1例B(A)02等位基因的鑒定及分子機制研究
2016-07-13 03:14:43邱麗么楠苗溫鄒偉蔡曉紅
天津醫(yī)藥 2016年5期
邱麗,么楠,苗溫,鄒偉,蔡曉紅
?
1例B(A)02等位基因的鑒定及分子機制研究
邱麗1,么楠1,苗溫1,鄒偉2,蔡曉紅2
目的B(A)02等位基因的鑒定及分子機制的研究。方法對1例B(A)02等位基因者ABO血清學定型使用標準血清學實驗方法,對ABO基因7個外顯子及其側翼序列做PCR擴增、基因克隆和測序分析;使用PyMOL軟件建立B糖基轉移酶(GTB)的空間模型并進行分析。結果血清學檢測結果符合B(A)亞型特點,DNA克隆和測序分析顯示被檢者為B(A)02/O01雜交基因,在B101等位基因上存在700C>G錯義突變,導致GTB發(fā)生P234A氨基酸置換。通過對GTB空間結構的分析,認為P234A置換影響了第234位氨基酸與Met-266的分子間作用力,從而改變了GTB識別供糖體的結合凹槽的結構,使GTB能夠結合并轉移N-乙酰氨基半乳糖(GalNac)至底物H抗原,從而在紅細胞膜上形成弱A抗原。結論P234A的置換影響了決定GTB識別特異性的Met-266與Ala-268所組成的特異性識別區(qū)域的空間結構,從而改變了ABO血型的抗原性。
ABO血型系統(tǒng);等位基因;B(A)亞型;PyMOL軟件
B(A)亞型是一種獨特的ABO變異型,表現(xiàn)為同一等位基因的編碼產物兼具A和B特異性的糖基轉移酶活性,其ABO血型表觀遺傳學不符合孟德爾規(guī)律,以順式遺傳為特點[1-2]。關于B(A)亞型的基因背景有兩種不同的假說[1]:(1)顯著增強的B轉移酶活性能夠催化N-乙酰氨基半乳糖(GalNac)從UDP-N-乙酰氨基半乳糖(UDP-GalNac)到2′-巖藻糖,形成一種具有A活性的結構。(2)編碼糖基轉移酶的基因(即ABO基因)發(fā)生錯義突變。目前研究發(fā)現(xiàn)后者更多見[3]。至2015年5月為止,人類血型基因突變庫(BGMUT)已報道的B(A)等位基因有6種[4]。1993年Yamamoto等[5]首次報道1種能夠編碼正常B糖基轉移酶(glycosyltransferases B,GTB)的ABO等位基因同時具有A糖基轉移酶(glycosyl-transferases A,GTA)活性,這個等位基因被命名為B(A)01。B(A)02、B(A)04、B(A)05和B(A)06均首先在中國人群中發(fā)現(xiàn),且以B(A)04和B(A)02的基因頻率為最高(1.6/10萬和0.78/10萬)[2]。本文旨在分析B(A)02等位基因,并初步探討其分子機制。
