油棕等熱帶植物DXS基因的生物信息學分析
2016-05-27 01:50:46曹紅星李東霞雷新濤
廣西植物 2016年4期
石 鵬, 曹紅星, 李東霞, 王 永, 雷新濤
( 中國熱帶農業科學院 椰子研究所 海南省熱帶油料作物生物學重點實驗室, 海南 文昌 571339 )
?
油棕等熱帶植物DXS基因的生物信息學分析
石鵬, 曹紅星, 李東霞, 王永, 雷新濤*
( 中國熱帶農業科學院 椰子研究所 海南省熱帶油料作物生物學重點實驗室, 海南 文昌 571339 )
摘要:油棕等熱帶植物含有豐富的胡蘿卜素和維生素E等類異戊二烯物質,類胡蘿卜素和甾醇等類異戊二烯物質在植物生命活動中扮演重要角色,并且對保護人類健康具有重要意義,MEP途徑是合成類異戊二烯的重要途徑之一。DXS是MEP途徑中的第一個限速酶,其功能在油棕等熱帶植物中極其保守。為了弄清油棕等熱帶植物DXS的結構和功能特點,該研究利用生物信息學工具和軟件對以油棕等熱帶植物類異戊二烯合成關鍵基因DXS為對象,進行核酸和氨基酸序列的理化性質、蛋白質結構以及功能結構域等分析,探討了不同物種間的親緣關系。結果表明:DXS基因起始密碼子均為ATG,終止密碼子則分為TAG、TAA和TGA,DXS蛋白質屬于不具有信號肽的親水性蛋白,可能作為轉運蛋白在葉綠體基質中發揮作用,未發現明顯的跨膜結構域,磷酸化位點有36個,其中絲氨酸、蘇氨酸和酪氨酸位點分別為17、11和8個,無規則卷曲和α-螺旋是蛋白質二級結構主要的結構元件,三級結構預測具有DXS酶特征,硫胺素焦磷酸鹽結合位點和PLN02582保守結構域,不同植物DXS功能結構域非常保守,可以作為判斷不同物種間親緣關系的重要依據。該研究結果為油棕等熱帶植物DXS的結構、功能分析和利用提供了進一步的信息,為其品質性狀分子機制研究及遺傳改良奠定了基礎。
關鍵詞:油棕, DXS基因, 生物信息學, 系統進化樹
油棕(ElaeisguineensisJacq.)屬棕櫚科多年生喬木,是熱帶產油植物,果實中類胡蘿卜素的含量是決定油棕品質的一個重要指標。粗棕櫚油中含有600 mg·kg-1類胡蘿卜素,其中80%都是β-胡蘿卜素和α-胡蘿卜素,這也是粗棕櫚油呈黃色或紅棕色的原因(雷新濤和曹紅星,2013)。巴西橡膠、可可、白木香、杜仲和麻瘋樹都是重要的熱帶植物,對油棕等熱帶植物進行DXS的生物信息學分析,對于了解這些重要植物的DXS蛋白質結構和功能具有重要意義,同時也為分析這些熱帶植物間進化關系提供依據。
類異戊二烯是最大的一類天然產物,目前為止已知的有3萬多種(Sacchettini & Poulter,1997)。一些類異戊二烯在細胞中有著重要作用,如甾醇是細胞膜結構的重要組分(Erick,2008),長萜醇在蛋白質糖基化中擔任糖基載體(Vincent & Dirk,2011),泛素和質體醌在光合作用和呼吸作用中扮演電子轉運體的角色(Keisuke et al,2010),脫落酸、細胞分裂素、赤霉素和甾醇作為激素控制著生理和發育過程(Jonathan & Neil,2001)。盡管各種類異戊二烯的結構和功能存在差異,但是所有的都由異戊烯焦磷酸(IPP)和它的同分異構體二甲基丙烯焦磷酸(DMAPP)構成。IPP和DMAPP能通過兩個不同生物合成路線:甲羥戊酸(MVA)途徑,其主要存在真菌、酵母和動物細胞質中;2-C-甲基-D-赤藻糖醇-4-磷酸(MEP)途徑,其存在原生生物,大多數細菌,綠藻和高等植物質體中(Lombard & Moreira,2011;Wanke et al,2001;Vranova et al,2013)。MEP途徑的第一步是丙酮酸和3-磷酸甘油醛在1-脫氧木酮糖-5-磷酸合成酶(DXS)的催化下生成1-脫氧木酮糖-5-磷酸(DXP)。DXS是一種新的轉酮酶,由DXS基因翻譯而來(Elizabeth et al,2009;Han et al,2013)。在植物和大腸桿菌中,由此反應產生的DXP被用來在質體中生成IPP。DXS基因最初在大腸桿菌中被克隆和分析,其它細菌和高等植物中隨后也找到了同源基因(Cordoba et al,2011;Zhang et al,2009;Gong et al,2006;Luisa et al,1998;Kuzuyama et al,2000;Hahn et al,2001)。胡蘿卜素和葉黃素統稱為類胡蘿卜素,是所有進行光合作用生物中的色素。在高等植物中,類胡蘿卜素在綠色組織光合作用中扮演重要角色。其中,β-胡蘿卜素是維生素A的前體,具有抗癌的作用,其缺乏會導致失明和早逝(Rao et al,2007;Shan & Li,2008;Aguila & Manuel,2012)。由于類胡蘿卜素的重要性,高等植物中編碼類胡蘿卜素生物合成的酶基因成為了增加類胡蘿卜素生物合成的潛在位點。有文獻報道證實DXS是途徑中的限速酶,MEP途徑中的基因高表達與植物中更高含量的類胡蘿卜素有直接關系(Peng,2008)。
