杧果乙烯抑制劑的篩選及其采后保鮮效果研究
2016-05-30 15:46:26宋康華賈志偉谷會常金梅張魯斌
熱帶作物學(xué)報 2016年9期
宋康華 賈志偉 谷會 常金梅 張魯斌
摘 要 比較研究不同濃度乙烯抑制劑AOA(aminooxyacetic acid)、AIB(α-amino-isobutyric acid)、AVG (Aminoethoxyvinyl glycine hydrochloride)、ATA(3-amino-1,2,4-triazole)、1-MCP(1-methylcyclopropene)、精胺、亞精胺對采后杧果(Mangifera indica L.)乙烯釋放的抑制效果,以期篩出最佳乙烯抑制劑并探究其對杧果的貯藏保鮮效果。結(jié)果表明:不同濃度乙烯抑制劑處理杧果,均可降低杧果果實乙烯釋放量,其中1 mmol/L精胺、10 mmol/L亞精胺和50 mmol/L ATA可有效延緩硬度、可滴定酸(Titratable acid, TA)下降,維持較高VC(Vitamin C)和TSS(Total soluble solids)含量。綜合分析,精胺、亞精胺、ATA處理可有效延緩采后杧果果實軟化,為采后杧果貯藏保鮮提供技術(shù)支持。
關(guān)鍵詞 杧果;乙烯抑制劑;乙烯釋放;保鮮效果
中圖分類號 TS255.4 文獻標(biāo)識碼 A
杧果(Mangifera indica L.)以其色、香、味俱佳以及豐富的營養(yǎng)價值而享有“熱帶果王”之美譽,是世界第二大熱帶水果[1]。在世界熱帶水果產(chǎn)量中僅此于香蕉,中國杧果種植地只限于海南、廣東、廣西、云南、福建、臺灣等省的部分地區(qū),人均產(chǎn)量不到1 kg[2],且杧果采后生理代謝旺盛,為典型的呼吸躍變型果實,對乙烯非常敏感,采收后隨著果實后熟,耐藏性和抗病性急劇下降,在極短時間內(nèi)喪失商品價值,每年中國杧果的采后損失率高達20%~30%[3]。乙烯是引發(fā)躍變型果實后熟和植物組織衰老的主要因素[4],杧果的呼吸躍變可由體內(nèi)系統(tǒng)Ⅰ乙烯觸發(fā),抑制杧果體內(nèi)系統(tǒng)Ⅰ乙烯的產(chǎn)生或作用成為延長杧果貯藏壽命的有效途徑[5]。
