木樨Ⅱ、黃花草木樨Ⅲ、白花草木樨Ⅳ、白花草木樨Ⅴ、黃花草木樨Ⅳ和黃花草木樨Ⅴ;第Ⅱ亞類包括黃花草木樨Ⅰ和白花草木樨Ⅲ。同一類或亞類材料間的親緣關(guān)系較近。圖2
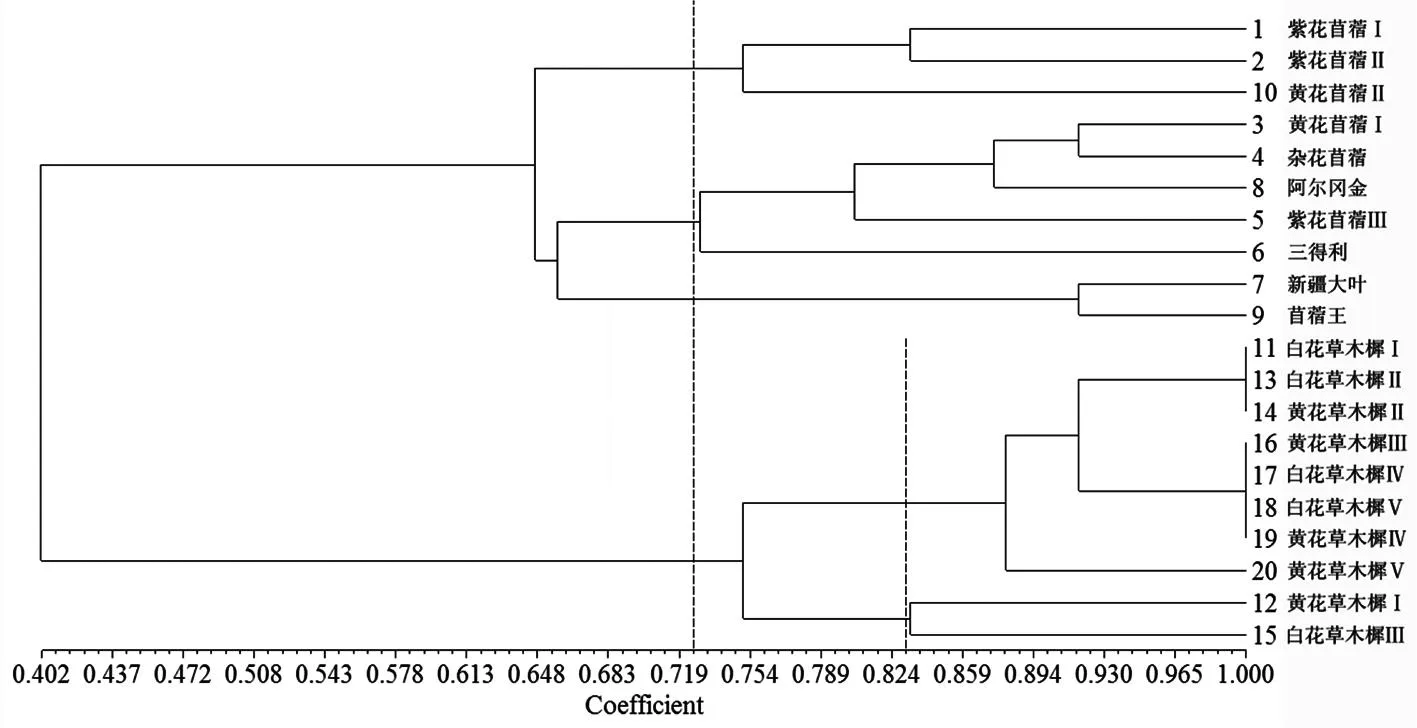
圖220份苜蓿屬和草木樨屬材料基于相似系數(shù)的UPGMA聚類圖
Fig. 2UPGMA dendrogram of 20MedicagoandMelilotusaccessions revealed by cluster analysis based on genetic similarity coefficients
2.4主成分分析
用NTSYSpc 2.1軟件進行主成分分析,構(gòu)建分析圖,圖中材料位置相互靠近者表示親緣關(guān)系較近,遠離者表示親緣關(guān)系較遠。苜蓿屬材料在圖中相對分散,遺傳變異水平較高,部分材料間親緣關(guān)系較遠。草木樨屬材料在圖中相對集中,彼此間親緣關(guān)系較近。白花草木樨Ⅰ、白花草木樨Ⅱ和黃花草木樨Ⅱ,黃花草木樨Ⅲ、白花草木樨Ⅳ、白花草木樨Ⅴ和黃花草木樨Ⅳ在圖中的位置相同,親緣關(guān)系較近,用過氧化物酶不能區(qū)分其遺傳差異。圖3
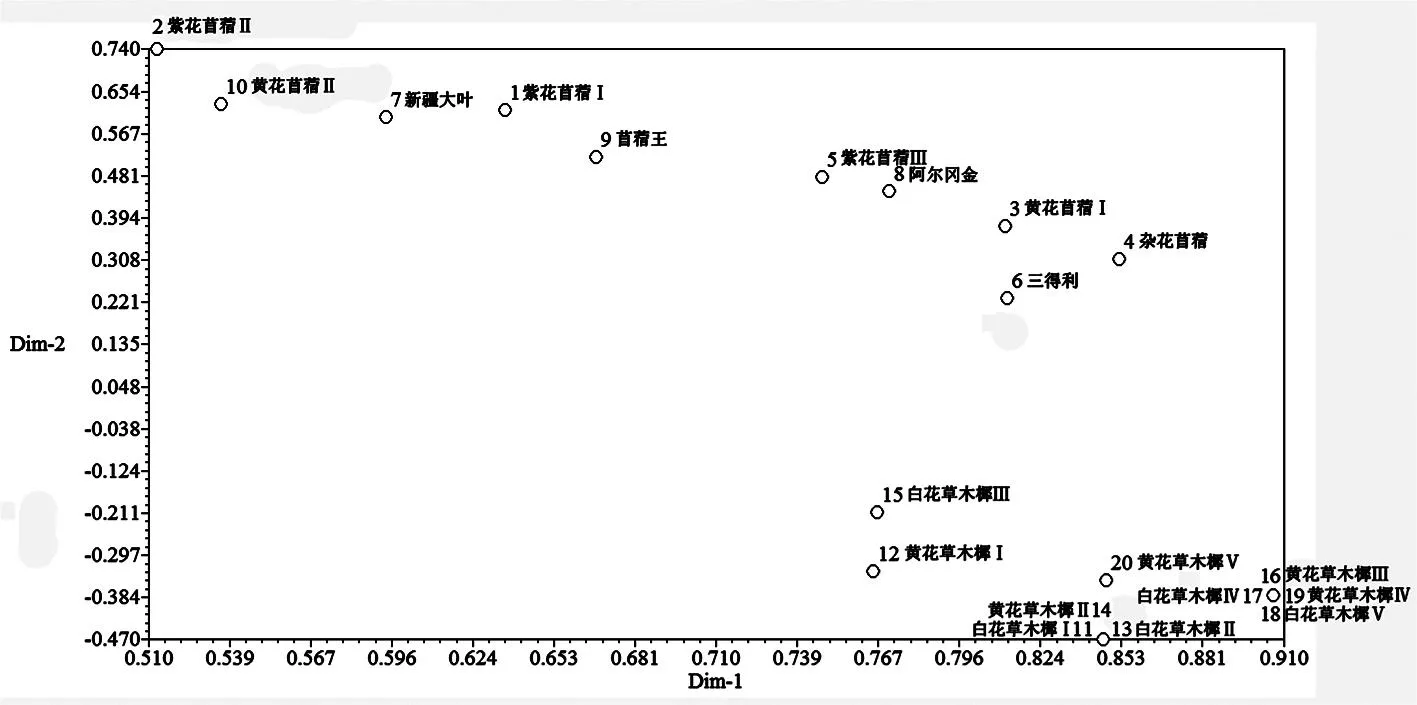
圖320份苜蓿屬和草木樨屬材料基于過氧化物酶的主成分分析二維圖
Fig. 3 Two-dimensional plot of 20MedicagoandMelilotusaccessions as revealed by principal component analysis using peroxidase data
3 討 論
3.1供試種質(zhì)資源的親緣關(guān)系
研究表明,苜蓿屬3個種10份材料間的相似系數(shù)介于0.417~0.917,平均為0.698,與張宇等[5]用SRAP標記研究紫花苜蓿和黃花苜蓿種質(zhì)資源親緣關(guān)系時所得結(jié)果相近,低于陳立強等[14]用過氧化物酶電泳技術(shù)研究紫花苜蓿品種親緣關(guān)系時所得結(jié)果、Talebi等[15]和Mandoulakani等[16]分別用不同分子標記研究紫花苜蓿群體遺傳多樣性時所得結(jié)果、周良彬等[17]用SRAP標記研究雜花苜蓿種質(zhì)資源遺傳多樣性時所得結(jié)果、高素玲等[18]用SDS-聚丙烯酰胺凝膠電泳技術(shù)研究黃花苜蓿種質(zhì)資源遺傳多樣性時所得結(jié)果。