生物信息學興起于21世紀,利用信息學工具分析核酸和蛋白質等大分子,大規模挖掘這些大分子的生物學意義(Luscombe et al,2001)。目前,植物中開展DXS的生物信息學分析還不多,尤其在油棕等熱帶植物中還很少報道。本文主要以油棕等熱帶植物的DXS核酸和蛋白質序列為分析對象,利用大量的生物信息學工具挖掘其序列的生物學意義,為進一步研究油棕等熱帶植物DXS基因功能和類胡蘿卜素代謝過程奠定基礎。
1材料與方法
1.1 數據來源
核酸和蛋白質序列均來源于NCBI(National Center for Biotechnology Information)數據庫中已有的油棕等熱帶植物1-脫氧木酮糖-5-磷酸合成酶基因的核酸序列及其對應的蛋白質序列,如油棕(ElaeisguineensisJacq.,AY583783、AAS99588)、白木香(Aquilariasinensis,JX860326、AFU75321)、杜仲(Eucommiaulmoides,JX458817、AFU93068)、巴西橡膠(Heveabrasiliensis,AB294698、BAF98288)、麻瘋樹(Jatrophacurcas,KF955327、AHL17583)、可可(Theobromacacao,XM007035370、XP007035432);另外,構建系統進化樹還用到了以下幾種植物的DXS蛋白質序列:高良姜(Alpiniaofficinarum,AEK69518)、陽春砂(Amomumvillosum,ACR02668)、擬南芥(Arabidopsisthaliana,NP193291)、巴豆(Crotonstellatopilosus,BAF75640)、海島棉(Gossypiumbarbadense,ABN13970)、苜蓿(Medicagotruncatula,XP003609567)、煙草(Nicotianatabacum,ACF60511)、粳稻(Oryzasativa,NP001055524)、葛麻姆(Puerariamontana,AAQ84169)、蓖麻(Ricinuscommunis,XP002516843)、羅漢果(Siraitiagrosvenorii,AEM42997)、番茄(Solanumlycopersicum,NP001234672)、馬鈴薯(S.tuberosum,NP001275130)、南非醉茄(Withaniasomnifera,AFI98878)、玉米(Zeamays,NP001157805)。

表 1 生物信息學分析使用的工具和軟件
1.2 方法
所用生物信息學工具和軟件如表1所示,具體參數設置見結果。
2結果與分析
2.1 油棕等熱帶植物DXS基因核酸及氨基酸序列分析
利用ORF Finder和ProtParam對油棕、巴西橡膠、麻瘋樹、杜仲、可可和白木香6種熱帶植物DXS基因的核酸序列及其氨基酸序列進行分析。分析結果如表2所示,不同植物DXS基因開放讀碼框的堿基數及所編碼的氨基酸殘基數變化不大,起始密碼子均為ATG,終止密碼子分為3種,即油棕、巴西橡膠和白木香均為TAG,麻瘋樹和杜仲為TAA,可可則為TGA ;而且不同植物DXS的分子量、理論等電點和高含量氨基酸成分基本一致; Leu、Ala、Gly是這6種熱帶植物DXS中含量最豐富的氨基酸;根據不穩定性指數小于40為穩定性蛋白質判斷,巴西橡膠、杜仲和可可的DXS為穩定類蛋白質,而油棕、麻瘋樹和白木香的DXS為不穩定類蛋白質;且它們的親水系數較低,均為親水性蛋白質。

表 2 6種熱帶植物DXS核酸及氨基酸序列分析
2.2 油棕等熱帶植物DXS親水性/疏水性的預測和分析
利用ProtScale對油棕氨基酸序列的疏水性/親水性進行預測(圖1)。從圖1發現,多肽鏈第55位的His具有最低的分值-2.656,親水性最強;而第185位的Gly具有最高的分值2.067,疏水性最強。整體來看,親水氨基酸分布比較均勻,且數量大于疏水氨基酸,故推測油棕DXS蛋白質為親水性蛋白。
2.3 油棕等熱帶植物DXS二級結構的預測和分析
