套種于不同樹齡橡膠林下謝君魔芋(Amorphophallus xiei)的光合特性研究
2016-05-30 23:10:22李珍謝世清徐文果巖所李建吳詩斌陳軍文
熱帶作物學報 2016年2期
李珍 謝世清 徐文果 巖所 李建 吳詩斌 陳軍文
摘 要 以生長在3 a和5 a樹齡橡膠林下的謝君魔芋為實驗材料,探索不同樹齡橡膠林下謝君魔芋的光合作用和光合誘導能力,以期揭示喜陰作物謝君魔芋對不同樹齡橡膠林下光的適應性。結果表明,在5 a橡膠林下謝君魔芋的暗呼吸(p<0.05)、光補償點(p<0.01)、光飽和點(p=0.459)明顯較小。在3 a橡膠林下謝君魔芋完成30%、50%、90%光合誘導進程的時間明顯較短;相對應地光合誘導1、5、10 min時的誘導狀態相對較高。這些結果說明,遮陰度增加時,謝君魔芋通過減小暗呼吸速率、光補償點等光合生理行為,降低呼吸消耗、增強對低光的利用能力來維持碳收獲,從而保證自身的正常生長;另一方面,遮陰度增加將導致謝君魔芋的光合誘導速率相對較慢,從而限制其對光斑的利用效率。因此,生產實踐中,當橡膠林冠層的遮陰度過高時,可適當采取修枝措施,增強林下套種的謝君魔芋的光合作用能力和光斑利用效率。
關鍵詞 謝君魔芋;光合特性;套種;橡膠林
中圖分類號 Q945.11 文獻標識碼 A
Photosynthetic Characteristics of Amorphophallus xiei
Intercropped to Rubber Plantation with Different Ages
LI Zhen1, XIE Shiqing1, XU Wenguo2, YAN Suo2,
LI Jian3, WU Shibin3, CHEN Junwen1*
1 College of Agronomy and Biotechnology. Yunnan Agricuhural University, Kunming,Yunnan 650201, China
2 Extension Center of Agricultural Technology of Dehong State, Mangshi,Yunnan 678400, China
3 Manager of Seed of Xishuangbanna State, Jinghong,Yunnan 666100, China
Abstract The purpose of the present study was to investigate the photosynthetic capacity and photosynthetic induction of Amorphophallus xiei, and to reveal the light adaptative mechanism of A. xiei grown under rubber plantation with different ages. The photosynthetic capacity and, photosynthetic induction characteristics were examined in A. xiei grown under rubber plantation with 3-and 5-year-old(3 a and 5 a). The result demonstrated that dark respiration rate(p<0.05), light compensation point(p<0.01) and light saturation point(p=0.459) significantly decreased in individuals grown under 5a rubber plantation. Meanwhile, in the process of photosynthetic induction, the time to reach 30%, 50% and 90% maximum photosynthesis were shorter and the induction state of 1 min, 5 min, 10 min were higher under the 3a rubber plantation. Those suggested that, with the increase in shade degree, the normal growth of A. xiei is depending on the decrease in dark respiration and light compensating point both of which maintains the harvest of carbon by enhancing utilization of low light and reducing respiration consumpation. On the other hand, the photosynthetic induction rate became slower and the use efficiency of sunflecks was depressed with the increase in shade degree. Thus, in the production practice, rubber tree branches should be pruned moderately to enhance the photosynthetic capacity and the utilization of sunflecks of A. xiei, when the shade degree was too high.