1對象與方法
1.1研究對象1例B(A)表現(xiàn)型的無償獻血者,男,25歲,漢族,于2014年在天津市濱海新區(qū)塘沽中心血站獻血時被檢出正反定型不符。使用EDTA-K2抗凝管采集5 mL獻血者外周靜脈血液進行血清學和分子生物學檢測。
1.2試劑與儀器單克隆抗-A、抗-B(批號20131224)、抗-H血清(批號20130408)、抗-A1血清(批號20140923)和ABO反定型紅細胞(批號20145345)均為上海血液生物醫(yī)藥有限公司生產。抗-AB(DIAGAST,批號439000);A2細胞(批號115247C,Immucor公司),人源抗-A(自制試劑,效價128),基因組DNA抽提試劑盒(批號M1218,天根生物產品),DNA瓊脂糖凝膠純化試劑盒(批號M00529,GeneMark產品);pMD18-T載體(批號K6601B,日本TaKaRa公司產品)。細胞洗滌離心機(KA-2200型,日本久保田),測序儀(ABI377,美國Applied Biosystems公司)。
1.3血型血清學檢測采用鹽水試管法進行ABO血型鑒定和吸收放散試驗。吸收放散試驗:取1 mL獻血者壓積紅細胞和1 mL正常O型壓積紅細胞作為對照,兩者中分別加入等體積的人源抗-A血清,混勻后放置4℃冰箱中吸收1 h,然后于56℃水浴中熱放散,兩者的放散液再分別與A1試劑紅細胞反應[6]。
1.4 DNA提取和PCR擴增反應使用基因組DNA抽提試劑盒抽提外周血基因組DNA。PCR使用的引物根據ABO基因序列(GenBank Accession Number N_000009)參照文獻[7]。引物設計,按如下條件擴增1~7外顯子:95℃3 min,95℃30 s、56℃30 s、72℃60 s,30個循環(huán),最后72℃延伸8 min。
1.5 PCR產物直接測序采用DNA瓊脂糖凝膠純化試劑盒純化PCR產物,并使用測序儀對ABO基因7個外顯子及其側翼序列直接測序分析。
1.6 PCR產物的克隆及測序將第7外顯子的PCR產物純化后克隆入pMD18-T載體,隨后應用測序儀分析PCR插入片段DNA序列。PyMOL軟件(DeLano Scientific LLC,v0.99)建立GTB三維模型。
2 結果
2.1血型血清學檢測
2.1.1 ABO血型鑒定試管法中,獻血者紅細胞與單克隆抗-A試劑反應呈弱陽性(2+),與單克隆抗-B和單克隆抗-H試劑反應均呈強陽性(4+),同時將正常O細胞、B細胞與抗-H反應作為對照,結果分別為強陽性(4+)、弱陽性(2+);獻血者血清與A1試劑紅細胞反應呈弱陽性(2+),與A2試劑紅細胞反應呈弱陽性(1+),與試劑B細胞和O細胞均不反應。血清學表現(xiàn)符合B(A)亞型特點。