利用SOPMA對油棕DXS氨基酸序列進行二級結構預測(圖2)。從圖2可以發現,油棕DXS由40.45%的無規則卷曲(Random coil),38.19%的α-螺旋(Alpha helix),15.28%的延伸鏈(Extended strand)和6.08%β-轉角(Beta turn)組成。在整個蛋白質結構中,無規則卷曲和α-螺旋是油棕DXS最主要的結構元件,而延伸鏈和β-轉角則次之。
2.4 油棕等熱帶植物DXS蛋白三級結構的預測和分析
利用SWISS-MODEL的同源建模方法預測油棕DXS蛋白的三級結構,獲得油棕DXS三維結構模式(圖3)。同源建模分析發現其與蛋白質模板2o1s.1.A有50.58%的相似性,且根據三級結構特征推測為1-脫氧木酮糖-5-磷酸合成酶(1-deoxy-D-xylulose-5-phosphate synthase)。從圖3可以看出,紫色卷曲狀結構代表的無規則卷曲遍布在油棕DXS三級結構中。
2.5 油棕等熱帶植物DXS跨膜結構域的預測和分析
利用TMHMM對不同熱帶植物DXS氨基酸序列的跨膜結構域進行預測(圖4),未發現明顯的跨膜結構域。
2.6 油棕等熱帶植物DXS翻譯后修飾的預測和分析
利用NetPhos 2.0 Server對油棕DXS的翻譯后修飾預測的結果表明(圖5),整個多肽鏈中分值在0.5(閾值)以上的氨基酸位點有36個,可知油棕DXS的磷酸化位點有36個,其中Ser:17,Thr:11,Tyr:8。
2.7 油棕等熱帶植物DXS的亞細胞定位分析
利用PSORT對油棕等熱帶植物的DXS基因的編碼產物進行亞細胞定位分析,結果表明,油棕和其它熱帶植物的DXS蛋白質定位在葉綠體中基質的可能性最大,該結果說明熱帶植物DXS蛋白可能主要是在葉綠體基質中行使功能。
2.8 油棕等熱帶植物DXS信號肽和導肽的預測及分析
利用SignalP 4.1 Server對油棕DXS氨基酸序列的信號肽進行預測(圖6)。圖6結果表明,油棕DXS不含有信號肽。
同時利用TargetP1.1 Server進行導肽分析也證實了其作為葉綠體轉運蛋白(分值為0.409)的可能性比較大(圖7),該蛋白存在信號肽(分值為0.030)的可能性很小。此結果與亞細胞定位及信號肽預測結果基本一致。
2.9 油棕等熱帶植物DXS保守結構域的預測和分析
通過NCBI中的保守結構域數據庫 (Conserved
Domain Database,CDD)工具對DXS基因的氨基酸序列分析發現,DXS含有硫胺素焦磷酸鹽(TPP,Thiamine pyrophosphate)和嘧啶(PYR,Pyrimidine)結合位點,TPP_DXS、TPP_PYR_DXS_TK_like和Transketolase_C保守域,TPP_enzymes、TPP_enzyme_PYR和Transketolase_C superfamily家族和PLN02582保守結構域。其中PLN02582是1-脫氧木酮糖-5-磷酸合成酶的特征序列(圖8)。

圖 1 油棕DXS的疏水性/親水性預測和分析Fig. 1 Hydrophobicity or hydrophilicity prediction and analysis of EgDXS
2.10 油棕等熱帶植物DXS序列比對及系統進化分析
用Blast程序比對油棕與其它植物DXS的核酸及氨基酸序列同源性,結果表明油棕DXS的核酸序列與其它植物如高良姜(HQ874656)、麻瘋樹(KF955327)、葛麻姆(FJ455512)、橡膠(AB294698)、番茄(NM001247743)和南非醉茄(JQ710678)的一致性分別是80%、80%、79%、79%、
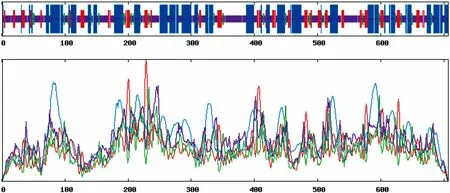
圖 2 油棕DXS二級結構預測 藍色線條,紅色線條,綠色線條和紫色線條分別表示α-螺旋,延伸鏈,β-轉角和無規則卷曲。橫坐標和縱坐標分別表示二級結構在所處的位置和數量。Fig. 2 Secondary structure prediction of EgDXS Blue lines, red lines, green lines, purple lines represent alpha helix, extended strand, beta turn and random coil respectively. Horizontal and vertical axis represent position and quantity of secondary structure respectively.
