UASB反應(yīng)器中產(chǎn)甲烷菌對溫度脅迫的響應(yīng)
2016-05-27 07:32:45張立國班巧英李建政山西大學(xué)環(huán)境與資源學(xué)院山西太原030006哈爾濱工業(yè)大學(xué)市政與環(huán)境工程學(xué)院黑龍江哈爾濱50090
中國環(huán)境科學(xué) 2016年4期
張立國,班巧英*,李建政(.山西大學(xué)環(huán)境與資源學(xué)院,山西 太原 030006;.哈爾濱工業(yè)大學(xué)市政與環(huán)境工程學(xué)院,黑龍江 哈爾濱 50090)
?
UASB反應(yīng)器中產(chǎn)甲烷菌對溫度脅迫的響應(yīng)
張立國1,班巧英1*,李建政2(1.山西大學(xué)環(huán)境與資源學(xué)院,山西 太原 030006;2.哈爾濱工業(yè)大學(xué)市政與環(huán)境工程學(xué)院,黑龍江 哈爾濱 150090)
摘要:采用間歇培養(yǎng)方式探討了升流式厭氧污泥床(UASB)反應(yīng)器中不同營養(yǎng)類型產(chǎn)甲烷菌對溫度脅迫的響應(yīng)規(guī)律.研究結(jié)果表明,產(chǎn)甲烷螺旋菌(Methanospirillum)是該反應(yīng)器的主要氫營養(yǎng)型產(chǎn)甲烷菌,而主要乙酸營養(yǎng)型產(chǎn)甲烷菌為產(chǎn)甲烷絲狀菌(Methanosaeta).在35℃條件下,氫營養(yǎng)型和乙酸營養(yǎng)型產(chǎn)甲烷菌的累計甲烷產(chǎn)量分別為24.7, 11.7mL,而最大產(chǎn)甲烷速率分別為0.74, 0.18mL/h.當溫度從35℃分別降低至30, 25, 20, 15℃時,導(dǎo)致氫營養(yǎng)型產(chǎn)甲烷菌的累計產(chǎn)甲烷量分別減少了14.2%, 34.0%, 47.0%, 57.5%,而乙酸營養(yǎng)型產(chǎn)甲烷菌的累計產(chǎn)甲烷量分別減少了5.1%, 23.9%, 45.3%, 95.7%.由此可見,在20~30℃時氫營養(yǎng)型產(chǎn)甲烷菌對溫度脅迫更加敏感,而在15℃以下時乙酸營養(yǎng)營養(yǎng)型產(chǎn)甲烷菌對溫度脅迫更加敏感.
關(guān)鍵詞:UASB反應(yīng)器;產(chǎn)甲烷菌;溫度脅迫;產(chǎn)甲烷量
* 責(zé)任作者, 講師, banqiaoying@163.com
有機物厭氧生物處理是在一系列微生物的協(xié)同代謝作用下完成的,包括水解發(fā)酵菌群,產(chǎn)氫產(chǎn)乙酸菌,同型產(chǎn)乙酸菌和產(chǎn)甲烷菌[1-3].其中,產(chǎn)甲烷菌位于食物鏈的最末端,在產(chǎn)甲烷系統(tǒng)中占有獨特的生態(tài)位[4].由于產(chǎn)甲烷菌只能利用H2/CO2,乙酸,甲酸,甲胺等簡單的底物進行生長代謝,且增殖緩慢,對環(huán)境因子變化亦較敏感,因此通常被認為是厭氧生物處理過程的限速步驟[5-6].溫度是厭氧微生物重要生態(tài)因子之一,它不僅能夠影響微生物的生長速率,同時可能會改變微生物的活性、豐富度等[7-8].另外,溫度變化可能會引起系統(tǒng)pH值下降和游離氨濃度的增加,進而影響水解,產(chǎn)酸發(fā)酵和產(chǎn)甲烷效率,最終影響到厭氧生物處理反應(yīng)器效能及其穩(wěn)定性[9-10].
目前,關(guān)于溫度變化對厭氧反應(yīng)器中不同營養(yǎng)型產(chǎn)甲烷菌活性影響的報道較少,本研究考察了產(chǎn)甲烷菌群對溫度脅迫的響應(yīng)規(guī)律,為厭氧生物處理反應(yīng)器運行調(diào)控提供參考.