Key words Amorphophallus xiei;Photosynthetic performance;Intercropping;Rubber plantation
doi 10.3969/j.issn.1000-2561.2016.02.005
光是植物進行光合作用的主要能源,影響著植物的生長發育;同時也是重要的環境生態因子,影響著植物生態系統結構和功能[1-2]。然而,自然界中的光是高度變化的,在不同季節(春、夏、秋、冬)、不同時間段(早、中、晚)、不同天氣條件和不同空間位置(林下、林緣、林窗)光照強度都會發生改變。一般認為,5%~10%的自然光會射入林中,且因森林類型的不同而發生變化[3]。林下光斑是林下植物進行光合作用的主要能量來源。然而,受自然光照強度和植被冠層的動態影響,林下的光資源變化復雜,屬于一種高度異質化的動態資源。因此,認為光照環境是限制林下經濟作物生長發育的重要因子[4]。
目前,國內外已有大量關于林下植物對光的適應性研究,大多數研究都表明植物為適應林下的特殊光照環境,除了改變其葉片比葉重、枝葉傾角、冠幅、葉片厚度等形態結構特征外[5-8],同時也改變葉片葉綠素、光補償點(LCP)、光飽和點(LSP)、暗呼吸速率(Rd)、凈光合速率(Anet)等光合生理生化特征[5, 9-12]。然而,林下光源主要來自瞬時變化的光斑,這樣林下植物對光斑的利用能力直接影響著自身的生長發育,其中葉片光合誘導速率的快慢很大程度上決定著植物對光斑的利用能力[13]。近幾十年來,在研究植物光合作用對光斑的響應時,人們發現光合誘導速率快慢除了與植物種類[13]、葉齡[14]、光合酶的激活狀態[15]等有關外,植物所處的光照環境是決定其誘導特征的重要因素[16-17]。
謝君魔芋(Amorphophallus xiei)為天南星科魔芋屬的一種多年生宿根草本植物,主要分布在云南南部德宏州的熱帶地區。謝君魔芋起源于熱帶雨林,喜溫暖,忌高溫,喜散射光和弱光,忌強光直射,是一種典型的喜陰作物[18]。因此,在生產中謝君魔芋常與高桿作物或經濟林進行間套作種植。有報道認為,喜陰的謝君魔芋與喜光的橡膠樹套作形成的經濟林有利于兩者的生長,從而有效提高土地利用率[19]。不同樹齡橡膠林受其樹冠大小的影響,林間遮陰度即光照環境不相同。本研究以套種在3和5 a樹齡橡膠林下的謝君魔芋為實驗材料,研究謝君魔芋的光合作用能力和光合誘導能力,以期揭示喜陰作物謝君魔芋對不同樹齡橡膠林下光照環境的適應性。