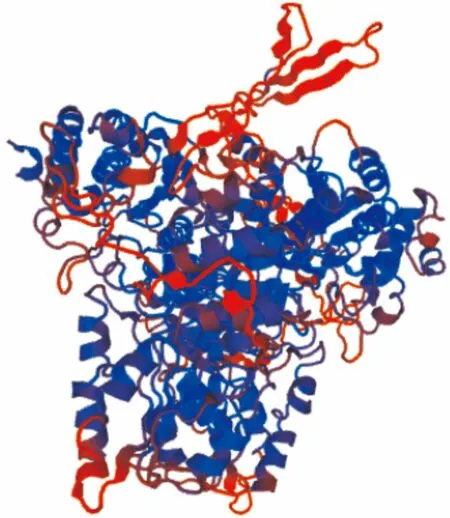
圖 3 油棕DXS三級結構預測Fig. 3 Tertiary structure prediction of EgDXS

圖 4 油棕DXS的跨膜結構域預測Fig. 4 Transmembrane structure prediction of EgDXS
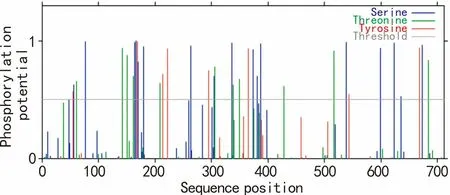
圖 5 油棕DXS翻譯后修飾預測 磷酸化位點分為絲氨酸,蘇氨酸和酪氨酸3種。Fig. 5 Posttranslational modification prediction of EgDXSPhosphorylation sites are divided into serine, threonine and tyrosine.
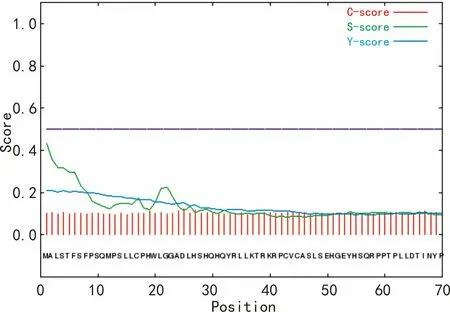
圖 6 油棕DXS信號肽預測 C-score. 切割位點的可能性; S-score. 氨基酸是信號肽的可能性; Y-score. 結合C-score和S-score的導數,能更好地反映切割位點的可能性。Fig. 6 Signal peptide prediction of EgDXS C-score. Possibility of cleavage site; S-score. Possibility of signal peptide; Y-score. Derivative of the C-score combined with the S-score resulting in a better cleavage site prediction than the raw C-score alone.

圖 7 油棕DXS導肽預測 Len. 蛋白質長度; cTP. 作為葉綠體轉運蛋白的可能性; mTP. 作為線粒體轉運蛋白的可能性; SP. 信號肽的可能性。Fig. 7 Leading peptide prediction of EgDXS Len. Length of protein; cTP. Possibility of chloroplast transport protein; mTP. Possibility of mitochondira transport protein; SP. Possibility of signal pepetide.
79%和79%;蛋白質的Blastp比對發現,油棕DXS的氨基酸序列與其他植物的氨基酸序列有更高的相似性,如與長春花(AGL40532)、陽春砂(ACR02668)、玉米(ACG27905)、麻瘋樹(AHL17583)、高良姜(AEK69518)和羅漢果(AEM42997)的一致性分別為86%、85%、86%、86%、86%和85%。以上結果表明,植物DXS氨基酸序列之間的相似性高于核酸序列。