2.1.2吸收放散試驗獻血者紅細胞放散液與A1試劑紅細胞反應結果為強陽性(3+),O型對照紅細胞放散液與A1試劑紅細胞反應結果為陰性,證實獻血者紅細胞上存在弱A抗原。
2.2 DNA測序獻血者的ABO基因型為B(A)02/ O01,B等位基因第7外顯子存在700C>G錯義突變,導致GTB的P234A氨基酸置換,見圖1。
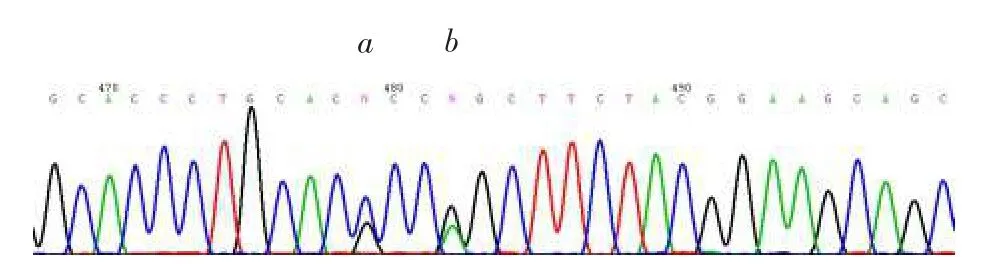
箭頭a所示為第7外顯子700C>G雜合突變;箭頭b所示為703G>A雜合突變Fig. 1 Partial sequence analysis of ABO gene sequencing圖1 ABO基因序列檢測的部分序列檢測圖譜
2.3對GTB的空間模型的分析P234與Met-266在空間結構上十分靠近,P234A置換使脯氨酸吡咯環(huán)C-γ原子處發(fā)生中空,影響其與Met-266間原有的分子間作用力,從而影響Met-266的側鏈方向,造成酶活性中心Met-266和Ala-268組成的“結合凹槽”的大小的改變,使GTB可以結合部分N-乙酰氨基半乳糖(GalNac),見圖2~5。
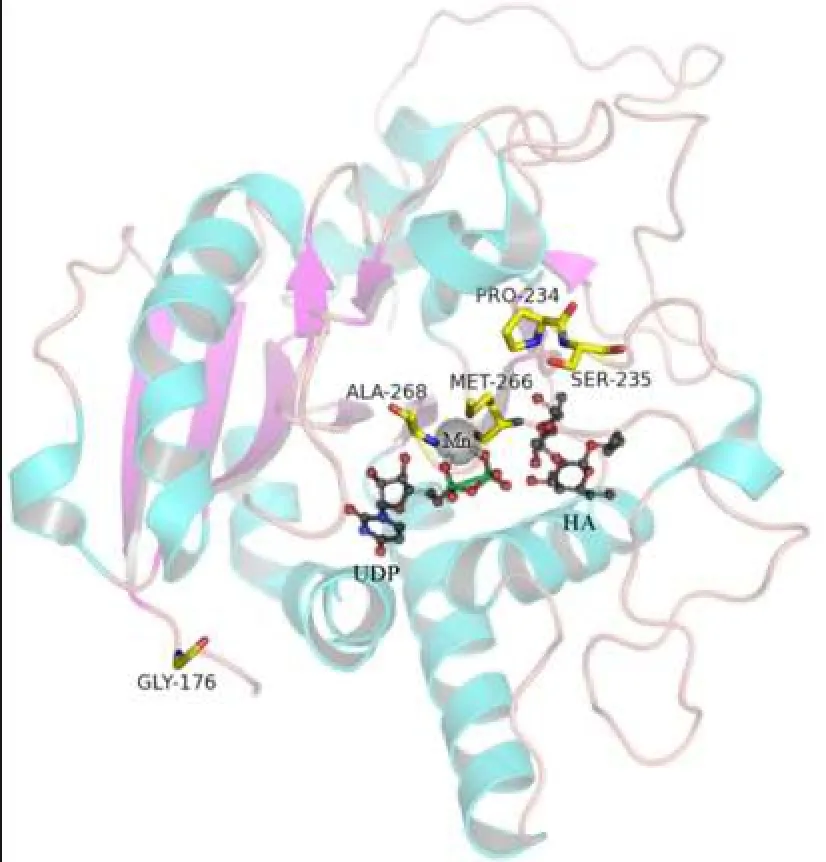
Fig. 2 Relationship between three dimensional structure of GTB and spatial position of four key amino acids(Gly-176,Ser-235,Met-266,Ala-268)and Pro-234 in GTB圖2 GTB三維結構圖(PDB ID:2RJ8)及4個關鍵氨基酸(Gly-176、Ser-235、Met-266、Ala-268)和Pro-234的空間位置關系
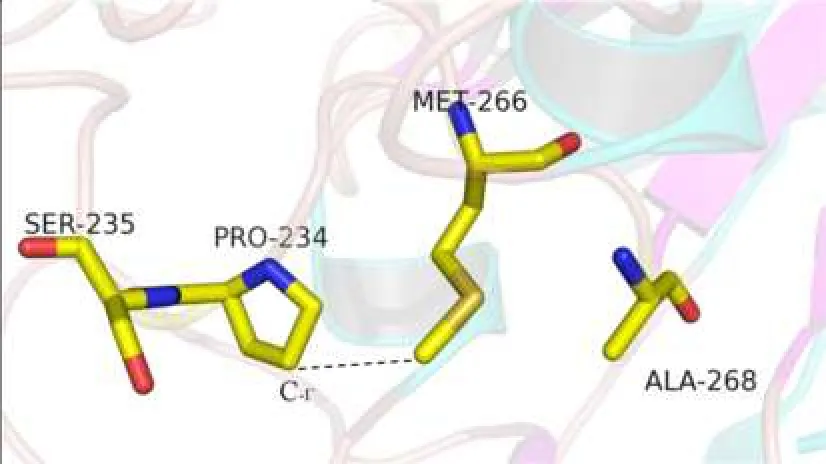
Fig. 3 Relationship between spatial position of Pro-234 and Met-266 side chain before mutation圖3 突變前Pro-234與Met-266側鏈的空間位置關系
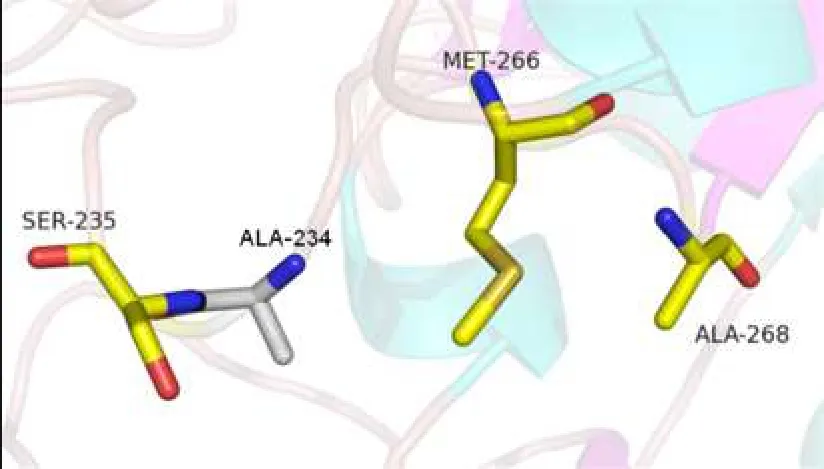
Fig. 4 Relationship between spatial position of ALA-234 and Met-266 side chain after mutation圖4 突變后ALA-234與Met-266側鏈的空間位置關系