乙烯對果實成熟的重要作用可使人們通過控制乙烯的生物合成及降低果實對乙烯作用的敏感性而實現(xiàn)對果實成熟衰老的控制,以提高果實的貯藏性并改善其品質(zhì)[6]。乙烯抑制劑主要有乙烯生物合成抑制劑和乙烯信號轉(zhuǎn)導(dǎo)抑制劑2種類型,其作用效果隨受物種、品種、采收成熟度、處理條件等因素影響而不盡相同,因而要確定不同種類果品最適宜的處理參數(shù),以使藥品達到最優(yōu)效果。筆者以杧果栽培種‘紅杧6號及‘臺農(nóng)1號為材料,測定不同濃度乙烯作用抑制劑處理對采后杧果乙烯釋放量及果實品質(zhì)的影響,為采用乙烯抑制劑延緩杧果后熟衰老及其調(diào)控機制奠定基礎(chǔ)。
1 材料與方法
1.1 材料
供試杧果品種‘紅杧6號和‘臺農(nóng)1號果實于2015年5月采自廣東省雷州市杧果基地,采收成熟度約七成熟的果實,立即運回實驗室,分別進行以下處理:挑取大小均勻一致,無病蟲傷的果實用500 mg/L施保克進行清洗,晾干后分成22組,用以下濃度的溶液浸泡果實5 min,AOA(2、4、8 mmol/L)、AVG(0.05、0.1、0.3 mmol/L)、AIB (2、5、10 mmol/L)、ATA(10、50、100 mmol/L)、精胺(1、5、10 mmol/L)亞精胺(1、5、10 mmol/L),用不同濃度的1-MCP(0.5、1、1.5 μL/L)密閉熏蒸12 h,對照果實用清水浸泡5 min。處理完畢后晾干分裝于塑料筐中,每框10個果,套上厚度約0.02 mm厚的聚乙烯塑料薄膜袋中,置于25 ℃條件下貯藏,定期觀察杧果果實表觀變化及測定其乙烯釋放量,并取樣以備各項生理指標(biāo)的測定。本次試驗中每個處理設(shè)3個重復(fù),每個重復(fù)30個果,每個處理共90個果。
1.2 方法
1.2.1 乙烯釋放率測定 每天隨機取2個果實,在室溫下密封2 h后,抽取氣體測定乙烯釋放量,參照張魯斌等[18]方法進行。用Agilent7890A型氣相色譜儀測定乙烯釋放量。色譜柱:HP-55%(毛細管柱)。氣相色譜工作條件為:火焰離子化檢測器(FID),柱溫90 ℃,進樣器溫度100 ℃,載氣為N2,流速25 mL/min,保留時間為3.5 min。以鮮果測定,試驗重復(fù)3次。