1 材料與方法
1.1 接種污泥
接種污泥取自本實驗室正在運行的升流式厭氧污泥床反應(yīng)器(UASB),該反應(yīng)器長期處理丙酸合成廢水(以丙酸為唯一碳源).UASB的運行情況,溫度35℃,水力停留時間(HRT)10h,進水COD 3000mg/L,比COD去除率和比甲烷生成率分別為1.4kgCOD/(kgVSS·d)和285.3LCH4/ (kgCODremoved· d).
1.2 靜態(tài)搖瓶試驗
靜態(tài)搖瓶試驗采用間歇培養(yǎng)方式進行,實驗容器為150mL血清瓶.每個血清瓶中加入10mLUASB反應(yīng)器中的厭氧顆粒污泥(MLVSS為2.4g/L)和40mL液體培養(yǎng)基,其中碳源分別為H2/CO2(150mL,體積比4:1)或乙酸(500mg/L).將培養(yǎng)液pH值用1mol/L HCl 或1mol/L NaOH調(diào)至7.0.然后將血清瓶分別置于5個恒溫空氣浴搖床(15,20,25,30,35℃)進行靜態(tài)搖甁試驗.每個溫度梯度做3個平行樣,數(shù)據(jù)分析取其平均值.每12h測定一次產(chǎn)氣量,氣體組成,每24h測定一次揮發(fā)酸組成.
1.3 DNA提取和qPCR分析
取0.15g污泥樣品用磷酸緩沖液沖洗若干次進行DNA提取.基因組DNA提取采用DNA提取試劑盒(MO Bio Laboratories,Inc.,Carlsbad,CA, USA).以前的研究發(fā)現(xiàn),該UASB反應(yīng)器中的產(chǎn)甲烷菌主要來自產(chǎn)甲烷絲狀菌屬(Methanosaeta)和產(chǎn)甲烷螺旋菌屬(Methanospirillum)[11].因此,對這兩個屬的產(chǎn)甲烷菌進行定量分析.所用引物分別為,Methanosaeta,5′-GCATCGAGATTTAAAG CTC-3′,5′-AGCCAGATTTGTAACCTGGC-3′;M ethanospirillum,5′-AACGGCCCAACAAGCCT GTC-3′,5′-GGACTTGTATATCCAGCA CG-3′.
本實驗中的定量分析采用絕對定量方式.標準樣品為含目的基因序列的重組pMD18-T質(zhì)粒.將標準樣品進行10倍梯度稀釋(101~107).標準樣品和待測樣品同時進行實時熒光定量PCR檢測(qPCR).qPCR在熒光定量分析系統(tǒng)(Model 7500,ABI,Foster,CA,USA)中完成,所用熒光染料為SYBR green.反應(yīng)體系:引物1μmol/L,SYBR Green Mix 10μL(Toyobo Co.,LTD.,Japan),H2O 5.96μL,ROX 0.04μL,DNA模板3μL.反應(yīng)程序:94℃預(yù)變性1min,94℃ 15s,58℃ 30s,72℃ 35s數(shù)據(jù)采集,40個循環(huán).每個qPCR反應(yīng)設(shè)置3個重復(fù).以達到對數(shù)增長時的循環(huán)數(shù)(Ct值)為橫坐標,標準質(zhì)粒數(shù)目的對數(shù)值為縱坐標繪制標準曲線,得到關(guān)于Methanospirillum和Methanosaeta的一元線性回歸方程分別為y=-0.3051x+11.626 (R2= 0.9929),y=-0.2669x+10.455 (R2=0.9982).
1.4 分析項目及方法
生物量(揮發(fā)性懸浮固體總量MLVSS)采用標準方法測定[12],甲烷發(fā)酵產(chǎn)氣量通過10~50mL的玻璃注射器排氣計量.發(fā)酵氣體的組分和揮發(fā)酸濃度分別采用山東魯南瑞虹化工儀器有限公司的SP-6800A型(TCD檢測器)和SP-6890型(FID檢測器)氣相色譜測定.累計甲烷產(chǎn)量參照Owen法進行計算[13].