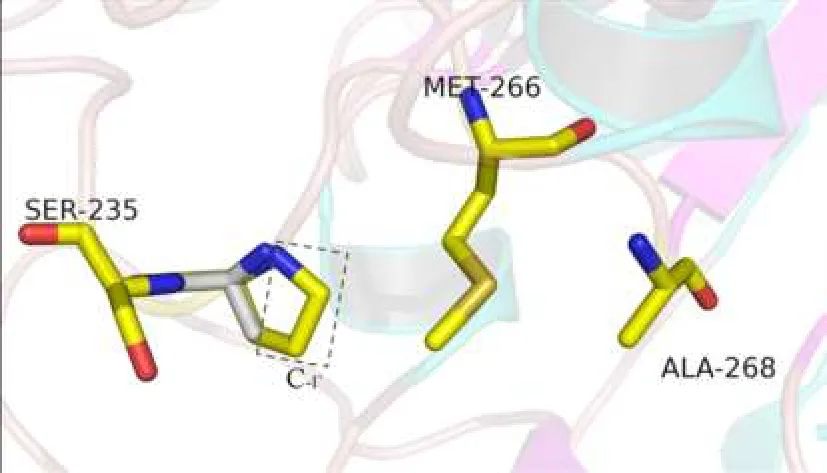
Fig. 5 The sketch diagram of the disappearance of C-γ after mutation圖5 突變后C-γ原子缺失示意圖
3 討論
人類血型A、B抗原是由糖基轉移酶催化寡糖不斷連接合成的碳水化合物結構,而特異性的糖基轉移酶是由ABO基因指導合成。A基因和B基因cDNA序列存在7個單核苷酸多態(tài)性(single nucleotide polymorphism,SNP),分別是297A>G,526C>G,657C>T,703G>A,796C>A,803G>C和930G>A,其中4個SNPs為錯義突變,造成Arg176Gly、Gly235Ser、Leu266Met、Gly268Ala這4個關鍵氨基酸置換[8]。01等位基因由于發(fā)生261delG移碼突變,編碼截短的無功能酶。人類ABO血型GTA和GTB具有高度同源性,354個氨基酸中僅4個關鍵氨基酸的不同卻造成了ABO抗原合成、酶動力學和結構的顯著區(qū)別。由于4個關鍵氨基酸中僅266R和268R占據了GTA和GTB活性反應區(qū)中能與核苷糖結合的位置,且兩者的側鏈和UDP-Gal/GalNac之間存在高度互補的空間立體化學關系[9],因此第266R和268R在酶對供糖體特異性識別和結合過程中起決定作用[10-11]。而酶動力學和晶體學研究證明,266R 比268R更為關鍵,因為只有266R是與核苷糖中顯有供糖體特異性的基團(Gal的羥基和GalNac的乙酰氨基)結合并反應,從而使GTA、GTB區(qū)別供糖體的特異性而形成A、B抗原,268R則是與兩種核苷糖共有的相同基團結合[11-12]。
本研究中的獻血者因ABO正反定型不一致,進一步檢測發(fā)現(xiàn)其紅細胞定型為AweakB特點,且與抗-H反應強度接近O細胞,血清中存在抗-A抗體,血清學表型符合B(A)亞型的特點。測序表明其基因型為B(A)02/O01,其B基因在B101的基礎上發(fā)生700C>G突變,導致第234位脯氨酸置換為丙氨酸,該突變使B基因帶有了A基因的特點。對于B(A)02等位基因導致B(A)表型分子機制的深入研究,國內文獻少見報道,僅有文獻推測是由于第234位氨基酸與Ser-235緊密相連,空間結構非常靠近,從而造成了影響[13-14],但論據缺乏。而國外研究認為,Ser-235的作用是影響酶-供糖體復合物與底物H抗原的識別和結合,對酶識別供糖體的特異性并無影響[11,15-17]。