與理論推理一致,種間資源的平均相似系數(shù)遠小于種內(nèi)資源,親緣關(guān)系較遠。
與苜蓿屬相比,草木樨屬材料間的相似系數(shù)較高,介于0.667~1.000,平均為0.867,與狄紅艷等[8]用ITS和trnL-trnF序列研究白花草木樨和黃花草木樨種群遺傳多樣性時所得結(jié)果相近。研究結(jié)果還表明,部分草木樨材料間的相似系數(shù)為1.000,親緣關(guān)系太近,用過氧化物酶不能區(qū)分其遺傳差異,與狄紅艷等[8]所得結(jié)論完全相符,總體上,草木樨種質(zhì)資源親緣關(guān)系較近。
苜蓿屬和草木樨屬材料間的相似系數(shù)介于0.083~0.667,平均為0.402,遠低于劉磊等[4]用ISSR標記研究苜蓿屬和胡盧巴屬種質(zhì)資源親緣關(guān)系時所得結(jié)果,與胡盧巴屬相比,苜蓿屬和草木樨屬種質(zhì)資源的親緣關(guān)系相對較遠。
3.2種質(zhì)資源地理分布與親緣關(guān)系的相關(guān)性
雜交和漸滲是生物界普遍存在的現(xiàn)象,是生物遺傳變異的重要途徑,在物種進化中扮演著重要角色[19-20],有研究表明,至少25%的植物存在種間雜交和基因漸滲現(xiàn)象[21]。紫花苜蓿、黃花苜蓿、白花草木樨和黃花草木樨均屬異花授粉植物[3,22],可借助風力和昆蟲傳粉,為其遺傳物質(zhì)在自然條件下相互滲透創(chuàng)造了條件。供試材料中,雜花苜蓿和黃花苜蓿Ⅰ、白花草木樨Ⅱ和黃花草木樨Ⅱ、白花草木樨Ⅳ和黃花草木樨Ⅲ兩兩間的采集地相同,在聚類分析中,也分別被聚于相同亞類,彼此親緣關(guān)系最近,可能與長期的種間基因漸滲有關(guān)。結(jié)合親緣關(guān)系和分布地區(qū),巴里坤雜花苜蓿可能就是黃花苜蓿Ⅰ和當?shù)刈匣ㄜ俎T耘嗥贩N通過開放授粉產(chǎn)生的野生群體。供試材料中也有部分種質(zhì)資源的親緣關(guān)系與地理來源沒有明顯的關(guān)系,這與張宇等[5]研究紫花苜蓿和黃花苜蓿種質(zhì)資源遺傳多樣性時所得結(jié)論相同,這些種質(zhì)資源在長期的進化過程中形成了完全的生殖隔離,即使分布地區(qū)相同,花期相遇,基因滲透程度也相對較小。