1.5 動力學(xué)分析
不同溫度下的產(chǎn)甲烷動力學(xué)分析參照Gompertz模型.公式如式(1):
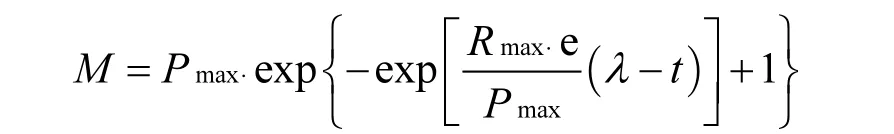
其中:M為反應(yīng)t(h)累計甲烷產(chǎn)量,mL;Pmax為最大產(chǎn)甲烷量,mL;Rmax為最大產(chǎn)甲烷速率,mL/h;,λ為延遲時間,h.將累計甲烷產(chǎn)量和相應(yīng)反應(yīng)時間代入公式,用統(tǒng)計軟件Origin 8.0計算出Pmax、Rmax、λ[14].
2 結(jié)果與討論
2.1 產(chǎn)甲烷菌的定量分析
在進水COD 3000mg/L和35℃條件下, UASB 的COD去除率為95%,比產(chǎn)甲烷速率為0.26L/g CODremoved,說明一些較高活性的產(chǎn)甲烷菌存在于該系統(tǒng)中.基于16S rRNA基因序列的PCR-DGGE分析表明,產(chǎn)甲烷絲狀菌屬(Methanosaeta)為該系統(tǒng)中優(yōu)勢乙酸營養(yǎng)型產(chǎn)甲烷菌,而優(yōu)勢氫營養(yǎng)型產(chǎn)甲烷菌為產(chǎn)甲烷螺旋菌屬(Methanospirillum)[11].產(chǎn)甲烷絲狀菌只能利用乙酸作為底物,而產(chǎn)甲烷螺旋菌只能利用H2/CO2作為底物[15-16].為了考察UASB中這兩類產(chǎn)甲烷菌的數(shù)量,對它們進行了qPCR分析.結(jié)果表明,Methanosaeta的數(shù)量為2.7×10516SrRNA基因拷貝數(shù)/ngDNA.Methanospirillum的數(shù)量比Methanosaeta少了2個數(shù)量級,只有1.8×10316S rRNA基因拷貝數(shù)/ng DNA.已有研究結(jié)果表明,在乙酸含量較低的生物反應(yīng)器中Methanosaeta為優(yōu)勢乙酸營養(yǎng)型產(chǎn)甲烷菌,而且Methanosaeta在顆粒污泥中的含量要高于絮狀污泥[17-18].這可能是由于產(chǎn)甲烷絲狀菌特殊的形態(tài)有利于顆粒污泥形成的原因.
2.2 甲烷生成對溫度脅迫的響應(yīng)規(guī)律
定量分析的結(jié)果表明,氫營養(yǎng)型產(chǎn)甲烷菌和乙酸營養(yǎng)型產(chǎn)甲烷菌分別來自產(chǎn)甲烷螺旋菌屬和產(chǎn)甲烷絲狀菌屬.H2/CO2和乙酸不僅是產(chǎn)甲烷菌可利用的主要底物,而且是有機物厭氧生物處理過程中重要的中間代謝產(chǎn)物.為了了解溫度對不同營養(yǎng)類型產(chǎn)甲烷菌活性的影響,本研究分別以H2/CO2和乙酸作為底物考察溫度降低對氫營養(yǎng)型產(chǎn)甲烷菌和乙酸營養(yǎng)型產(chǎn)甲烷菌活性的影響.圖1表明,在35℃條件下,接種培養(yǎng)60h后累計產(chǎn)甲烷量就達到了最大值24.7mL.當溫度從35℃分別降低至30℃和25℃時,甲烷生成速率降低,最終累計產(chǎn)甲烷量比對照(35℃)減少了14.2%和34.0%(表1).