1 材料與方法
1.1 試驗地概況
本試驗于2011年4月~11月在云南省德宏傣族景頗族自治州芒市遮放鎮(東經98°10′,北緯24°11′)戶拉村進行。遮放鎮平均海拔1 160 m,屬南亞熱帶季風氣候,年平均氣溫為19.8 ℃,年日照在2 000~2 452 h之間,無霜期達300 d以上,年平均降雨量1 300~1 653 mL,雨季集中在5~10月,雨量充沛,氣候溫和。試驗地土壤的基本理化性狀是:pH (5.75±0.02),有機質含量(40.12±3.89)g/kg,全N含量(2.15±0.19)g/kg, 全P含量(0.48±0.05)g/kg, 全K含量(17.29±3.06)g/kg。
1.2 方法
1.2.1 試驗設計 選擇土壤深厚、質地疏松、地勢平坦的3和5 a樹齡的三葉橡膠林套種魔芋。橡膠林種植規格為行距6 m,株距4 m;3 a樹齡橡膠林的透光率為35%左右,5 a樹齡橡膠林的透光率為20%左右。試驗設置3和5 a兩個處理,重復5次,共10個小區,每個小區長8.5 m,寬3.5 m,小區間隔1 m。先撒生石灰750 kg/hm2對土壤進行消毒,隨后整地,起壟(壟寬70~80 cm,壟高20~25 cm),做溝(30 cm)。磷肥和鉀肥作為底肥一次性施用,用量按120 kg/hm2 P2O5和100 kg/hm2 K2O比例進行施用;氮肥作為追肥施用(150 kg/hm2 N),其中出苗期施用30%,生長旺盛期施用70%(7月初)。試驗選取重量150 g左右健康的種芋作為實驗材料,用農用鏈霉素以1 ∶ 10兌水浸種30 min,晾曬,待種芋出芽后,于2011年4月20日播種,種植規格為行距0.8 m,株距0.5 m。在謝君魔芋生長期間,定時進行病蟲草害的防控,于魔芋生長旺盛期進行各項指標測定。
1.2.2 相關指標測定方法 光響應曲線的測定。