3.3苜蓿和草木樨屬間雜交的可行性論證和親和性預測
隨著種內(nèi)資源的日趨枯竭,通過遠緣雜交實現(xiàn)種質(zhì)創(chuàng)新越來越受到人們的重視。目前,我國在苜蓿遠緣雜交領(lǐng)域進行了大量研究和實踐,取得了豐碩成果。通過紫花苜蓿和黃花苜蓿雜交,育成了甘農(nóng)1號、草原1號、草原2號、圖牧1號、赤草1號和新牧1號苜蓿;通過扁蓿豆和紫花苜蓿雜交,育成了龍牧801苜蓿和龍牧803苜蓿[23-25]。隨著生物工程技術(shù)的迅速發(fā)展,遠緣雜交也逐漸成為各種育種技術(shù)相互滲透,相互綜合的焦點。毛雅妮[26]對苜蓿和草木樨體細胞融合的基礎(chǔ)條件進行了研究,為體細胞融合奠定了基礎(chǔ)。賀輝[11]建立了完整的紫花苜蓿、草木樨組培體系,對紫花苜蓿、白花草木樨原生質(zhì)體融合的條件進行了研究,并以PEG為融合劑,誘導產(chǎn)生了紫花苜蓿和白花草木樨雜種細胞和雜種愈傷組織。基于現(xiàn)代生物技術(shù),苜蓿和草木樨雜交已初見成效,然而,目前的研究多集中于雜交和培養(yǎng),親本選配方面的研究鮮見報道,實踐表明,正確選配親本,是遠緣雜交能否取得成功的關(guān)鍵,通常親本間親緣關(guān)系愈近,雜交成功的可能性就愈大[3]。黃花苜蓿Ⅰ和雜花苜蓿、黃花苜蓿Ⅰ和阿爾岡金之間,白花草木樨Ⅰ、白花草木樨Ⅱ和黃花草木樨Ⅱ之間,黃花草木樨Ⅲ、白花草木樨Ⅳ、白花草木樨Ⅴ和黃花草木樨Ⅳ之間,雜花苜蓿和黃花草木樨Ⅲ、三得利和黃花草木樨Ⅲ、雜花苜蓿和白花草木樨Ⅳ、三得利和白花草木樨Ⅳ、雜花苜蓿和白花草木樨Ⅴ、三得利和白花草木樨Ⅴ、雜花苜蓿和黃花草木樨Ⅳ、三得利和黃花草木樨Ⅳ之間的相似系數(shù)相對較大,親緣關(guān)系較近,雜交成功的可能性較大。
同工酶是基因表達的產(chǎn)物[13],親本間共有的酶帶數(shù)越多,基因的相似程度就越大,理論上,遠緣雜交的親和性就越高。從酶帶相似程度分析,苜蓿材料中,黃花苜蓿Ⅰ和阿爾岡金酶帶相似率最高;草木樨材料中,白花草木樨Ⅰ、白花草木樨Ⅱ、黃花草木樨Ⅱ之間,黃花草木樨 Ⅲ、白花草木樨Ⅳ、白花草木樨Ⅴ、黃花草木樨Ⅳ之間的酶帶相似率最高;苜蓿和草木樨材料中,阿爾岡金和白花草木樨Ⅲ的酶帶相似率最高,酶帶相似率越高,基因相似程度較高,雜交成功的可能性較大。