當溫度分別降低至20℃和15℃時(圖1),甲烷的生成出現(xiàn)了明顯的停滯期,該停滯期約為24~28h,表明低溫對氫營養(yǎng)型產(chǎn)甲烷菌代謝活性造成了顯著抑制作用.經(jīng)過24~28h的停滯期后,累積產(chǎn)甲烷量開始逐漸增加,表明氫營養(yǎng)型產(chǎn)甲烷菌經(jīng)過24h的適應(yīng),其代謝活性緩慢恢復(fù).累計產(chǎn)甲烷量最后分別穩(wěn)定在13.1,10.5mL左右.值得注意的是,20和15℃時的累積產(chǎn)甲烷趨勢與35,30,25℃時的累積產(chǎn)甲烷趨勢明顯不同.20℃和15℃時出現(xiàn)了明顯的停滯期,更重要的是累積產(chǎn)甲烷量呈線性增加趨勢,而35,30,25℃時的累積產(chǎn)甲烷量呈現(xiàn)的是先線性增加,后緩慢增加(平臺期)的趨勢.這2種累積產(chǎn)甲烷趨勢的不同,意味著在20℃和15℃繼續(xù)延長培養(yǎng)時間,產(chǎn)甲烷量會持續(xù)增加,而在35,30,25℃條件下產(chǎn)甲烷量則不會持續(xù)增加.由此可以看出,低溫對氫營養(yǎng)型產(chǎn)甲烷菌的脅迫是通過控制半速度常數(shù)Ks來實現(xiàn)的.隨著溫度的下降,Ks迅速增加,使得氫營養(yǎng)型產(chǎn)甲烷菌對H2/CO2轉(zhuǎn)化速率顯著下降,最終導(dǎo)致甲烷產(chǎn)量的下降.
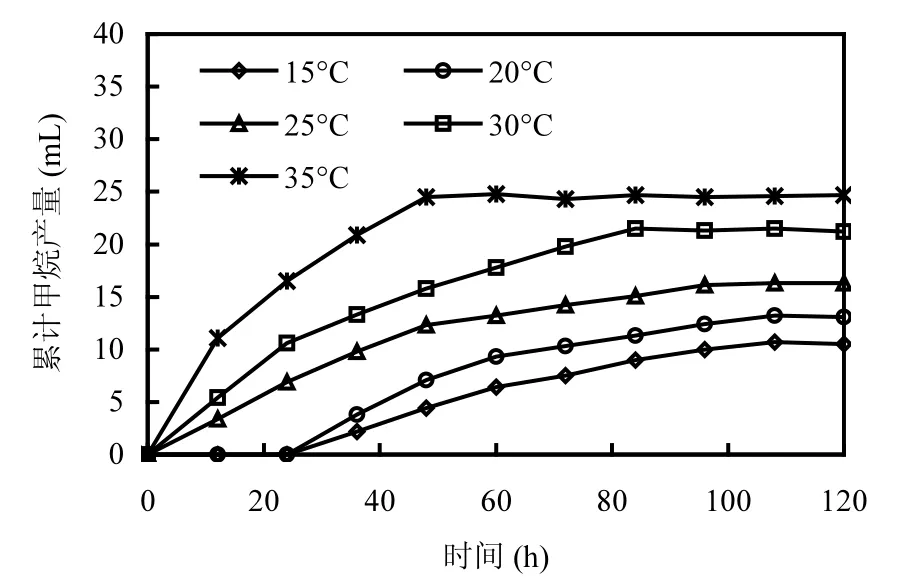
圖1 不同溫度條件下氫營養(yǎng)型產(chǎn)甲烷菌甲烷產(chǎn)量Fig.1 CH4 production of hydrogenotrophic methanogens at different temperature conditions
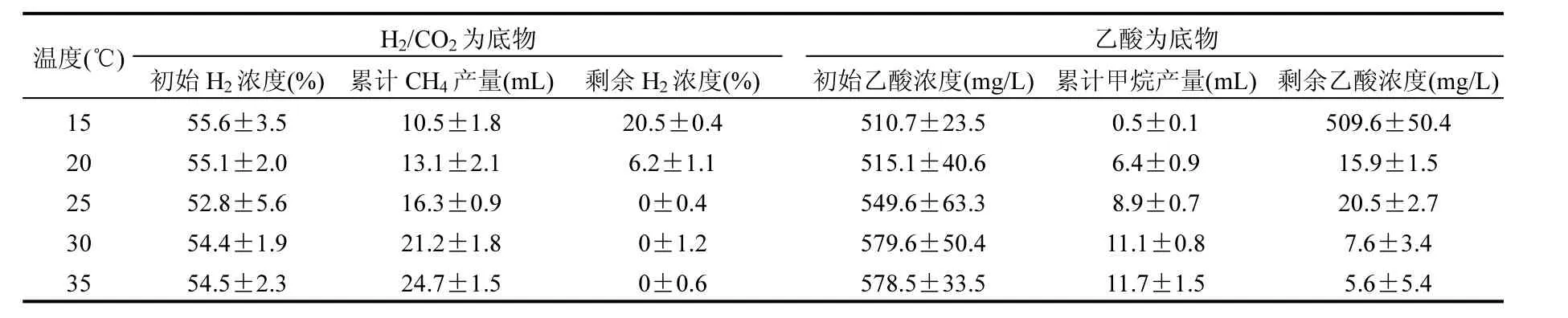
表1 不同溫度條件下的產(chǎn)甲烷特征Table 1 Methanogenic characteristics at different temperature conditions
如圖2所示,當以乙酸為底物時,產(chǎn)甲烷菌在35℃條件下表現(xiàn)出良好的活性,在接種培養(yǎng)84h后累計產(chǎn)甲烷量達到最大值11.7mL.溫度從35℃降低至30,25,20,15℃導(dǎo)致產(chǎn)甲烷速率顯著降低.在30,25,20℃,15℃條件下的累計產(chǎn)甲烷量比對照(35 ℃)分別減少了5.1%,23.9%,45.3%,95.7%(表1).