當年8月中旬,從每個小區中隨機選取完全展開的成熟健康葉片,利用美國Li-COR公司生產的Li-6400XT便攜式光合儀測定謝君魔芋的凈光合速率,參照張強等[20]描述的方法進行測定。不同的是參比室CO2濃度設為380 μmol/mol,誘導光強為800 μmol/(m2·s),誘導3~5 min,自動測量程序設定光強梯度為1 500、1 200、1 000、800、600、400、200、150、100、80、60、40、20、0 μmol/(m2·s)。參照Webb[21]計算公式計算相關光合參數:實時凈光合速率(P),光補償點(LCP),光飽和點(LSP),暗呼吸速率(Rd),表觀量子產額(AQY),瞬時水分利用效率(WUE)。
CO2響應曲線的測定。選取測光響應曲線時的葉片,同樣用Li-6400XT便攜式光合儀測定CO2響應曲線[20]。除了CO2濃度外其他設置與光響應曲線測定設置的條件相同。設定CO2梯度為400、300、200、150、100、50、400、600、800、1 200、1 500 μmol/mol,等待時間為2~3 min。參照Bernacchi[22]計算公式求出最大羧化速率Vcmax和最大電子傳遞速率Jmax。
光合誘導曲線的測定。光合誘導曲線測定同樣參照張強等[20]的測定方法,不同的是,低光照強度設為20 μmol/(m2·s),記錄時間為4 min,而瞬時轉換到高光照的強度為1 500 μmol/(m2·s),記錄時間20 min,光合誘導期間每隔2 s記錄1個數據。以光照時間為橫坐標,光合速率為縱坐標建立光合誘導曲線,并依據曲線計算誘導過程中凈光合速率(Anet)達到最大光合速率(Amax)30%、50%和90%的時間(t30%A、t50%A和t90%A)。同時,根據公式100×Anet/Amax [23]計算1、5、10 min的光合誘導狀態IS1 min、IS5 min、IS10 min。
1.3 數據統計與分析
利用Excel 2003及SPSS 14.0軟件對數據進行統計分析,采用獨立樣本t檢驗進行顯著性分析,當p≦0.05時,認為處理間差異顯著。運用Sigmaplot10.0軟件作圖。
2 結果與分析
2.1 不同樹齡橡膠林下的謝君魔芋的穩態光合特性
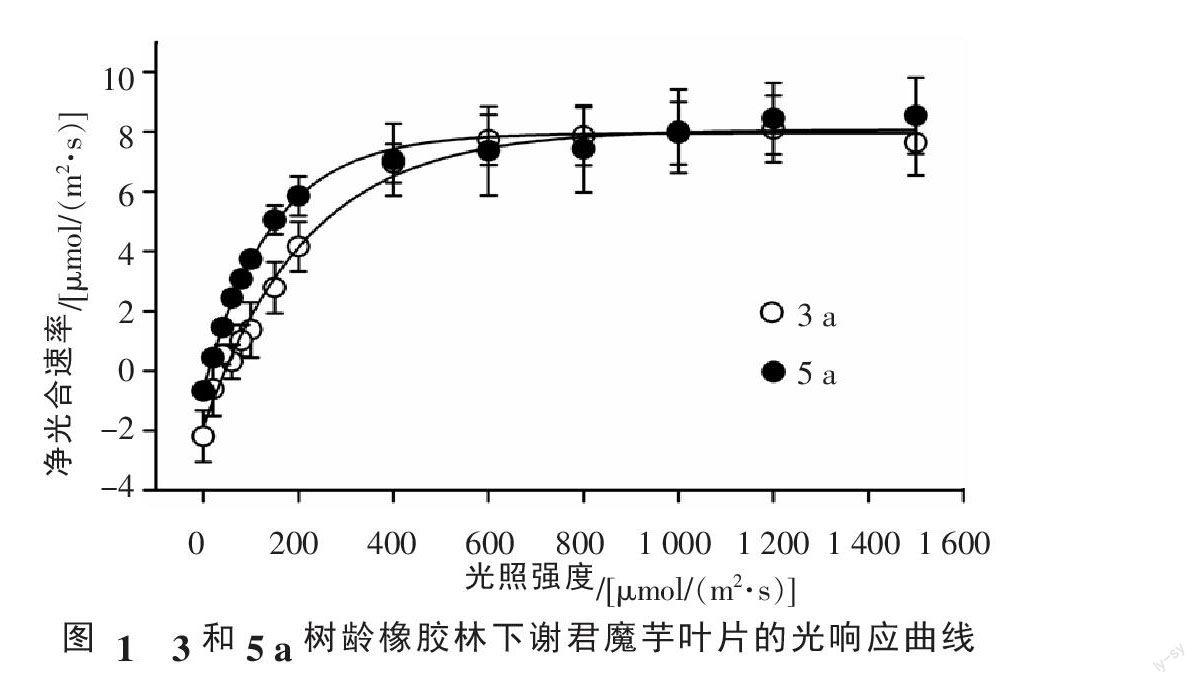