把21個來自不同植物的DXS氨基酸序列用MEGA 6.06軟件采用Neighbor-Joining法,進行1 000次bootstrap統計學檢驗構建系統進化樹(圖9)。圖9結果表明,21種不同植物的DXS氨基酸序列可以分為3大類,巴西橡膠和蓖麻等10種植物分為第Ⅰ類,其中大戟科植物麻瘋樹、巴豆、蓖麻和巴西橡膠在此類,而白木香、可可和杜仲等熱帶植物也聚在此類;豆科植物和茄科植物分在第Ⅱ類,其中同為豆科植物的葛麻姆和苜蓿聚成一簇的信心指數為100%,馬鈴薯、番茄、南非醉茄和煙草為茄科,其中馬鈴薯和番茄聚成一簇的信心指數為100%;油棕、姜科和禾本科植物分在第Ⅲ類,其中同為禾本科的粳稻和玉米聚成一簇的信心指數為100%,同為姜科的高良姜和陽春砂聚成一簇的信心指數為100%。從聚類分析結果可以發現,用DXS氨基酸序列構建的系統進化樹可以比較真實地反映不同植物之間的進化關系,對判斷植物間的親緣關系具有很好的參考價值。
3討論與結論
類胡蘿卜素對于植物來說是必需的,并且在為人類提供營養和保護健康方面具有重要作用。本文利用生物信息學工具和軟件對油棕等熱帶植物的DXS基因的核酸及蛋白質理化性質、結構組成和功能特征等進行了預測和分析,并構建了油棕等21種植物的系統進化樹。預測結果表明,不同植物DXS的理化性質差異不大,蛋白質穩定性不一,都屬于親水性蛋白質。跨膜結構域、信號肽和導肽預測結果表明,油棕等熱帶植物的DXS都沒有發現明顯的跨膜結構域和信號肽,主要是在葉綠體基質中發揮作用,可能作為葉綠體轉運蛋白行使功能。DXS的保守結構域分析發現,DXS含有硫胺素焦磷酸鹽和嘧啶結合位點, TPP_DXS、TPP_PYR_DXS_TK_like和Transketolase_C保守域,TPP_enzymes、TPP_enzyme_PYR和Transketolase_C superfamily家族以及PLN02582保守結構域,其中PLN02582保守結構域是1-脫氧木酮糖-5-磷酸合成酶的特征結構域。
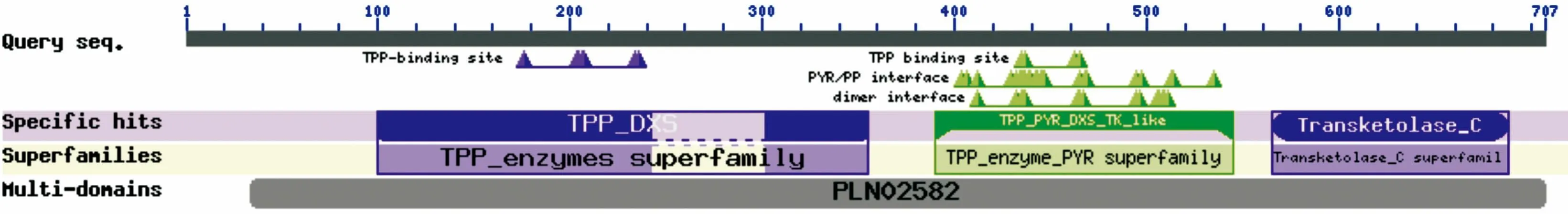
圖 8 油棕DXS保守結構域預測Fig. 8 Conserved domain prediction of EgDXS
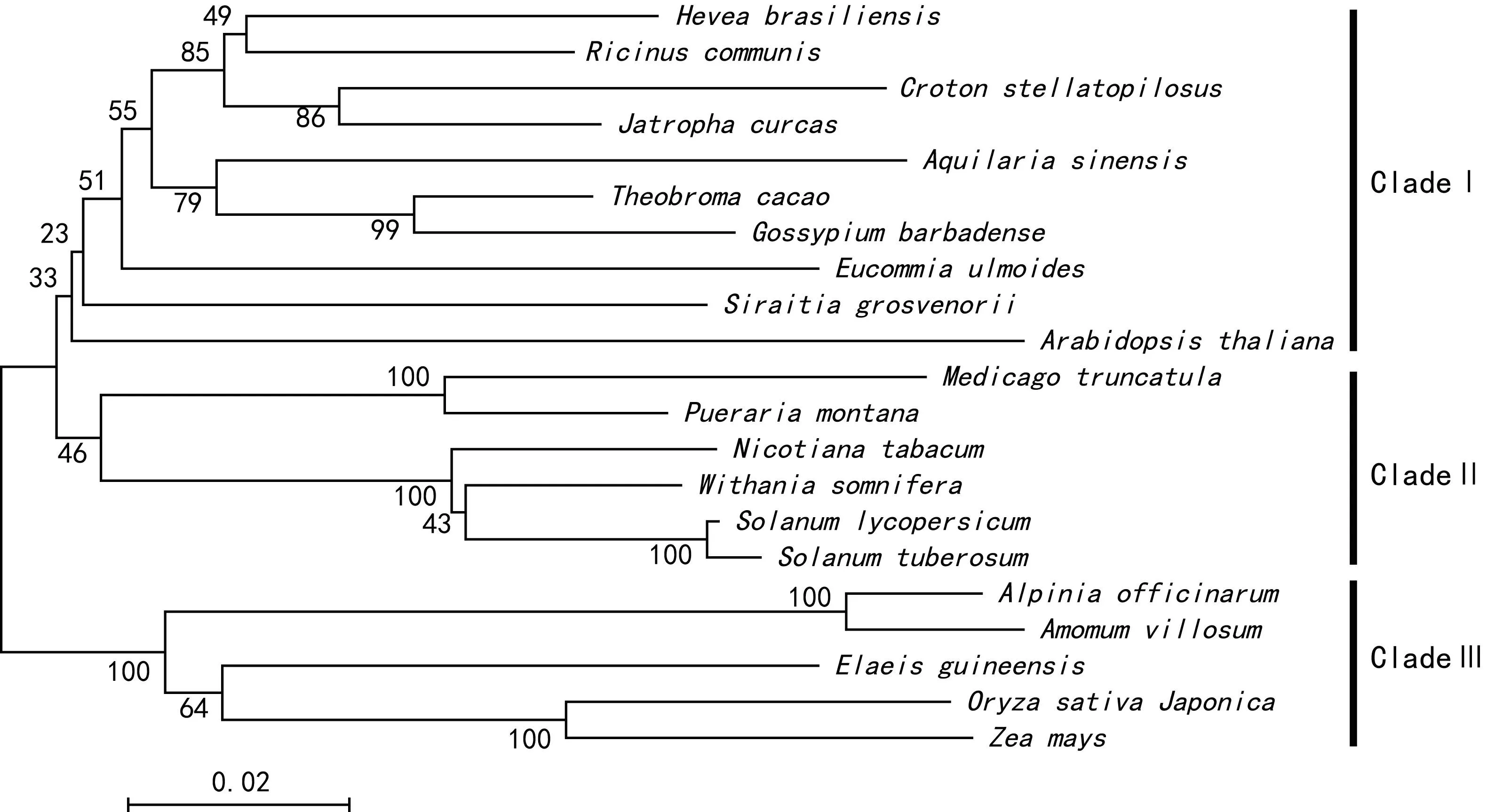
圖 9 油棕等21種植物DXS氨基酸序列系統進化樹分析Fig. 9 Phylogenetic tree analysis of Amino acid series from 21 plants including oil palm