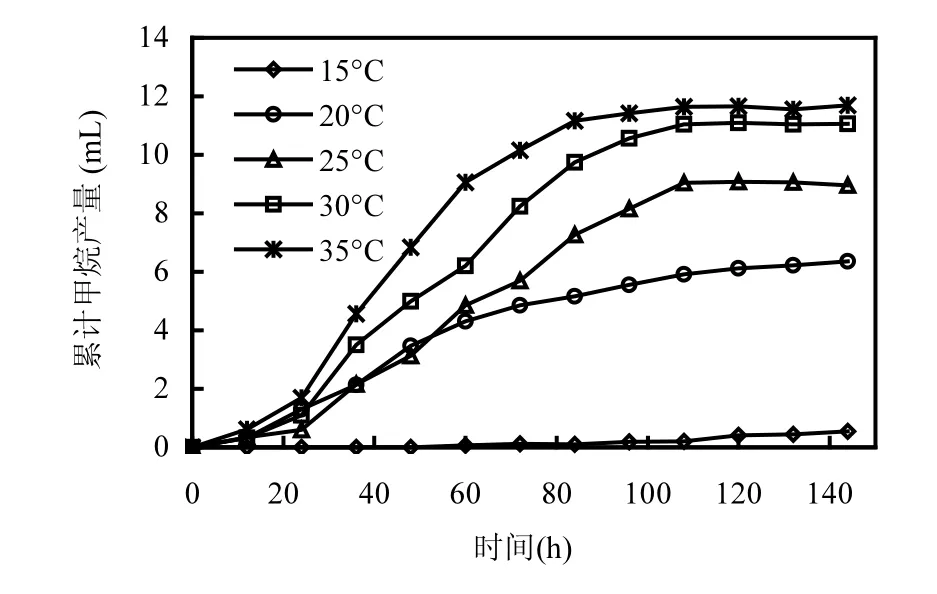
圖2 不同溫度條件下乙酸營養(yǎng)型產(chǎn)甲烷菌甲烷產(chǎn)量Fig.2 CH4 production of acetotrophic methanogens at different temperature conditions
當溫度降低至15℃時(圖2),甲烷的生成出現(xiàn)了明顯的停滯期,并且該停滯期長達96h,表明低溫對乙酸營養(yǎng)型產(chǎn)甲烷菌代謝活性造成了嚴重的抑制作用.盡管經(jīng)過了96h的停滯期,累積產(chǎn)甲烷量有所增加,但甲烷產(chǎn)量與35℃時相比,減少了95.7%.可見,低溫(15℃)對乙酸營養(yǎng)型產(chǎn)甲烷菌代謝活性造成了長期的抑制作用.與氫營養(yǎng)型產(chǎn)甲烷菌累積產(chǎn)甲烷趨勢相似,乙酸營養(yǎng)型產(chǎn)甲烷菌在35,30,25,20℃時的累積產(chǎn)甲烷量也是先線性增加,后緩慢增加(平臺期),而在15℃時出現(xiàn)了96h的停滯期,且隨后累積產(chǎn)甲烷量呈線性增加趨勢.由此可見,低溫對乙酸營養(yǎng)型產(chǎn)甲烷菌的脅迫也是通過控制半速度常數(shù)Ks來實現(xiàn)的,溫度的下降導(dǎo)致Ks迅速增加,使得乙酸營養(yǎng)型產(chǎn)甲烷菌利用乙酸的速率顯著下降,導(dǎo)致甲烷產(chǎn)量的迅速下降.盡管在15℃條件下,乙酸營養(yǎng)型產(chǎn)甲烷菌的活性一直比較低,但如果延長培養(yǎng)時間,乙酸營養(yǎng)型產(chǎn)甲烷細菌可能適應(yīng)低溫環(huán)境后仍會表現(xiàn)出一定的產(chǎn)甲烷性能.在以前的研究中,也證實了這一點[11].