4結(jié) 論
供試種質(zhì)資源中,草木樨屬材料的平均相似系數(shù)最大,在主成分分析圖中的位置相對集中,大部分材料間的親緣關(guān)系相對較近。與之相比,苜蓿屬材料的平均相似系數(shù)較小,在主成分分析圖中的位置分散,材料間的親緣關(guān)系相對較遠。苜蓿屬和草木樨屬材料間的平均相似系數(shù)最小,在聚類分析中,供試種質(zhì)資源被聚為兩類,苜蓿屬材料為一類,草木樨屬材料為一類,大部分材料間親緣關(guān)系較遠。
苜蓿材料中,黃花苜蓿Ⅰ和雜花苜蓿、黃花苜蓿Ⅰ和阿爾岡金之間親緣關(guān)系較近,雜交成功的可能性較大。草木樨材料中,白花草木樨Ⅰ、白花草木樨Ⅱ和黃花草木樨Ⅱ之間,黃花草木樨Ⅲ、白花草木樨Ⅳ、白花草木樨Ⅴ和黃花草木樨Ⅳ之間親緣關(guān)系較近,雜交成功的可能性較大。苜蓿和草木樨材料中,雜花苜蓿和黃花草木樨Ⅲ、三得利和黃花草木樨Ⅲ、雜花苜蓿和白花草木樨Ⅳ、三得利和白花草木樨Ⅳ、雜花苜蓿和白花草木樨Ⅴ、三得利和白花草木樨Ⅴ、雜花苜蓿和黃花草木樨Ⅳ、三得利和黃花草木樨Ⅳ之間親緣關(guān)系較近,雜交成功的可能性較大;阿爾岡金和白花草木樨Ⅲ的酶帶相似率最高,基因相似程度較高,雜交成功的可能性較大。
參考文獻(References)
[1] 孫啟忠,玉柱,馬春暉,等.我國苜蓿產(chǎn)業(yè)過去10年發(fā)展成就與未來10年發(fā)展重點[J].草業(yè)科學,2013,30(3):471-477.(in Chinese)
SUN Qi-zhong, YU Zhu, MA Chun-hui, et al. (2013).Achievements of the alfalfa industry in last decade and priorities in next decade in China [J].PrataculturaeSinica, 30(3): 471-477. (in Chinese)
[2] 劉慧,席琳喬,吳曉樂,等.24個草木樨品種生產(chǎn)性能比較研究[J].草食家畜,2014,(1):54-59.