3 a樹齡、5 a樹齡橡膠林下謝君魔芋葉片具有類似的光響應曲線變化趨勢,即有快速增長階段和緩慢增長到穩定態階段(圖1)。然而,不同樹齡橡膠林下的謝君魔芋光合特性存在明顯差異(表1),5 a橡膠林下的謝君魔芋的暗呼吸(Rd)(P=212.628,F=0.000)和光補償點(LCP)(P=8.108,F=0.036)顯著小于3 a橡膠林下謝君魔芋的Rd和LCP。雖然在統計意義上兩個處理間謝君魔芋的光飽和點(LSP)、最大氣孔導度(gs-max)、水分利用效率(iWUE)無顯著差異(p>0.05),但數據中5 a橡膠林下謝君魔芋的LSP、gs-max、iWUE的值相對較小。此外,謝君魔芋在5 a橡膠林下的最大凈光合速率(Amax)和表觀量子產額值(AQY)在兩處理間無明顯差異。
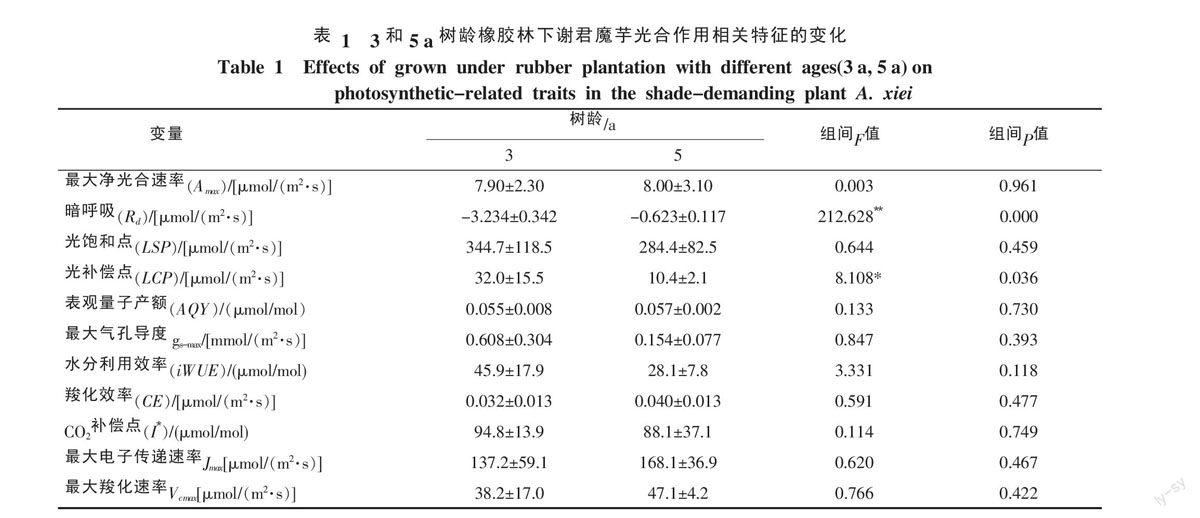
CO2是進行光合作用的重要原料,植物在光合作用中將CO2轉變為碳水化合物,從而固定太陽能。從圖2中看出,隨著胞間CO2濃度的增加,兩個處理下的謝君魔芋凈光合速率都增大。然而,謝君魔芋的羧化效率(CE)、最大電子傳遞速率(Jmax)、最大羧化速率(Vcmax)在5 a橡膠林下要比3 a橡膠林下的稍大。
2.2 不同樹齡橡膠林下的謝君魔芋在連續飽和強光下光合誘導的時間進程
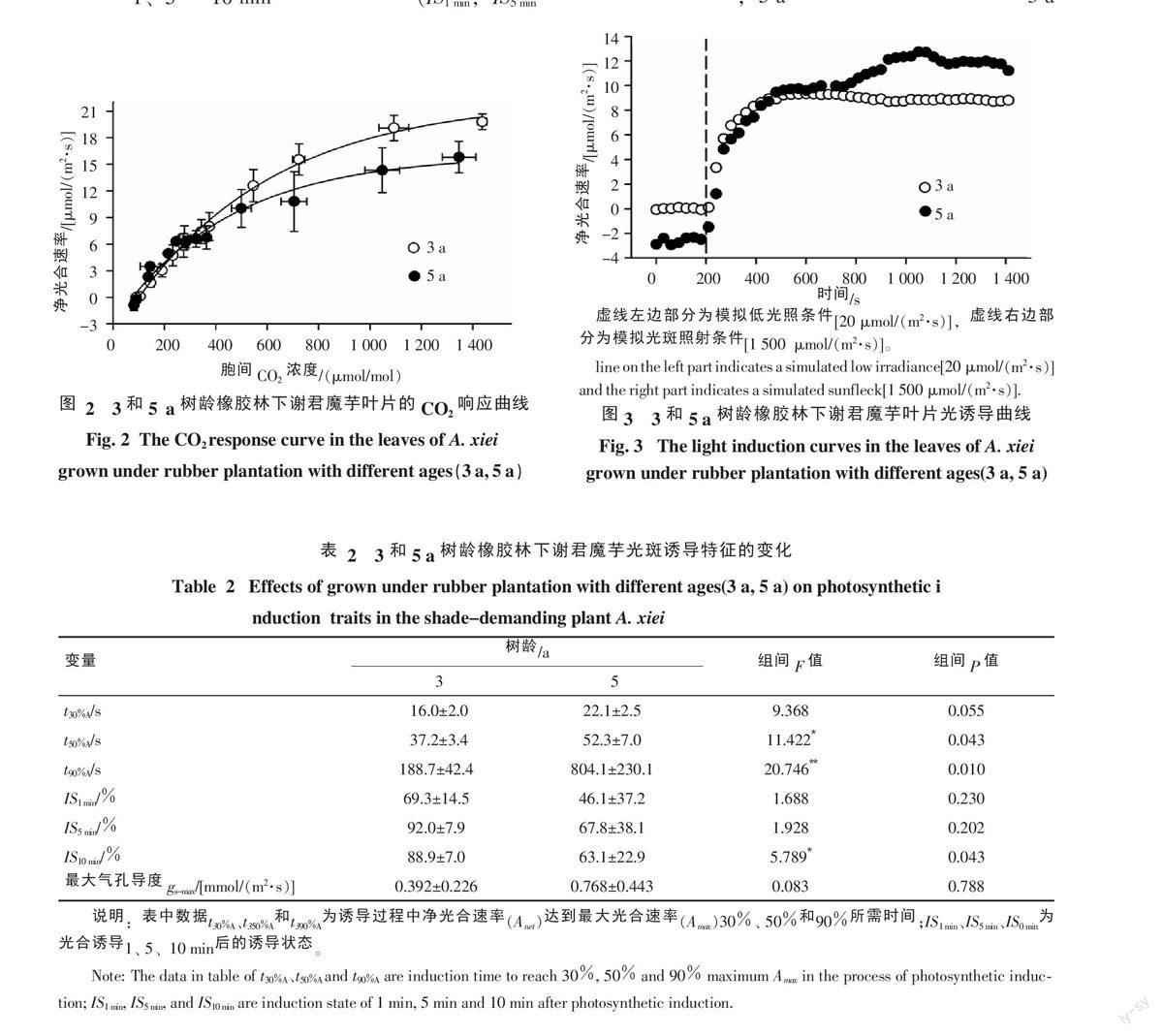
當從低光照[20 μmol/(m2·s)]瞬時變換到飽和強光[1 500 μmol/(m2·s)]時,兩種處理下的謝君魔芋的凈光合速率的誘導曲線呈半S形狀(圖3)。然而,兩個處理間謝君魔芋的光合誘導特性存在明顯差異(表2),t50%A和t90%A的值在5 a橡膠林下顯著最大,說明達到最大光合速率所需時間較長、光合誘導速率較慢,并且兩處理間的差異隨誘導時間的延長表現越顯著;相對應地,5 a橡膠林下的謝君魔芋在1、5和10 min時的誘導狀態(IS1 min,IS5 min和IS10 min)相對3 a橡膠林下較低。
3 討論與結論
光是影響植物生長發育和分布的主要環境因子之一[24]。受樹冠遮陰影響,林下光照環境復雜多變,植物通過調節生理生化特性以適應光環境的變化。植物光合作用能力適應性變化的大小是衡量其對生境適應性的重要指標[25]。前人研究發現,隨著上層樹冠蔭蔽度的增加及林內光照強度降低,林下植物通過降低光補償點和光飽和點以及增加表觀量子效率來提高對光的捕捉及利用效率[24];同時通過降低暗呼吸、提高碳水化合物的凈積累,保證植物凈碳獲得和正向生長[25]。