結(jié)合圖1和圖2可以看出,溫度降低對2種營養(yǎng)類型的產(chǎn)甲烷菌的代謝活性都產(chǎn)生了抑制作用,這是因為任何自發(fā)反應(yīng)總是朝著吉布斯自由能減小的方向進行,而吉布斯自由能是溫度的狀態(tài)函數(shù),溫度下降會使吉布斯自由能增加,進而導(dǎo)致反應(yīng)速率的下降.從不同溫度條件下累積產(chǎn)甲烷趨勢上可以進一步看出,低溫對兩類產(chǎn)甲烷菌的脅迫是通過控制半速度常數(shù)Ks來實現(xiàn)的,溫度的下降導(dǎo)致Ks迅速增加,使得兩類產(chǎn)甲烷菌代謝速率顯著下降,導(dǎo)致甲烷產(chǎn)量的迅速下降. Lawrence等[18]研究發(fā)現(xiàn),當溫度從35℃降至20 ℃,乙酸鹽轉(zhuǎn)化為甲烷的Ks值從164mg/L增加到了2130mg/L,與本研究的結(jié)果一致.
另外,本研究還發(fā)現(xiàn)溫度降低對不同營養(yǎng)類型產(chǎn)甲烷菌代謝活性的抑制程度存在差異.在20~30℃時乙酸營養(yǎng)營養(yǎng)型產(chǎn)甲烷菌表現(xiàn)出更好的耐受性,而在15℃以下時氫營養(yǎng)型產(chǎn)甲烷菌則表現(xiàn)出更好的耐受性,這可能與溫度降低對2種營養(yǎng)類型的半速度常數(shù)Ks影響程度不同.在20~30℃時,溫度降低對乙酸產(chǎn)甲烷Ks的影響程度弱于H2/CO2產(chǎn)甲烷Ks,而在15℃以下時氫營養(yǎng)型產(chǎn)甲烷菌則表現(xiàn)出更好的耐受性.換言之,乙酸營養(yǎng)營養(yǎng)型產(chǎn)甲烷菌在低溫條件下微生物活性喪失更快.
2.3 動力學(xué)分析
如表2所示,擬合系數(shù)(R2)均大于0.98,表明該模型用來描述累計產(chǎn)甲烷量的進程是合理的. 在35℃條件下,氫營養(yǎng)型產(chǎn)甲烷菌的最大產(chǎn)甲烷速率(Rmax)為0.74mL/h,而最大甲烷產(chǎn)量(Pmax)為24.5mL.溫度從35℃分別降至30,25,20,15℃,導(dǎo)致Rmax和Pmax分別減少了58.1%~63.3%和29.5%~58.9%.當溫度≥30℃,甲烷生成沒有被延遲,但是溫度≤25℃時,甲烷生成推遲了1.7~28.7h.對于乙酸營養(yǎng)型產(chǎn)甲烷菌而言,當溫度從35℃降低至30,25,20,15℃時,Pmax和Rmax減少2.5%~65.8%和13.5%~92.6%.與對照(35℃)相比,溫度下降導(dǎo)致λ增加了0.4~7.4倍.
甲烷生成動力學(xué)的結(jié)果表明,氫營養(yǎng)型產(chǎn)甲烷菌對溫度下降具有更好的適應(yīng)性.因此,在相同溫度條件下,氫營養(yǎng)型產(chǎn)甲烷菌的甲烷生成速率高于乙酸營養(yǎng)型產(chǎn)甲烷菌.盡管在15℃條件下,2類產(chǎn)甲烷菌的甲烷生成速率都比較低,但乙酸營養(yǎng)型產(chǎn)甲烷菌表現(xiàn)出了更長的遲滯期.該結(jié)果很好解釋了在15~20℃時UASB反應(yīng)器中氫分壓不會增加而乙酸出現(xiàn)短時間積累的原因,這與H?j等[19]前期研究結(jié)果一致.