LIU Hui, XI Lin-qiao, WU Xiao-le, et al. (2014).24 varieties of sweet clover introduction experiment comparison [J].Grass-FeedingLivestock, (1):54-59.(in Chinese)
[3] 云錦鳳.牧草及飼料作物育種學[M].北京:中國農(nóng)業(yè)出版社,2001:69-71.
YUN Jin-feng.(2001).BreedingScienceofGrassandForageCrops[M]. Beijing: China Agriculture Press: 69-71.(in Chinese)
[4] 劉磊,王宗禮,李志勇.利用ISSR標記解析紫花苜蓿、黃花苜蓿和胡盧巴屬植物的親緣關(guān)系[J].安徽農(nóng)業(yè)科學,2012,40(25):12 393-12 395.
LIU Lei, WANG Zong-li, LI Zhi-yong, et al.(2012).Analysis on genetic relationship amongMedicagosativa,MedicagofalcataandTrigonellafoenum-graecum using ISSR [J].JournalofAnhuiAgriculturalSciences, 40(25):12,393-12,395.(in Chinese)
[5] 張宇,于林清,慈忠玲.利用SRAP標記研究紫花苜蓿和黃花苜蓿種質(zhì)資源遺傳多樣性[J].中國草地報,2014,31(6):1 070-1 079.
ZHANG Yu, YU Lin-qing, CI Zhong-ling.(2014).Genetic diversity of germplasm resources based on SRAP markers [J].ChineseJournalofGrassland, 31(6):1,070-1,079.(in Chinese)
[6] 李飛飛,崔大方,羊海軍,等.中國新疆紫花苜蓿復合體3個種的遺傳多樣性及親緣關(guān)系研究[J].草業(yè)學報,2012,21(1):190-198.
LI Fei-fei, CUI Da-fang, YANG Hai-jun, et al.(2012).Genetic diversity and genetic relationships among three species ofMedicagosativacomplex in Xingjiang province of China [J].PrataculturaeSinica, 21(1):190-198. (in Chinese)
[7] Bagavathiannan, M. V., Julier, B., Barre, P., Gulden, R. H., & Van Acker, R. C. (2010). Genetic diversity of feral alfalfa (MedicagosativaL.) populations occurring in Manitoba, Canada and comparison with alfalfa cultivars: an analysis using SSR markers and phenotypic traits.Euphytica, 173(3): 419-432.
[8] 狄紅艷,駱凱,張吉宇,等.基于ITS和trnL-trnF序列的草木樨種群遺傳多樣性研究[J].西北植物學報,2014,34(2):265-269.
DI Hong-yan, LUO Kai, ZHANG Ji-yu, et al. (2014).Genetic diversity analysis of Melilotus populations based on ITS and trnL-trnF sequences [J].ActaBotanicaBoreali-OccidentaliaSinica, 34(2): 265-269.(in Chinese)
[9] 孫啟忠,王宗禮,徐麗君.旱區(qū)苜蓿[M].北京:科學出版社,2008:25-26.
SUN Qi-zhong, WANG Zong-li, XU Li-jun. (2014).AlfalfainAridRegions[M]. Beijing: Science Press: 25-26. (in Chinese)
[10] 曲善民,馮乃杰,鄭殿峰,等.苜蓿與草木樨種子生物學特征特性比較研究[J].黑龍江八一農(nóng)墾大學學報,2011,23(2):5-7.
QU Shan-min, FENG Nai-jie, ZHENG Dian-feng, et al. (2011).Alfalfa and sweet clover seed comparative study of biological characteristics [J].JournalofHeilongjiangBayiAgriculturalUniversity, 23(2):5-7. (in Chinese)
[11] 賀輝.紫花苜蓿和白花草木樨細胞融合技術(shù)研究[D].蘭州:甘肅農(nóng)業(yè)大學碩士畢業(yè)論文,2008.