在本試驗中,5 a橡膠林下的謝君魔芋與3 a橡膠林下的謝君魔芋相比具有較小的Rd、LCP、LSP以及較大的Vcmax和Jmax(表1),可能由于5 a橡膠林下光照強度相對較小,謝君魔芋通過降低LCP和LSP來提高對弱光的利用能力;同時,通過降低呼吸速率減少能量消耗來維持體內碳平衡。蔡仕珍等[25]研究林下植物蝴蝶花(IIris japonica Thunb.)發現Rd、LCP、LSP隨著植株上層光強的降低而減小,這是蝴蝶花對低光的適應反應。研究結果與前人類似的研究[26-27]都表明,當生長環境光照強度降低時陰生植物通過降低碳消耗和增強碳收獲能力來適應低光環境。同時發現,5 a橡膠林和3 a橡膠林下的謝君魔芋Amax值相近,這說明陰生作物謝君魔芋在低光生長環境中保持最大光合作用能力及最大碳收獲是其優先的適應策略。
光斑是林下植物進行光合作用的主要能量來源,生長環境中頻繁遭受光斑照射的植物其光合誘導速率一般較快[28]。然而光合誘導受到光合碳同化酶[15]、光照環境[29]、光合碳同化中間產物積累[30]及葉片氣孔[31]的影響。其中初始氣孔導度與光合誘導時間存在著負相關關系[32]。在3 a橡膠林下,謝君魔芋t30%A、t50%A、t90%A相對較短,且IS1 min、IS5 min、IS10 min相對較大,說明3 a橡膠林下的謝君魔芋光合誘導速率較快,表征光斑的利用能力較強。這可能是由于3 a橡膠林下的較高透光率有利于激活光合酶和光合底物產生,促進光合誘導的進行[33-34]。此外,張守仁等[35]認為植物對光的響應還取決于氣孔密度,一般來說高光照下氣孔密度相對要大[36]。這也暗示著3 a橡膠林下的謝君魔芋氣孔密度可能較大,有利于促進誘導的快速發生,但這還有待進一步研究。
總之,本研究結果顯示,在5 a橡膠林的低光環境下,謝君魔芋通過減小Rd、LCP等光合生理行為,降低呼吸消耗、增強對低光的利用能力來維持碳收獲,從而保證自身的正常生長。另一方面,在5 a橡膠林下謝君魔芋其光合誘導速率相對較慢,這將限制其對林下光斑的高效利用。因此,在生產中,當橡膠林冠層的遮陰度過高,可適當采取修枝措施,增加謝君魔芋對林下光斑的利用效率。
參考文獻
[1] 陳曉亞, 湯章城. 植物生理與分子生物學[M]. 北京: 高等教育出版社, 2007: 375-397.