因此本文通過使用PyMOL軟件建立GTB空間模型,對700C>G突變導致的P234A置換進行分析。從圖2可以看出第1個關鍵氨基酸Gly-176距離酶活性中心較遠;Met-266和Ala-268為一對面對面的氨基酸,兩者共同形成一“瓶頸”,控制與之結合的供糖體大小及結構[18];Ser-235則出現(xiàn)在與底物結合較近的區(qū)域。圍繞P234的氨基酸有Met-266、Ser-235、His-233。野生型GTB中第234位脯氨酸屬于雜環(huán)氨基酸,圖3顯示脯氨酸中的吡咯環(huán)在空間構象上與Met-266的長側鏈非常靠近,GTB發(fā)生P234A置換后,丙氨酸的空間結構使原有的脯氨酸吡咯環(huán)位置的C-γ原子發(fā)生缺失。而脯氨酸吡咯環(huán)的C-γ原子與Met-266的側鏈通過范德華力連接,C-γ原子的改變會影響Met-266側鏈的方向[19]。因此P234A置換后,丙氨酸失去了該位置原有的C-γ原子,進而失去其與Met-266側鏈之間的范德華力,使Met-266的側鏈發(fā)生方向改變,導致Met-266和Ala-268形成的結合凹槽的空間結構發(fā)生改變,可以容納下UDP-GalNac,最終使突變的GTB能夠轉移一部分UDP-GalNac到底物H抗原,從而在紅細胞表面形成弱A抗原。此即本文提出的B(A)02等位基因導致B(A)表形的分子機制。
此外,P234在空間位置上除了和Met-266接近外,與Ser-235和His-233也十分接近。Ser-235和His-233主要影響酶-供糖體復合物與底物H抗原的結合。本例獻血者紅細胞與抗B試劑反應呈強凝集,并未減弱,說明GTB-UDPGal與H抗原的結合正常,因此推測P234A置換沒有影響Ser-235和His-233。本文沒有能夠進行酶動力學試驗,進一步考察GTB催化轉移Gal的效率,是為不足之處。
目前BGMUT數據庫已報道的B(A)02等位基因對應的氨基酸置換有5種:R176G、P234A、G235S、L266M和G268A。可以看出,除了P234A,其余4種置換均發(fā)生在4個關鍵氨基酸處,由此可見,由于P234在空間構象上距離影響酶識別供糖體特異性的Met-266非常近,因此P234的改變會造成兩者之間分子間作用力的改變,從而影響GTB對供糖體底物的識別和結合,形成B(A)亞型的紅細胞血型。
[1]Daniels G. Human blood groups[M]. 2nd ed. Oxford:Blackwell Scientific,2002:40-41.