1.2.2 品質(zhì)及生理指標(biāo)的測定 篩選對‘臺農(nóng)1號最有效乙烯抑制劑處理,并對其進行品質(zhì)相關(guān)指標(biāo)測定。可溶性固形物(TSS)含量采用VBR50手持折光儀測定[7];果實硬度利用GY-1型硬度計測定[8];可滴定酸(TA)含量采用酸堿滴定法測定,以檸檬酸計[9];維生素C(Vc)含量采用2,6-二氯酚靛酚滴定法[10]測定。
1.3 統(tǒng)計分析
試驗數(shù)據(jù)采用Excel軟件進行統(tǒng)計分析,所得數(shù)據(jù)為3次平均值,用SPSS 11.5軟件進行差異顯著性分析。p<0.05表示差異顯著。
2 結(jié)果與分析
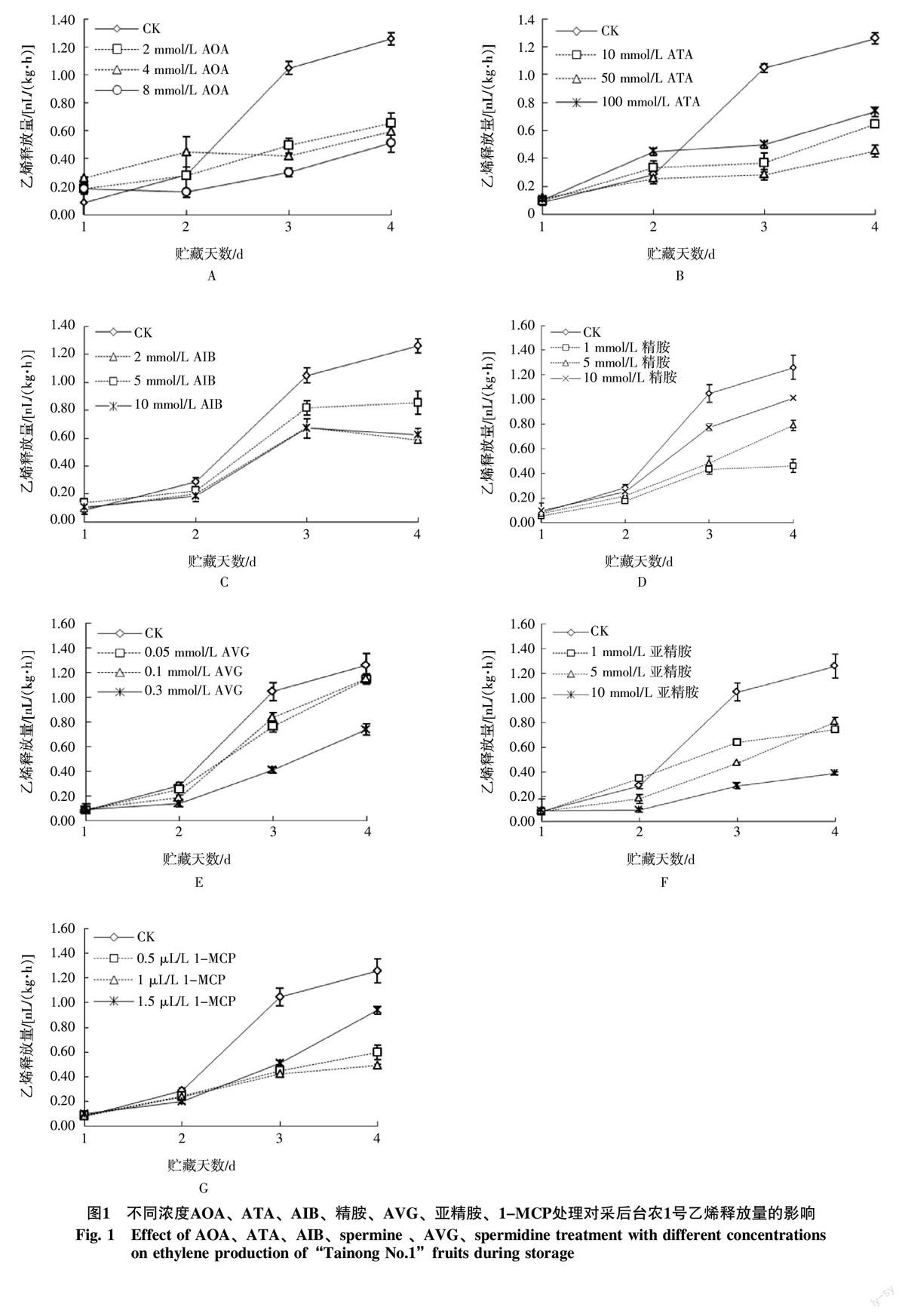
2.1 不同濃度乙烯抑制劑處理對‘臺農(nóng)杧果果實乙烯釋放量的影響
結(jié)果表明(圖1),與對照相比,不同乙烯抑制劑處理在貯藏第2天時對乙烯釋放量的抑制效果不明顯,在第3天時,對照果實乙烯釋放量明顯升高,不同濃度的乙烯抑制劑均可不同程度的降低杧果果實的乙烯釋放量(p<0.05)。貯藏至第4天,對照果實乙烯釋放量達到為1.259 nL/(kg·h);2 mmol/L AIB處理乙烯釋放量為0.589 nL/(kg·h),僅為對照的46.7%;8 mmol/L AOA處理為0.514 nL/(kg·h),為對照的40.8%;0.3 mmol/L AVG處理為0.739 nL/(kg·h),為對照的58.6%,50 mmol ATA處理為0.254 nL/(kg·h),為對照的20.1%;1 mmol/L精胺為0.46 nL/(kg·h),僅為對照的36.5%;10 mmol/L亞精胺為0.388 nL/(kg·h),僅為對照的30.8%;0.5 μL/L 1-MCP為0.489 nL/(kg·h),為對照的38.8%。綜合比較,發(fā)現(xiàn)1 mmol/L精胺、10 mmol/L亞精胺、10 mmol/L ATA對‘臺農(nóng)杧果果實采后乙烯釋放抑制效果較好(圖2)。