在MEP途徑中,以丙酮酸和3-磷酸甘油醛為底物,先后經過8個酶的催化最終形成了合成類異戊二烯的通用底物IPP和DMAPP。實驗證實作為MEP途徑第一個酶DXS在IPP和DMAPP合成過程中發揮重要作用。擬南芥中對MEP途徑中的酶研究較多,目前MEP途徑中的所有酶都已經確認。MEP途徑的第一個反應就是在DXP合成酶DXS的催化作用下甘油醛-3-磷酸硫胺素縮合生成1-脫氧木酮糖-5-磷酸。研究表明,通過MEP途徑合成類胡蘿卜素主要受到限速酶DXS,DXR和HDR的控制(Sawitri & Wallie,2005;Elizabeth et al,2009)。Paruicia et al(2006)認為在充分了解植物類胡蘿卜素合成機制的基礎上,可以通過生物技術手段提高植物中的類胡蘿卜素,為人類提供類胡蘿卜素含量更高的食物。目前,番茄、馬鈴薯、油菜和水稻中都已經開展了相關轉基因實驗。植物中類胡蘿卜素的代謝和生物合成途徑已經得到廣泛研究,合成途徑中幾乎所有編碼合成酶的基因都被分離出來。Sanae & Akemi(2006)研究菊花花瓣顏色發現,是類胡蘿卜素的合成和積累機制不同形成了黃色和白色兩種花瓣顏色,其中LCYE,LCYB和DXS的表達量與花瓣中類胡蘿卜素的含量息息相關。在番茄果實發育過程中,DXS的表達量與果實中類胡蘿卜素含量呈正相關關系,說明DXS在番茄果實發育和器官形成過程中發揮重要作用(Luisa et al,2000)。萊茵衣藻中研究發現,DXS和DXR等基因的表達量隨著光周期波動,DXS等基因的表達量表現出典型的晝夜節律模式,并且也會受到中間代謝產物的反饋調節(Tian et al,2010)。因此,在植物類胡蘿卜素合成代謝過程中,DXS是調控類胡蘿卜素合成的關鍵酶,對其進行生物信息學分析為今后深入研究具有重要意義。油棕和巴西橡膠等熱帶植物DXS生物信息學分析,可以尋找這些不同科的熱帶植物在DXS上的共同特征和不同點,希望為熱帶植物DXS結構和功能研究奠定基礎。植物激素、類胡蘿卜素和甾醇等類異戊二烯物質在熱帶植物生命活動中具有重要作用,而它們在熱帶植物中的具體功能作用及其控制基因研究較少,需要進一步開展相關研究,揭開類異戊二烯在熱帶植物中的生物合成途徑及其調控機制。油棕等21種植物DXS氨基酸序列比對和系統進化樹分析發現,不同植物的DXS序列相似性較高,功能域非常保守,以DXS氨基酸序列構建的進化樹能準確反映不同植物間的親緣關系,也可作為評價油棕等熱帶植物種質資源的重要依據。
參考文獻:
AGUILA RS, MANUEL RC, 2012. Carotenoid biosynthesis inArabidopsis: a colorful pathway [J]. Arabidopsis Book, 10:e0158.CORDOBA E, PORTA H, ARROYO A, et al, 2011. Functional characterization of the three genes encoding 1-deoxy-D-xylulose 5-phosphate synthase in maize [J]. J Exp Bot, 62(6):2 023-2 038.