[2]Jin S,Cai XH,Liu X,et al. Study on cisAB and B(A)subgroups in Shanghai blood donors[J]. Chin J Blood Transfusion,2013,26 (12):1198-1201.[金沙,蔡曉紅,劉曦,等.上海地區(qū)獻血人群cisAB和B(A)血型的研究[J].中國輸血雜志,2013,26(12):1198-1201].
[3]Zhang A,Lin HK,Ren BC,et al. Identification of rare B(A)blood group [J]. JClin Transfus Lab Med,2014,16(2):190-192.[張愛,林洪鏗,任本春,等.罕見B(A)亞型的鑒定[J].臨床輸血與檢驗,2014,16 (2):190-192]. doi:10.3969/j.issn.1671-2587.2014.02.02.
[4]Patnaik SK,Helmberg W,Blumenfeld OO,et al. BGMUT:NCBI dbRBC database of allelic variations of genes encoding antigens of blood group systems[J]. Nucleic Acids Res,2012,40(1):D1023-D1029. doi:10.1093/nar/gkr958.
[5]Yamamoto F,McNeill PD,Yamamoto M,et al. Molecular genetic analysis of the ABO blood group system:3. A(x)and B(A)alleles [J]. Vox Sang,1993,64(3):171- 174. doi:10.1111/j.1423-0410.1993.tb05157.x.
[6]Fung MK,Grossman BJ,Hillyer CD,et al. Technical Manual[M]. 18th ed. Bethesda:American Association of Blood Banks(AABB),2014:2-7.
[7]Cai XH,Jin S,Liu X,et al. Molecular genetic analysis for the Bx subgroup revealingtwo novel alleles in the ABO gene[J]. Transfusion, 2008,48(11):2442-2447. doi:10.1111/j.1537-2995.2008.01878.x.
[8]Yamamoto F,Clausen H,White T,et al. Molecular genetics basis of the hiso-blood group ABO system[J]. Nature,1990,345:229-233.
[9]Yazer MH,Olsson ML,Palcic MM. The cis-AB blood group phenotype:Fundamental lessons in glycobiology[J]. Transfusion Medicine Reviews,2006,20(3):207-217. doi:10.1016/j.tmrv.2006.03.002.
[10]Alfaro JA,Zheng RB,Mattias Persson M,et al. ABO(H)blood group A and B glycosyltransferases recognize substrate via specific conformational changes[J]. J Biol Chem,2008,283(15):10097-10108. doi:10.1074/jbc.M708669200.
[11]Patenaude SI,Seto NO,Borisova SN,et al. The structural basis for specificity in human ABO(H)blood group biosynthesis[J]. Nat Struct Biol,2002,9(9):685-690. doi:10.1038/nsb832.
[12]Persson M,Letts JA,Hosseini-Maaf B,et al. Structural effects of naturally occurring human blood group B galactosyltransferase mutations adjacent to the DXD motif[J]. J Biol Chem,2007,282(13):9564-9570. doi:10.1074/jbc.M610998200.
[13]WangH,ZhangX,ZhangKL,et al. l case of B(A)02 assessment report [J]. Chin JBlood Transfusion,2014,27(3):335-336.[王慧,章旭,張坤蓮,等. l例B(A)02鑒定報告[J].中國輸血雜志,2014,27(3):335-336]. doi:10.13303/j.cjbt.issn.1004-549x.2014.03.045.