[2] Bazzaz F A, Pickett S T A. Physiological ecology of tropical succession: a comparative review[J]. Annu Rev Ecol Syst, 1980, 11: 287-310.
[3] 李 燁, 褚國英. 林下光環境研究進展及其對經濟植物生長的影響[J]. 山東林業科技, 2009(2): 131-133.
[4] Bazzaz F A. Plants in changing environments: linking physiological, population, and community ecology[M]. Cambridge: Cambridge University Press, 1996: 13-38.
[5] Osada N, Takeda H, Kitajima K, et al. Functional correlates of leaf demographic response to gap release in saplings of a shade-tolerant tree, Elateriospermum tapos[J]. Oecologia, 2003, 137(2): 181-187.
[6] Malavasi U C, Malavasi M M. Leaf characteristics and chlorophyll concentration of Schyzolobium parahybum and Hymenaea stilbocarpa seedlings grown in different light regimes[J]. Tree Physiol, 2001, 21(10): 701-703.
[7] 張 健, 張金政, 姜闖道, 等. 鳶尾(Iris L.)葉片取向與其光合特性及光抑制的關系[J]. 生態學報, 2008, 28(8): 3 637-3 643.
[8] 陸鑾眉, 閻光宇, 杜曉娜, 等. 8種園林植物耐蔭性與光合特性的研究[J]. 熱帶作物學報, 2011, 32(7): 1 249-1 254.
[9] 王立豐, 陸艷茜, 鄧玉杰. 光照強度對巴西橡膠樹淡綠期葉片光合特性和活性氧代謝的影響[J]. 熱帶作物學報, 2014, 35(6): 1 131-1 136.
[10] Pons T L, Pearcy R W. Nitrogen reallocation and photosynthetic acclimation in response to partial shading in soybean plants[J]. Physiol Plant, 1994, 92(4): 636-644.
[11] Leroux X, Walcroft A S, Daudet F A, et al. Photosynthetic light acclimation in peach leaves: importance of changes in mass: area ratio, nitrogen concentration, and leaf nitrogen partitioning[J]. Tree Physiol, 2001, 21(6): 377-386.
[12] 劉文海, 高東升, 束懷瑞. 不同光照處理對設施桃樹光合及熒光特性的影響[J]. 中國農業科學, 2006, 39(10): 2 069-2 075.
[13] 蔡志全, 曹坤芳, 鄭 麗. 六種熱帶雨林木本植物幼苗光合誘導的研究[J]. 植物生態學報, 2003, 27(5): 617-623.
[14] 張小全, 徐德應. 杉木中齡林針葉光合作用對光斑的響應[J]. 植物生態學報, 2001, 21(3): 409-414.
[15] Pons T L, Pearcy R W, Seemann J R. Photosynthesis in flashing light in soybean leaves in different conditions. 1.Photosynthetic induction state and regulation of ribulose-1, 5-bisphosphate carboxylase activity[J]. Plant Cell Environ, 1992, 15(5): 569-576.
[16] Valladares F, Allen M T, Pearcy R W. Photosynthetic responses to dynamic light under field conditions in six tropical rainforest shrubs occurring along a light gradient[J]. Oecologia, 1997, 111(4): 505-514.
[17] Tang Y H, Hiroshi K, Mitsumasa S, et al. Characteristics of transient photosynthesis in Quercus serrata seedlings grown under lightfleck and constant light regimes[J]. Oecologia, 1994, 100(4): 463-469.