ERICK JD, 2008. Sterols and membrane dynamics [J]. J Chem Biol, 1(1-4):63-77.
ELIZABETH C, MARI S, PATRICIA L, 2009. Unravelling the regulatory mechanisms that modulate the MEP pathway in higher plants [J]. J Exp Bot, 60(10):2 933-2 943.
GONG YF, LIAO ZH, GUO BH, et al, 2006. Molecular cloning and expression profile analysis ofGinkgobilobaDXS gene encoding 1-deoxy-D-xylulose 5-phosphate synthase, the first committed enzyme of the 2-C-methyl-D-erythritol 4-phosphate pathway [J]. Plant Med, 72(4):329-335.
HAN M, HEPPEL SC, SU T, et al, 2013. Enzyme inhibitor studies reveal complex control of meghyl-D-erythritolf-4-phosphate (MEP) pathway enzyme expression inCatharanthusroseu[J]. PLoS ONE, 8(5):e62467.
HAHN FM, EUBANKS LM,TESTA CA, et al, 2001. 1-deoxy-D-xylulose-5-phosphate synthase, the gene product of open reading frame (ORF) 2816 and ORF 2895 inRhodobactercapsulatus[J]. J Bact, 183:1-11.JONATHAN DB, NEIL WP, 2001. Plant hormones and the control of physiological processes [J]. New Phytol, 152(3):375-407.
KEISUKE Y, MASARU S, ICHIRO T,et al, 2010. Simultaneous determination ofinvivoplastoquinone and ubiquinone redox states by HPLC-based analysis [J]. Plant Cell Physiol, 51(5):836-841.
KUZUYAMA T, TAKAGI M, TAKAHASHI S, et al, 2000. Cloning and characterization of 1-deoxy-D-xylulose-5-phosphate synthase fromStreptomycessp. Strain CL190, which uses both the mevalonate and nonmevalonate pathway for isopentenyl diphoshate biosynthesis [J]. J Bacteriol, 182(4):891-897.
LEI XT,CAO HX, 2013. Oil Palm [M]. Beijing: China Agriculture Press:103-104. [雷新濤,曹紅星, 2013. 油棕 [M]. 北京: 中國農業出版社:103-104.]
LOMBARD J, MOREIRA D, 2011. Origins and early evolution of the mevalonate pathway of isoprenoid biosynthesis in the three domains of life [J]. Mol Biol Evol, 28(1):87-99.
LUISA ML, NARCISO C, SURYA RP, et al, 1998. Cloning and characterization of a gene fromEscherichiacoliencoding a transketolase-like enzyme that catalyzes the synthesis of D-1-deoxyxylulose-5-phosphate, a common precuisor for isoprenoid, thiamin, and pyridoxol biosynthesis [J]. Proc Natl Acad Sci, 95(5):2 105-2 110.
LUSCOMBE NM, GREENBAUM D, GERSTEIN M, et al, 2001. What is bioinformatics? A proposed definition and overview of the field [J]. Meth Inf Med, 40(4):346-358.
LUISA ML, MANUEL RC, FRANCESCA G,et al, 2000. Carotenoid biosynthesis during tomato fruit development: regulatory role of 1-deoxy-D-sylulose 5-phosphate synthase [J]. Plant J, 22(6):503-513.
PENG HC, 2008. Bioimage informatics: a new area of engineering biology [J]. Bioinformatics, 24(17):1 827-1 836.
PATRICIA BP, MANUEL RC, 2006. Carotenoid biotechnology in plants for nutritionally improved foods [J]. Physiol Plant, 126(3):369-381.
RAO AV, RAO LG, 2007. Carotenoids and human health [J]. Pharmacol Res, 55:207-216.
SACCHETTINI JC, POULTER CD. 1997. Creating isoprenoid diversity [J]. Science, 277:1 788-1 789.
SANAE K, AKEMI O, 2006. Regulation of carotenoid biosynthesis in petals and leaves of chrysanthemum (Chrysanthemummorifolium) [J]. Physiol Plant, 128(3):436-447.SAWITRI K, WALLIE S, 2005. Molecular cloning and expression of a cDNA encoding 1-deoxy-D-xylulose-5-phosphate synthase from oil palmElaeisguineensisJacq. [J]. Plant Sci,169:571-578.