HE Hui.(2008).StudyoncellfusiontechnologyofMedicagosativaandMelilotussuaveolen[D].Master Dissertation. Gansu Agricultural University, Lanzhou.(in Chinese)
[12] Moghaddam, M., Ehdaie, B., & Waines, J. G. (2000). Genetic diversity in populations of wild diploid wheat Triticum urartu Tum. ex. Gandil. revealed by isozyme markers.GeneticResourcesandCropEvolution, 47(3):323-334.
[13] 周延清,楊清香,張改娜.生物遺傳標記與應用[M].北京:化學工業(yè)出版社,2008:47-54.
ZHOU Yan-qing, YANG Qing-xiang, ZHANG Gai-na.(2008).BiologicalGeneticMarkersandItsUtilization[M]. Beijing: Chemical Industry Press:47-54. (in Chinese)
[14] 陳立強,馬春暉,周海寧,等.南疆地區(qū)引進苜蓿品種親緣關(guān)系的過氧化物酶分析[J].新疆農(nóng)業(yè)科學,2014,51(8):1 496-1 503.
CHEN Li-qiang, MA Chun-hui, ZHOU Hai-ning, et al. (2014).Genetic relationship among introduced alfalfa varieties in south Xinjiang revealed by peroxidase [J].XinjiangAgriculturalSciences, 51(8):1,496-1,503.(in Chinese)
[15] Talebi, M., Hajiahmadi, Z., & Rahimmalek, M. (2011). Genetic diversity and population structure of four Iranian alfalfa populations revealed by sequence-related amplified polymorphism (SRAP) markers.JournalofCropScienceandBiotechnology, 14(3):173-178.
[16] Mandoulakani, B. A., Piri, Y., Darvishzadeh, R., Bernoosi, I., & Jafari, M. (2012). Retroelement insertional polymorphism and genetic diversity inMedicagosativapopulations revealed by IRAP and REMAP markers.PlantMolecularBiologyReporter, 30(2):286-296.
[17] 周良彬,盧欣石,王鐵梅,等.雜花苜蓿種質(zhì)SRAP標記遺傳多樣性研究[J].草地學報,2010,18(4):544-549.
ZHOU Liang-bin, LU Xin-shi, WANG Tie-mei, et al. (2010).Genetic diversity ofMedicagovariaMartyn. germplasms detected by SRAP markers[J].ActaAgrestiaSinica,18(4):544-549.(in Chinese)
[18] 高素玲,郭麗,朱飛雪,等.黃花苜蓿不同種質(zhì)材料的遺傳多樣性研究[J].中國草地學報,2013,35(1):61-66.
GAO Su-ling, GUO Li, ZHU Fei-xue, et al.(2013).Study on genetic diversity ofMedicagofalcateL.germplasms [J].ChineseJournalofGrassland, 35(1):61-66.(in Chinese)
[19] Tovar-Sánchez, E., & Oyama, K. (2004). Natural hybridization and hybrid zones between Quercus crassifolia and Quercus crassipes (Fagaceae) in Mexico: morphological and molecular evidence.AmericanJournalofBotany, 91(9):1,352-1,363.
[20] Van Droogenbroeck, B., Kyndt, T., Romeijn-Peeters, E., Van Thuyne, W., Goetghebeur, P., Romero-Motochi, J. P., & Gheysen, G. (2006). Evidence of natural hybridization and introgression between Vasconcellea species (Caricaceae) from southern Ecuador revealed by chloroplast, mitochondrial and nuclear DNA markers.Annalsofbotany, 97(5):793-805.
[21] Mallet, J. (2005). Hybridization as an invasion of the genome.TrendsinEcology&Evolution, 20(5):229-237.
[22] Turkington, R. A., Cavers, P. B., & Rempel, E. (1978). The Biology of Canadianweeds.: 29.MelilotusalbaDesr. andM.officinalis(L.) Lam.CanadianJournalofPlantScience, 58(2):523-537.
[23] 曹致中.優(yōu)質(zhì)苜蓿栽培與利用[M].北京:中國農(nóng)業(yè)出版社,2001:24-28.