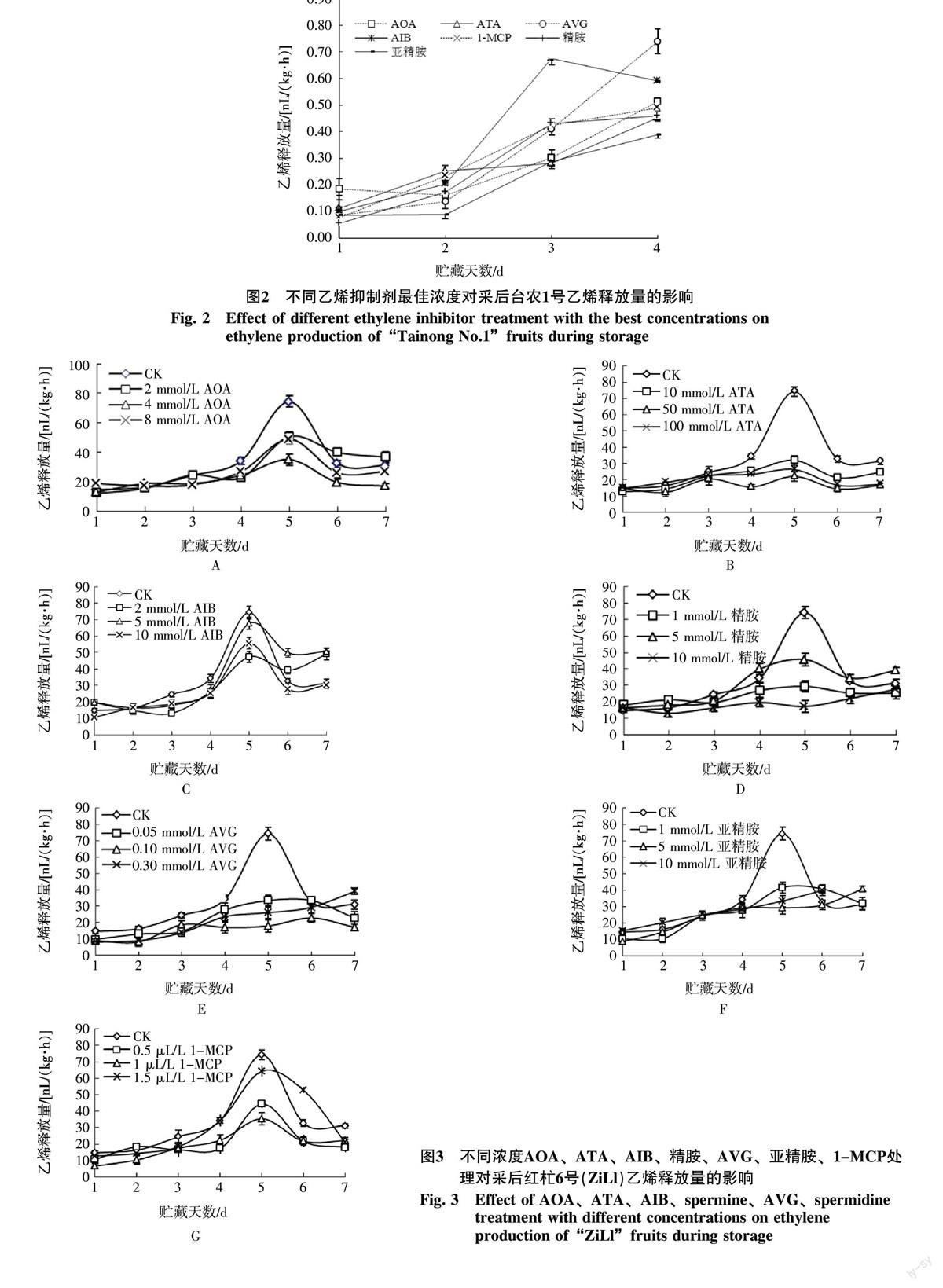
2.2 不同濃度乙烯抑制劑處理對‘紅杧果實乙烯釋放量的影響
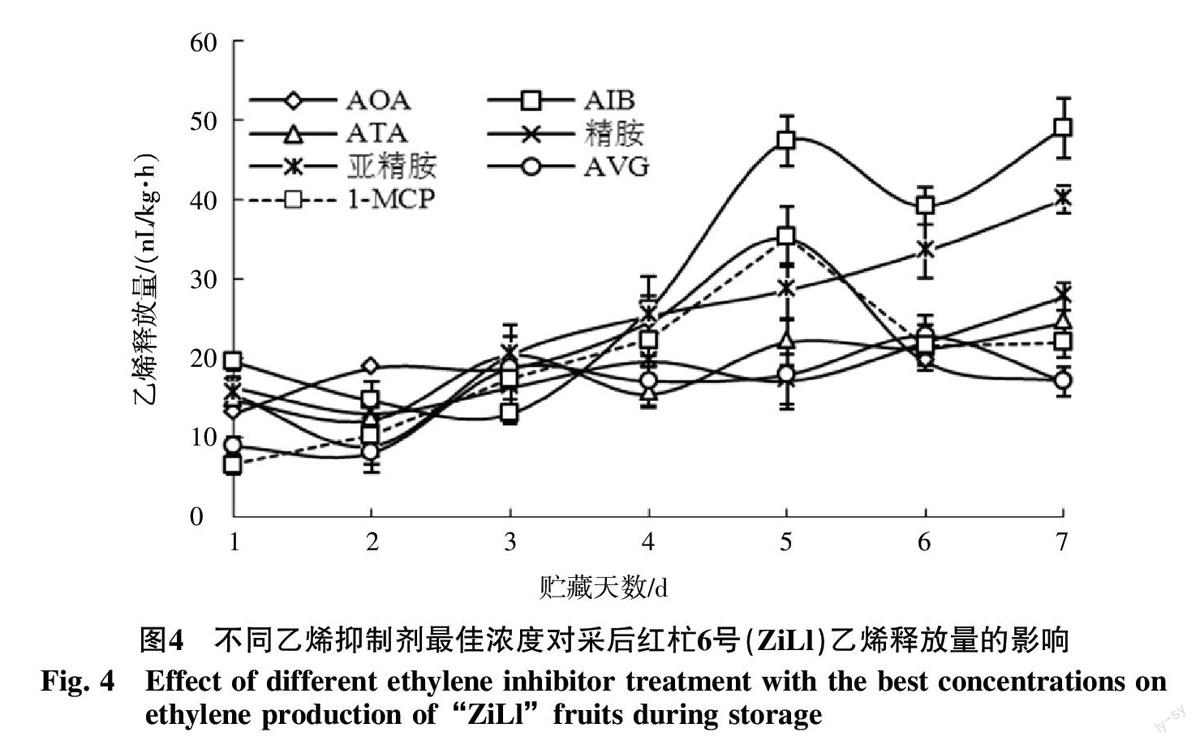
從圖3可見,對照果實采后乙烯釋放量逐漸增加,在第5天出現(xiàn)呼吸高峰,不同乙烯抑制劑處理后第2天時乙烯釋放量已低于對照,同樣乙烯釋放高峰在第5天出現(xiàn),但是顯著低于對照果實的乙烯釋放量(p<0.05)。采后第5天,對照果實乙烯釋放量為74.329 nL/(kg·h),而8 mmol/L AOA處理為49.01 nL/(kg·h),僅為對照的65.9%;2 mmol/L AIB處理為47.37 nL/(kg·h),為對照的63.7%;1 μL/L 1-MCP處理僅為35.33 nL/(kg·h),為對照的47.5%;50 mmol/L ATA、0.1 mmol/L AVG、10 mmol/L精胺、10 mmol/L亞精胺處理后,果實未出現(xiàn)明顯的乙烯釋放高峰,在整個貯藏過程中均抑制了乙烯釋放。綜合對比發(fā)現(xiàn)50 mmol/L ATA、0.1 mmol/L AVG、10 mmol/L精胺、10 mmol/L亞精胺抑制乙烯產(chǎn)生效果較好(圖4)。
2.3 精胺、亞精胺、ATA處理對杧果硬度、TSS、TA和Vc含量的影響
結(jié)果表明(圖5-A),在貯藏期間,杧果果實硬度下降速率不斷變小,以0~3 d硬度下降最快。在第3天時,ATA、精胺、亞精胺處理果實硬度顯著高于對照果實(p<0.05),對照果實硬度為2.01 kg/cm2,而處理組果實分別為2.25、2.33、2.60 kg/cm2。第6天時,對照組果實硬度下降到0.95 kg/cm2,而處理組果實則分別為1.35、1.40、1.42 kg/cm2,顯著高于對照組(p<0.05)。
杧果果實在貯藏期間,可滴定酸含量變化見圖5-B,在整個貯藏過程中,對照組和處理組果實的可滴定酸均呈現(xiàn)下降趨勢,但精胺、亞精胺、ATA處理的果實,可滴定酸含量下降速度較為緩慢,在貯藏第6 d時,精胺、亞精胺、ATA處理杧果的滴定酸含量為4.85%、4.31%和2.51%,分別為對照組杧果果實的2.9、2.66和1.55倍。