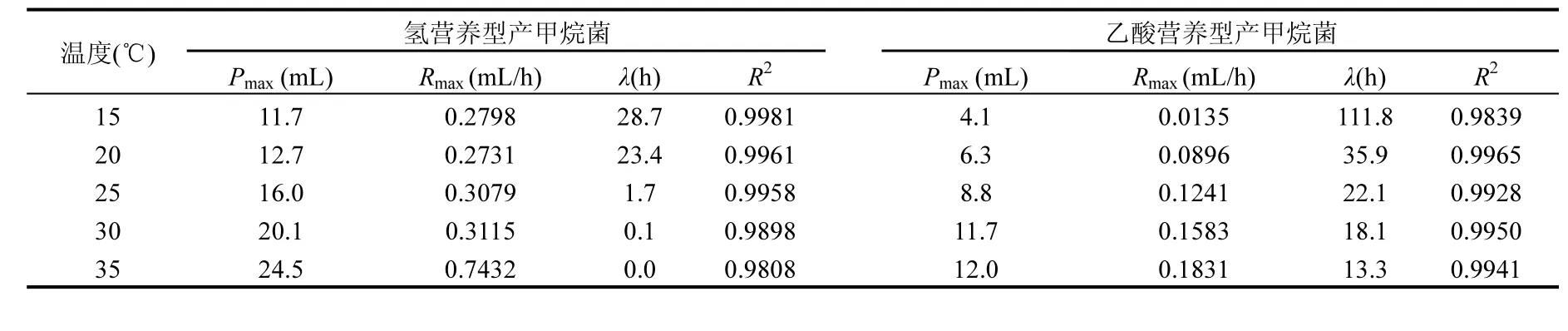
表3 不同溫度條件下產(chǎn)甲烷動力學(xué)分析Table 3 Kinetic parameters of methane production under different temperatures
3 結(jié)論
3.1 溫度下降可導(dǎo)致氫營養(yǎng)型產(chǎn)甲烷菌和乙酸營養(yǎng)型產(chǎn)甲烷菌活性受到抑制,并且溫度降低對不同營養(yǎng)類型產(chǎn)甲烷菌活性的抑制程度存在顯著差別.在20~30℃時乙酸營養(yǎng)營養(yǎng)型產(chǎn)甲烷菌對溫度脅迫表現(xiàn)出更好的耐受性,而在15℃以下時氫營養(yǎng)型產(chǎn)甲烷菌則表現(xiàn)出更好的耐受性.
3.2 溫度降低使兩類產(chǎn)甲烷菌的Pmax和λ發(fā)生了顯著變化.氫營養(yǎng)型產(chǎn)甲烷菌的Pmax由35℃時的24.5mL逐漸降低至15℃時的11.7mL,λ被延長了1.7~28.7h.而乙酸營養(yǎng)型產(chǎn)甲烷菌Pmax比35
℃條件下減少了2.5%~92.6%,λ被顯著延長,特別是在15℃時λ達到了111.8h.
參考文獻:
[1] 何 強,孫興福,艾海男,等.兩相一體式污泥濃縮消化反應(yīng)器運行效能及其微生物特性 [J]. 中國環(huán)境科學(xué), 2012,32(11):2039-2046.
[2] 張立國,李建政,班巧英,等.pH對UASB運行效能及產(chǎn)甲烷互營菌群的影響 [J]. 哈爾濱工業(yè)大學(xué)學(xué)報, 2013,45(8):44-49.
[3] 張立國,李建政,班巧英,等.升流式厭氧污泥床和連續(xù)流攪拌槽式反應(yīng)器的廢水處理效能及產(chǎn)甲烷菌群組成的對比分析 [J].科技導(dǎo)報, 2012,30(31):23-27.
[4] Liu Y, Whitman W B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea [J]. Annals of the New York Academy of Sciences, 2008,1125:171-189.
[5] 陳廣銀,鄭 正,常志州,等.不同氮源對麥稈厭氧消化過程的影響 [J]. 中國環(huán)境科學(xué), 2011,31(1):73-77.
[6] 宋佳秀,任南琪,錢東旭,等.醌呼吸影響厭氧消化產(chǎn)CO2/CH4及轉(zhuǎn)化有毒物質(zhì)的研究 [J]. 中國環(huán)境科學(xué), 2014,34(5):1236-1241.