[14]Hong XZ,Xue XG,Ma KR,et al. Molecular basis of the B(A)phenotype and its pedigree analysis[J]. Chin J Lab Med,2010,33(1):51-55.[洪小珍,許先國,馬開榮,等. B(A)血型分子機制研究及其家系分析[J].中華檢驗醫(yī)學雜志,2010,33(1):51-55]. doi:10.3760/cma.j.issn.1009-9158.2010.01.01.
[15]Johal AR,Blackler RJ,Alfaro JA,et al. pH-induced conformational changes in human ABO(H)blood group glycosyltransferases confirm the importance of electrostatic interactions in the formation of the semi-closed state[J]. Glycobiology,2014,24(3):237-246. doi:10.1093/glycob/cwt098.
[16]Johal AR,Schuman B,Alfaro JA,et al. Sequence-dependent effects of cryoprotectants on the active sites of the human ABO(H)blood group A and B glycosyltransferases[J]. Acta Cryst,2012,D68:268-276. doi:10.1107/S0907444912001801.
[17]Letts JA,Rose NL,F(xiàn)ang YR,et al. Differential recognition of the type I and II H antigen acceptors by the human ABO(H)blood group A and B glycosyltransferases[J]. JBiol Chem,2006,281(6):3625-3632. doi:10.1074/jbc.M507620200.
[18]Qiu L,Cai XH,Miao W,et al. Identification of a novel Bm allele and literature review[J]. Chin J Blood Transfusion,2014,27(2):143-146.[邱麗,蔡曉紅,苗溫,等. 1個新的Bm等位基因鑒定附文獻復習[J].中國輸血雜志,2014,27(2):143-146]. doi:10.13303/j.cjbt.issn.1004-549x.2014.02.0011.
[19]Marcus SL,Polakowski R,Seto NO,et al. A single point mutation reverses the donor specificity of human blood group B-synthesizing galactosyltransferase[J]. J Biol Chem,2003,278(14):12403-12405. doi:10.1074/jbc.M212002200.
(2015-09-22收稿2015-12-20修回)
(本文編輯魏杰)
Identification and molecular mechanism study of a case with B(A)02 allele
QIU Li1,YAO Nan1,MIAO Wen1,ZOU Wei2,CAI Xiaohong2
1 Binhai Tanggu Blood Station,Tianjin 300456,China;2 Rui Jin Hospital Shanghai Jiao Tong University School of Medicine Corresponding Author E-mail:cxh8407@126.com
Objective To identify and investigate B(A)02 allele in a patient. Methods Serological tests were performed with standard serological methods in a patient with B(A)02 allele. DNA sequences of all seven exons and exon -intron boundaries of ABO gene were analyzed by polymerase chain reaction(PCR),direct DNA sequencing and sequencing after gene cloning. In order to analyze the allele,PyMOL software was used to establish 3D model of Glycosyltransferases B (GTB). Results The serological results showed the characteristics of B(A)phenotype. DNA analysis revealed that ABO gene of the individual was heterozygous of B(A)02/O01 allele. 700C>G mutation was identified in B101 allele,which resulted in the amino acid substitution P234A in GTB. Through the analysis of the 3D structure of GTB,it was speculated that the P234A replacement affected the intermolecular forces of the 234 amino acid and Met- 266,thus changed the conformation of the donor-binding pocket of GTB,that made GTB capable of recognizing and tranferring the GalNac to the H antigen,which can lead to the formation of the weak A antigen on membrane of red blood cells. Conclusion The P234A replacement can affect the spatial conformation of the specific recognition region conformed by Met- 266 and Ala-268 residues,which leads to the antigenicity change of the ABO blood group.
ABO blood-group system;alleles;B(A)subgroup;PyMOL software
R457.1+1
A
10.11958/20150187
1天津市濱海新區(qū)塘沽中心血站檢驗科(郵編300456);2上海交通大學醫(yī)學院附屬瑞金醫(yī)院臨床輸血科
邱麗(1982),女,主管技師,學士,主要從事免疫血液學方面研究
E-mail:cxh8407@126.com