杧果果實的TSS含量變化如圖5-C,對照組杧果在貯藏的前6 d,其TSS含量逐漸升高,并在第6天達到峰值19.33%,之后開始出現(xiàn)緩慢下降,而精胺、亞精胺、ATA處理組杧果在整個貯藏期,均為上升趨勢,在第9天時分別為16%、16.83%、16.6%,尚未達到峰值,其峰值比對照推遲3 d以上,比對照的峰值低17.22%、12.9%、14.1%。
果實維生素C含量變化見圖5-D,在貯藏期間,總體上呈現(xiàn)先升后降再上升的趨勢,對照果在第3天達到峰值19.16 mg/hg,之后出現(xiàn)略微下降,在第9天時,其含量僅為19.22 mg/hg,精胺、亞精胺、ATA處理組杧果變化趨勢相同,并在第3天達到峰值,分別為20.35 mg/hg、19.86 mg/hg、22.52 mg/hg,之后緩慢下降,在第9天時含量分別為24.8 mg/hg、20.04 mg/hg、21.08 mg/hg,比對照分別高出29%、9.6%、4.2%,差異顯著(p<0.05)。
3 討論與結(jié)論
減少杧果果實乙烯氣體的產(chǎn)生、控制呼吸強度、延緩呼吸高峰的到來,及抑制植物外源和內(nèi)源乙烯的作用,是延長果實貯藏期的主要途徑[11]。本試驗結(jié)果表明,7種不同濃度不同乙烯抑制劑能夠有效地不同程度地抑制‘臺農(nóng)1號和‘紅杧6號杧果貯藏過程中乙烯的釋放,對于臺農(nóng)1號,乙烯釋放抑制效果尤以1 mmol/L精胺、10 mmol/L亞精胺、10 mmol/L ATA、0.5 μL/L 1-MCP、8 mmol/L AOA、2 mmol/L AIB、0.3 mmol/L AVG處理較好。而進一步不同藥劑處理效果比較發(fā)現(xiàn),以1 mmol/L精胺、10 mmol/L亞精胺、50 mmol/L ATA為優(yōu)于其它藥劑處理;在紅杧6號(Zill)采后貯藏過程中抑制乙烯釋放效果最佳濃度的乙烯抑制劑有:10 mmol/L精胺、10 mmol/L亞精胺、50 mmol/L ATA、1 μL/L 1-MCP、8 mmol/L AOA、2 mmol/L AIB、0.1 mmol/L AVG,經(jīng)過綜合比較又以10 mmol/L精胺、10 mmol/L亞精胺、50 mmol/L ATA、0.1 mmol/L AVG效果較好。