[7] Glissmann K, Chin K J, Casper P, et al. Methanogenic pathway and archaeal community structure in the sediment of eutrophic lake dagow: effect of temperature [J]. Microbial Ecology, 2004,48:389-399.
[8] Leit?o R C, van Haandel A C, Zeeman G, et al. The effects of operational and environmental variations on anaerobic wastewater treatment system: A review [J]. Bioresource Technology, 2006, 97:1105-1118.
[9] Borja R, Bank C J. Responses of an anaerobic fluidized bed reactor treating ice-cream wastewater to organic, hydraulic, temperature and pH shocks [J]. Journal of Biotechnology, 1995, 36:251-259.
[10] El-Mashad H E M H. Solar thermophilic anaerobic reactor (STAR) for renewable energy production [D]. Ph.D. Thesis. Wageningen University, 2003.
[11] Ban Q, Li J, Zhang L, Jha A K, et al. Syntrophic propionate degradation response to temperature decrease and microbial community shift in an UASB Reactor [J]. Journal of Microbiology and Biotechnology, 2013,23:382–389.
[12] APHA. Standard methods for the examination of water and wastewater [M]. American Public Health Association (Ed). Washington, 1995.
[13] Owen W F, Stuckey D C, Healy J B, et al. Bioassay for monitoring biochemical methane potential and anaerobic toxicity [J]. Water Research, 1979,13:485-492.
[14] Chang S, Li J, Liu F. Evaluation of different pretreatment methods for preparing hydrogen-producing seed inocula from waste activated sludge [J]. Renewable Energy, 2011,36:1517-1522.
[15] Demirel B, Scherer P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass methane: a review [J]. Reviews in Environmental Science and Bio/Technology, 2008,7:173-190.
[16] Ferry J G, Smith P H, Wolfe R S. Methanospirillum, a new genus of methanogenic bacteria, and characterization of Methanospirillum hungatei sp. nov [J]. International Journal of Systematic Bacteriology, 39(1974):10-13.
[17] Zheng D, Raskin L. Quantification of Methanosaeta species in anaerobic bioreactors using genus- and species-specific hybridization probes [J]. Microbial Ecology, 2000,39:246-262.
[18] Lawrence A W, McCarty P L. Kinetics of methane fermentation in anaerobic treatment [J]. Water Pollution Control Federation. 1969, 41(2):1-17.
[19] H?j L, Olsen R A, Torsvik V L. Effects of temperature on the diversity and community structure of known methanogenic groups and other archaea in high arctic peat [J]. ISME Journal, 2008,2:37-48.
水厭氧生物處理研究.
Response of methanogens on temperature stress in an UASB reactor.
ZHANG Li-guo1, BAN Qiao-ying1*, LI Jian-zheng2(1.College of Environment and Resource, Shanxi University, Taiyuan 030006, China;2.School of Municipal and Environmental Engineering, Harbin Institute of Technology, Harbin 150090, China). China Environmental Science, 2016,36(4):1082~1086
Abstract:The effects of temperature stress on different nutrition type methanogens in an upflowed anaerobic sludge blanket (UASB) reactor was investigated by batch culture. The results showed that the dominant hydrogenotrophic and acetotrophic methanogens in this reactor were Methanospirillum and Methanosaeta, respectively. Under 35℃ condition, the accumulative methane production from hydrogenotrophic and acetotrophic methanogens were 24.7 and 11.7mL, separately. And the maximum methane production rate (Rmax) were 0.74 and 0.18mL/h, respectively. Temperature decreases, from 35℃ to 30, 25, 20, 15℃, resulted in accumulative methane productions from hydrogenotrophic methanogens were decreased by 14.2%, 34.0%, 47.0%, 57.5%, respectively. While those from acetotrophic methanogens were reduced by 5.1%, 23.9%, 45.3%, 95.7%, separately. These results indicated that hydrogenotrophic methanogens are more sensitive to temperature tress at 20~30℃, while acetotrophic methanogens are more sensitive at ≤15℃.
Key words:UASB reactor;methanogens;temperature stress;methane production
作者簡介:張立國(1980-),男,河南安陽人,講師,博士,主要從事廢
基金項目:國家自然科學(xué)基金項目(51508316);山西省基礎(chǔ)研究項目(2015021136,2015021134)
收稿日期:2015-09-18
中圖分類號:X703.5
文獻標識碼:A
文章編號:1000-6923(2016)04-1082-05