SHAN L, LI L, 2008. Carotenoid metabolism: biosynthesis, regulation, and beyond [J]. J Integr Plant Biol, 50(7):778-785.
TIAN HS, CHENG QL, YUAN YH, et al, 2010. Coordinated regulation of gene expression for carotenoid metabolism inChlamydomonasreinhardtii[J]. J Integr Plant Biol, 52(10):868-878.
VINCENT C, DIRK JL, 2011. From glycosylation disorders to dolichol biosynthesis defects: a new class of metabolic diseases [J]. J Inherit Metab Dis, 34(4):859-867.
VRANOVA E, COMAN D, GRUISSEM W, 2013. Network analysis of the MVA and MEP pathway for isoprenoid synthesis [J]. Ann Rev Plant Biol,64:665-700.
WANKE M, SKORUPINSKA TK, SWIEZEWSKA E, 2001. Isoprenoid biosynthesis via 1-deoxy-D-xylulose-5-phosphate/2-C-methyl-D-erythritol-4-phosphate (DOXP/MEP) pathway [J]. Acta Biochim Pol, 48(3):663-672.
ZHANG M, LI K, ZHANG C, et al, 2009. Identification and characterization of class 1 DXS gene encoding 1-deoxy-D-xylulose-5-phosphate synthase, the first committed enzyme of the MEP pathway from soybean [J]. Mol Biol Rep, 36(5):879-887.
Bioinformatics analysis ofDXSgene from six tropical plants including oil palm (ElaeisguineensisJacq.)
SHI Peng, CAO Hong-Xing, LI Dong-Xia, WANG Yong, LEI Xin-Tao*
(HainanKeyBiologicalLaboratoryofTropicalOilCrops,CoconutResearchInstitute,ChineseAcademyofTropicalAgriculturalSciences, Wenchang 571339, China )
Abstract:The paper unraveled structure characteristics, function features and phylogenetic relationships among species through analyzing nucleic acid and protein sequences of key gene DXS controlling isoprenoids synthesis of tropical plants as oil palm (Elaeis guineensis Jacq.) by using bioinformatics tools. Isoprenoids like carotenoids and sterols play an important role in life of plant, and have important significance for the protection of human health. Tropical plants as oil palm contains abundant isoprenoids like carotenes and vitamin E, MEP is one of the important pathway for isoprenoids biosynthesis. DXS as the first rate limiting enzyme in MEP pathway, its functions were highly conserved in diverse tropical plants. In order to understand the structure and function characteristics of DXS from tropical plants as oil palm, DXS genes nucleic acid and protein sequences from different plants including oil palm were analyzed such as physicochemical property, protein structure, and conserved domain by the tools of bioinformatics in this study. The results showed that the start codon of DXS gene was ATG, but termination codon was divided into TAG, TAA and TGA; DXS protein belonged to a hydrophilic protein without a signal peptide; It might played a role in the chloroplast stroma as a transport protein; No transmembrane domain was found, there were 36 phosphorylation sites, including 17 serine, 11 threonine and 8 tyrosine sites respectively; Random coil and alpha helix were the major structural elements of secondary structure; It had DXS enzyme character, thiamin diphosphate (TPP) binding sites and PLN02582 conserved multi-domains with tertiary structure analysis; Domains of DXS protein from different plant, were very conservative, and could be used as an important basis for determining genetic relationships among species. The results would provide further information for structure and functional analysis of DXS, and lay the foundation for molecular mechanism research and genetic improvement of quality traits for tropical plants including oil palm.
Key words:oil palm (Elaeis guineensis Jacq.), DXS gene, bioinformatics, phylogenetic tree
中圖分類號:Q811.4
文獻標識碼:A
文章編號:1000-3142(2016)04-0471-08
作者簡介:石鵬(1987-),男,湖南安鄉人,碩士,研究實習員,主要從事油棕種質資源評價等研究,(E-mail)624811593@qq.com。*通訊作者: 雷新濤,博士,研究員,主要從事熱帶油料作物遺傳育種等研究,(E-mail)xtlei@263.com。
基金項目:國家林業局“948”項目(No.2013-4-40)[Supported by the Plan of Introducing the Advanced Technology for Agriculture of State Forestry Administration(No.2013-4-40)]。
*收稿日期:2014-12-19修回日期: 2015-02-15
DOI:10.11931/guihaia.gxzw201406013
石鵬,曹紅星,李東霞,等. 油棕等熱帶植物DXS基因的生物信息學分析 [J]. 廣西植物, 2016, 36(4):471-478
SHI P,CAO HX,LI DX,et al. Bioinformatics analysis ofDXSgene from six tropical plants including oil palm (ElaeisguineensisJacq.)[J]. Guihaia, 2016, 36(4):471-478