說明采后乙烯抑制效果因乙烯抑制劑的濃度及杧果品種的不同而存在差異。在采后貯藏過程中,果實品質(zhì)的保持是評估采后保鮮處理方法是否有效合理的關(guān)鍵指標(biāo)[12]。本試驗結(jié)果表明,1 mmol/L精胺、10 mmol/L亞精胺、50 mmol/L ATA均能有效地保持果實的品質(zhì)并在一定程度上延緩杧果果實的后熟。
國內(nèi)外學(xué)者前期研究表明,減少貯藏環(huán)境中乙烯氣體或者阻斷乙烯作用途徑,可以在一定程度上代替低溫延緩果實后熟衰老[13]。高錳酸鉀結(jié)合MA包裝處理可使香蕉在30 ℃貯藏28 d,在25 ℃貯藏38 d,因此尋找合適的乙烯作用抑制劑是取代冷鏈(高能耗高成本)的低碳環(huán)保的有效方式。1-MCP具有結(jié)構(gòu)簡單、使用劑量小、高效和無污染等特點[14],目前已在蘋果[7]、桃[8]、番木瓜[9]和香蕉[10]中廣泛應(yīng)用,芒果是一種比較特殊的呼吸躍變型果實,ETR類型受體基因(METR1)的表達水平隨果實成熟進程推進及乙烯合成能力增強而增加,因而單獨應(yīng)用1-MCP在乙烯受體水平上控制芒果后熟衰老的措施效果并不明顯。ATA(3-氨基1,2,4-三氮唑)可以阻斷腺苷蛋氨酸合成ACC從而抑制乙烯合成,50 mmol/L ATA處理康乃馨顯著抑制其呼吸躍變的產(chǎn)生并延緩其衰老[15],目前弓德強等已將ATA作為杧果采后保鮮劑的有效成分應(yīng)用在杧果采后抗病保鮮中。AOA(氨基氧乙酸)和AVG(氧基乙氧基乙烯基甘氨酸)通過抑制ACS催化所需要的輔助因子磷酸吡哆醛來抑制ACS活性[16],AIB則可抑制ACO酶的活性[17]。胡偉等[21]研究表明,外源AOAA處理香蕉可通過下調(diào)MaACS1和MaGAD1基因表達延緩香蕉果實生理躍變,推遲香蕉果實采后成熟。