CAO Zhi-zhong. (2001).CultivationandUtilizationofQualityAlfalfa[M]. Beijing: China Agriculture Press:24-28. (in Chinese)
[24] 王殿魁,李紅,羅新義.扁蓿豆與紫花苜蓿雜交育種研究[J].草地學報,2008,16(5):458-465.
WANG Dian-kui, LI Hong, LUO Xin-yi. (2008).Crossbreeding ofMelilotoidesruthenicusandMedicagosativa[J].ActaAgrestiaSinica, 16(5):458-465. (in Chinese)
[25] 王雪,李志萍,孫建軍,等.中國苜蓿品種的選育與研究[J].草業(yè)科學,2014,31(3):512-518.
WANG Xue, LI Zhi-ping, SUN Jian-jun, et al.(2014).Progress of alfalfa breeding in China [J].PrataculturalScience,31(3):512-518.(in Chinese)
[26] 毛雅妮.苜蓿和草木樨體細胞融合的基礎(chǔ)研究[D].蘭州:甘肅農(nóng)業(yè)大學碩士畢業(yè)論文,2010.
MAO Ya-ni. (2010).Fundamentalstudiesonsomaticfusionofalfalfaandsweetclover[D]. Master Dissertation. Gansu Agricultural University, Lanzhou.(in Chinese)
Genetic Relationship amongMedicagoandMelilotusGermplasm
Resources Revealed by Peroxidase Electrophoresis
CHEN Li-qiang1, BIAN Yan-xia2, MA Chun-hui3,XU Li-ming1,ZHAO Hao-bo1,YANG Hao-hong1
(1.CollegeofAnimalScience,TarimUniversity/KeyLaboratoryofTarimAnimalHusbandryScienceandTechnology,XinjiangProduction&ConstructionCorps,AlarXinjiang843300,China; 2.LogisticManagementOffice,TarimUniversity,AlarXinjiang843300,China; 3.CollegeofAnimalScienceandTechnology,ShiheziUniversity,ShiheziXinjiang832000,China)
Abstract:【Objective】 This study aims to analyze the generic relationships among Medicago and Melilotus germplasm resources and enhance the predictability of distant hybridization of those accessions.【Method】The genetic relationships among 20 accessions of 3 Medicago species and 2 Melilotus species were investigated using peroxidase electrophoresis.【Result】The band similar rates within and between Medicago and Melilotus accessions ranged from 30.00% to 83.33%, 33.33% to 100.00% and 0% to 44.44%, respectively. The similarity coefficients of those accessions ranged from 0.417 to 0.917, 0.667 to 1.000 and 0.083 to 0.667, respectively. The Medicago and Melilotus accessions could be classified into two separate groups at the similarity coefficient of 0.402 based on cluster analysis. Compared to Melilotus accessions, the Medicago accessions in principal component analysis plot were relatively dispersed, indicating that the level of genetic variation within Medicago species was relatively higher.【Conclusion】Some accessions from different species and genera had relatively close genetic relationships and high band similar rates, therefore, the hybridization among them may be easy to succeed.
Key words:Medicago; Melilotus; peroxidase; genetic relationship; distant hybridization
中圖分類號:S812.3
文獻標識碼:A
文章編號:1001-4330(2016)03-0523-10
作者簡介:陳立強(1981-),男,甘肅會寧人,講師,碩士,研究方向為牧草種質(zhì)資源及遺傳育種,(E-mail)clqdky@126.com通訊作者:馬春暉(1966-),男,新疆哈密人,教授,博士,研究方向為牧草生產(chǎn)與加工,(E-mail)chunhuima@126.com
基金項目:新疆生產(chǎn)建設(shè)兵團塔里木畜牧科技重點實驗室開放課題(HS201405);塔里木大學校長基金項目(TDZKQN201502);新疆生產(chǎn)建設(shè)兵團塔里木畜牧科技重點實驗室開放課題(HS201103);國家牧草產(chǎn)業(yè)技術(shù)體系(CARS-35)
收稿日期:2015-07-09
doi:10.6048/j.issn.1001-4330.2016.03.019
Fund project:Open Projects of Key Laboratory of Tarim Animal Husbandry Science & Technology (HS201405); Principal Foundation Project of Tarim University (TDZKQN201502); Open Projects of Key Laboratory of Tarim Animal Husbandry Science & Technology (HS201103); National Forage Industry Technological System (CARS-35).