[18] 謝世清, 張云峰, 謝慶華, 等. 魔芋的離體繁育及優質魔芋規范化栽培技術研究[M]. 昆明: 云南科技出版社, 2010: 57-58.
[19] 沈建榮, 李宗益, 劉曉蓮, 等. 西雙版納州經濟林-紅魔芋混農林業發展前景初探[J]. 中國農業信息, 2013(5): 54.
[20] 張 強, 陳軍文, 陳亞軍, 等. 西雙版納熱帶雨林中兩種生態型蕨類植物的光和特性比較研究[J]. 植物學通報, 2008, 25(6): 673-679.
[21] Webb W L, Newton M, Starr D. Carbon dioxide exchange of Alnus rubra: a mathematical model[J]. Oecologia, 1974, 17(4): 281-291.
[22] Bernacchi C J, Singsaas E L, Pimentel C, et al. Improved temperature response functions for models of Rubisco-limited photosynthesis[J]. Plant Cell Environ, 2001, 24(2): 253-259.
[23] Allen M T, Pearcy R W. Stomatal behavior and photosynthetic performance under dynamic light regimes in a seasonally dry tropical rain forest[J]. Oecologia, 2000, 122(4): 470-478.
[24] 吳統貴, 虞木奎, 孫海菁, 等. 林藥復合系統林下植物光合特性對生長光強的響應[J]. 中國生態農業學報, 2011, 19(2): 338-341.
[25] 蔡仕珍, 李 西, 潘遠智, 等. 不同光照對蝴蝶花光合特性及生長發育研究[J]. 草葉學報, 2013, 2(22): 264-272.
[26] Chen J W, Kuang S B, Long G Q, et al. Steady-state and dynamic photosynthetic performance and nitrogen partitioning in the shade-demanding plant P. notoginseng under different levels of growth irradiance[J]. Acta Physiol Plant, 2014, 36(9): 2 409-2 420.
[27] 鄭順林, 田孟良, 劉金亮, 等. 人工馴化栽培中弱光脅迫對七葉一枝花光合特性的影響[J]. 中藥材, 2014, 37(9): 1 518-1 522.
[28] Kursar T A, Coley P D. Photosynthetic induction times in shade-tolerant species with long and short-lived leaves[J]. Ocologia, 1993, 93(2): 165-170.
[29]Rijkers T, Pons T L, Bongers F. Photosynthetic induction in saplings of three shade-tolerant tree species: comparing understorey and gap habitats in a French Guiana rain forest[J]. Oecologia, 2000, 125(3): 331-340.
[30] 許大全. 光合作用效率[M]. 上海:上海科學技術出版社, 2002: 47-48.
[31] Sassenrath-Cole G F, Pearcy R W. The role of ribulose-1,5-bisphoate regeneration in the induction requirement of photosynthetic CO2 exchange under transient light conditions[J]. Plant Physiol, 1992, 99(1): 227-234.
[32] Zhang Q, Chen J W, Li B G, et al. Epiphytes and hemiepiphytes have slower photosynthetic response to lightfleck than terrestrial plants: evidence from ferns and figs[J]. J Trop Ecol, 2009, 25(5): 46-472.
[33] 張 昆, 萬勇善, 劉鳳珍. 苗期弱光對花生光合特性的影響[J]. 中國農業科學, 2010, 43(1): 65-71.
[34] 吳正鋒, 王才斌, 萬書波, 等. 弱光脅迫對花生葉片光合特征及光合誘導的影響[J]. 青島農業大學學報(自然科學版), 2010, 27(4): 277-281.
[35] 張守仁, 高榮孚. 光誘導下雜種楊無性系葉角和葉綠體的運動[J]. 生態學報, 2001, 21(1): 68-74.
[36] 蔡志全, 齊 欣, 曹坤芳. 七種熱帶雨林樹苗葉片氣孔特征及其可塑性對不同光照強度的響應[J]. 應用生態學報, 2004, 15(2): 201-204.