土耳其果樹研究工作站發(fā)現(xiàn)采前7天噴施AVG 150 mg/L可顯著降低乙烯產(chǎn)生速率,延緩果實品質(zhì)下降。Wang[18]等發(fā)現(xiàn)AVG處理抑制了蘋果果實采后貯藏期的乙烯釋放并延緩其軟化過程,此外還有效地抑制了果實虎皮病的發(fā)生。杧果在常溫下對乙烯敏感性較高,AOA、AVG、AIB在杧果上的應(yīng)用至今未見報道。精胺和亞精胺屬于多胺(Pas),多胺類物質(zhì)與果實的乙烯生物合成特性有密切關(guān)系,這一點已在番茄上得到證實[19]。張輝等[20]等發(fā)現(xiàn)適宜濃度精胺亞精胺處理杏果實可通過抑制ACO、ACS活性而降低內(nèi)源乙烯生成量,從而延緩果實品質(zhì)下降。目前精胺亞精胺在抑制采后杧果果實乙烯釋放上的研究也未見報道。與傳統(tǒng)的乙烯合成抑制劑AOA、AVG、AIB相比,精胺、亞精胺、ATA抑制乙烯合成效果較好,本研究中確定不同品種杧果最適宜的乙烯抑制劑及使用濃度,根據(jù)乙烯與溫度互作原理,減少乙烯釋放量而相對提高貯藏溫度,可以緩解冷鏈低溫運營高成本高能耗的難題,是采后物流保鮮中亟待探尋的一種有效方法。
參考文獻
[1] Emongor V E. The Effects of Temperature on Storage Life of Mango(Mangifera indica L.)[J]. American Journal of Experimental Agriculture, 2015, 5(3): 252-261.
[2] 李日旺, 黃國弟, 蘇美花, 等. 我國杧果產(chǎn)業(yè)現(xiàn)狀與發(fā)展策略[J]. 南方農(nóng)業(yè)學(xué)報, 2013, 44(5): 875-878.
[3] 楊 楊, 范 蓓, 申 琳, 等. 杧果采后冷害發(fā)生及控制技術(shù)研究進展[J]. 食品科學(xué), 2014, 35(7): 292-297.
[4] Burg S P, Burg E A. Role of ethylene in fruit ripening[J]. Plant Physiology, 1962, 37: 179-189.
[5] 何全光, 任 惠, 蘇偉強, 等. 1-MCP 對田陽香杧采后生理指標(biāo)和貯藏品質(zhì)的影響[J]. 廣西農(nóng)業(yè)科學(xué), 2008, 39(1): 80-83.
[6] 李富軍, 翟 衡, 楊洪強, 等. 1-MCP對富士果實貯藏期間乙烯合成代謝的影響[J]. 中國農(nóng)業(yè)科學(xué), 2004, 5: 734-738.
[7] Bizjak J, Slatnar A, Stampar F, Veberic R. Changes in quality and biochemical parameters in ‘Idared apples during prolonged shelf life and 1-MCP treatment[J]. Food Science and Technology International, 2012, 18(6): 569-577.
[8] Jiang L, Zhang L, Shi Y, et al. Proteomic analysis of peach fruit during ripening upon post-harvest heat combined with 1-MCP treatment[J]. Journal of Proteomics, 2014, 98: 31-43.
[9] Huerta-Ocampo J A, Osuna-Castro J A, Lino-Lopez G J, et al. Proteomic analysis of differentially accumulated proteins during ripening and in response to 1-MCP in papaya fruit[J]. Journal of Proteomics, 2012, 75(7): 2 160-2 169.
[10] Botondi R, De Sanctis F, Bartoloni S, et al. Simultaneous application of ethylene and 1-MCP affects banana ripening features during storage[J]. Journal of the Science of Food and Agriculture, 2014, 94(11): 2 170-2 178.
[11] 馮敘橋, 孫海娟, 何曉慧, 等. 1-MCP應(yīng)用于杧果貯藏保鮮的研究進展[J]. 食品與生物學(xué)報, 2013, 12(32): 1 233-1 243.
[12] 張紅印, 王 雷, 姜 松, 等. 熱水處理對草莓采后病害的抑制作用及對貯藏品質(zhì)的影響[J]. 農(nóng)業(yè)工程學(xué)報, 2007, 23(8): 270-273.
[13] Wills R B H,.HarrisD R,SpohrL J,etal.Reduction of energy usage during storage and transport of bananas by management of exogenous ethylene levels[J]. Postharvest Biology and Technology, 2014, 89: 7-10.
[14] 周擁軍, 郜海燕, 陳文煊, 等. 1-MCP處理對翠冠梨貯藏效果的影響[J]. 浙江農(nóng)業(yè)學(xué)報, 2006, 18(2): 121-124.
[15] Altman S A, Solomos T. 3-amino-1,2,4-triazole, a catalase inhibitor, prolongs carnation vaselife[J]. HortScience, 1990, 25(9): 1127.
[16] 賈培義, 李春嬌, 董 麗. 乙烯抑制劑在采后觀賞植物中的應(yīng)用研究進展[J]. 園藝園林科學(xué), 2006, 22: 246-251.
[17] Toshima H N, Niwayama Y, Nagata H. Inhibition effect of coronamic acid derivates on senescence in cut carnation flowers[J]. Bioscience, Biotechnology, and Biochemistry, 1993, 57: 1 394-1 395.
[18] Wang Z, Dilley D R. Aminoethoxyvinylglycine, combined with ethephon, can enhance red color development with overripening apples[J]. HortScience, 2001, 36: 328-331.
[19] Dibble A R G. Davies P J, Mutschler M A. Polyamine content of long-keeping alcobaca tomato fruit[J]. Plant Physiology, 1988, 86: 338-340.
[20] 張 輝, 朱新衛(wèi), 車鳳斌, 等. 多胺對賽買提杏品質(zhì)及乙烯代謝的影響[J]. 新疆農(nóng)業(yè)科學(xué). 2013, 50(7): 1 213-1 219.
[21] 胡 偉, 顏 彥, 徐碧玉, 等. 氨氧乙酸對香蕉采后成熟的調(diào)控及其生理機制[J]. 熱帶作物學(xué)報, 2015, 36(2): 377-383.
