以“科學思維邏輯”組織“噬菌體侵染細菌實驗”的教學建議
2016-08-21 02:38:26黃建華
生物學教學 2016年9期
關鍵詞:實驗
黃建華
(江蘇省南通大學附屬中學 226019)
1 考證“噬菌體侵染細菌實驗”的科學思維邏輯
1952年,赫爾希(A.D.Hershey)和蔡斯(M.Chase)所做“噬菌體侵染細菌的實驗”是在前人研究的基礎上完成的。
1.1 噬菌體的早期研究為同位素標記噬菌體提供了依據 英國人Twort和法國人D′Herelle分別于1915年和1917年發現一種能使細菌裂解的過濾因子,命名為噬菌體。進一步的研究發現,噬菌體是一種病毒,感染細菌后合成子代噬菌體的原料全部來自宿主細胞。另外,已知硫元素只存在于蛋白中,磷元素幾乎都存在于核酸中,這為赫爾希和和蔡斯用35S和32P分別標記噬菌體的蛋白質和核酸提供了依據。
1.2 T2噬菌體一步生長曲線的研究為噬菌體侵染細菌實驗的時間控制提供了理論依據 德國人M·Delbrück(噬菌體學派創始人)測定了烈性噬菌體T2侵染細菌和成熟噬菌體釋放的時間間隔,繪制出每個被侵染細胞釋放噬菌體的生長曲線(稱為T2噬菌體一步生長曲線),并于1939年發表“噬菌體的生長”一文。T2噬菌體的一步生長曲線表明:T2噬菌體侵染細菌的潛伏期為20 min(潛伏期是指噬菌體感染細菌至細菌釋放子代噬菌體的一段時間)。這為赫爾希和蔡斯在實驗中將噬菌體感染細菌、攪拌、離心的實驗時間控制在20 min之內提供了理論依據。
1.3 靜止噬菌體化學形態的研究推測噬菌體的增殖首先要除去保護性的外殼 美國人Anderson于1949年發現用水快速稀釋懸浮T2噬菌體的高濃度氯化鈉溶液,利用滲透壓的驟變可使噬菌體失活,在電子顯微鏡下可見這種失活的噬菌體呈蝌蚪形的“空殼”。1951年Herriott研究發現滲透壓驟變能使噬菌體的DNA釋放到溶液中,而“空殼”仍可以吸附到細菌上。這些研究推測噬菌體的增殖首先要除去保護性的外殼。
1.4 噬菌體吸附細菌后DNA對DNA酶變敏感,推測噬菌體吸附后DNA從外殼中排出 Graham和他的同事于1951年發現噬菌體吸附到加熱殺死的細菌上后,其DNA對DNA酶變敏感了;Benzer Dulbecco于1952年也發現在不引起生長的緩沖液中,噬菌體吸附到細菌上后,其DNA變得對DNA酶敏感了。據此推測噬菌體吸附到細菌上以后,其DNA就從其保護性外殼中排出,由于失去外殼的保護,故其DNA對DNA酶變敏感。
1.5 噬菌體吸附到細菌碎片上后,其DNA可從外殼中釋放出來 在前人實驗和推測的基礎之上,為了更好地驗證噬菌體吸附細菌后DNA從外殼中排出,赫爾希和蔡斯用35S和32P標記的噬菌體吸附細菌碎片(選擇吸附細菌碎片能更好的觀察到DNA釋放的結果),在37℃保溫30 min,離心15 min,然后對沉淀和上清液分別進行分析,其結果如表1[1]所示(用同位素的百分比來表示)。
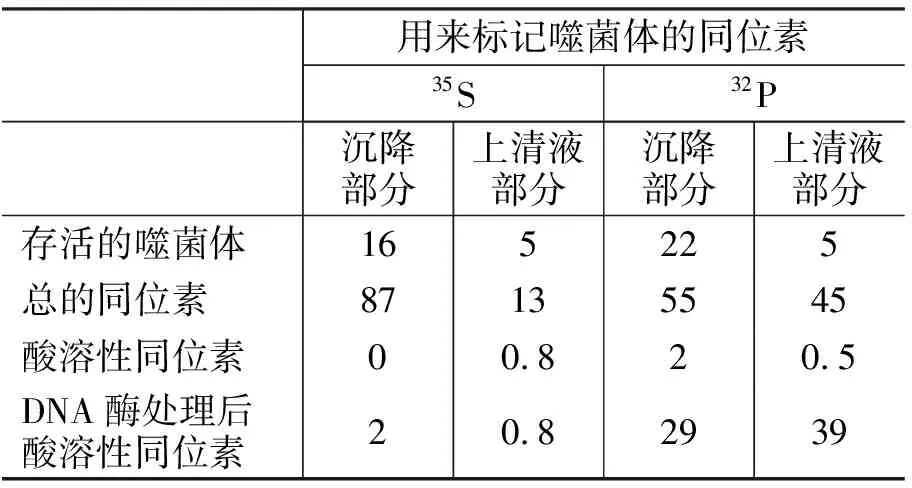
表1 赫爾希和蔡斯同位素標記實驗的結果
從35S標記的噬菌體吸附細菌碎片的結果可知:①有87%的噬菌體分布在沉降部分中,說明噬菌體的蛋白尾部吸附在細菌碎片上(因沒有搗碎攪拌),其中16%是存活的噬菌體(表明噬菌體吸附細菌后有少部分仍保持活性);②有13%的噬菌體(分布在上清液中)沒有吸附到細菌碎片上,其中5%是存活的噬菌體。
從32P標記的噬菌體吸附細菌碎片的結果可知:①有55%的總噬菌體DNA隨細菌碎片沉降到底部,其中22%是存活的噬菌體;29%對DNA酶敏感(能被DNA酶分解),說明這部分噬菌體吸附細菌后釋放出DNA并失活;酸溶性同位素占2%,說明其未釋放DNA;②有45%的總噬菌體DNA分布在上清液中,其中5%存在于存活的噬菌體中;剩下的約40% DNA基本都對DNA酶敏感(39%能被DNA酶分解)。
上述實驗數據說明,噬菌體的蛋白尾部起吸附作用;噬菌體吸附細菌后,大部分釋放DNA并失活;小部分不失活;還有些失活,但不釋放DNA。
1.6 攪拌可從受侵染的細菌上去掉噬菌體外殼 1951年,Anderson通過電子顯微鏡發現T2噬菌體是靠它的尾部吸附到細菌上去的。假定這種吸附不穩定,攪拌應該能將吸附在菌體上的噬菌體脫落下來。赫爾希和蔡斯用35S和32P標記的噬菌體在吸附培養基中侵染細菌,離心除去未吸附的噬菌體,然后將菌體放在一定的溶液(MgSO41 mM;CaCl20.1 mM;明膠 0.1 g;水 1000 mL)中,在waring搗碎器中攪拌,每隔60 s取樣,用抗噬菌體血清進行滴定,測出細菌產生的噬菌體數目,并離心測定同位素的比例及受侵染細菌的存活曲線[1](圖1)。
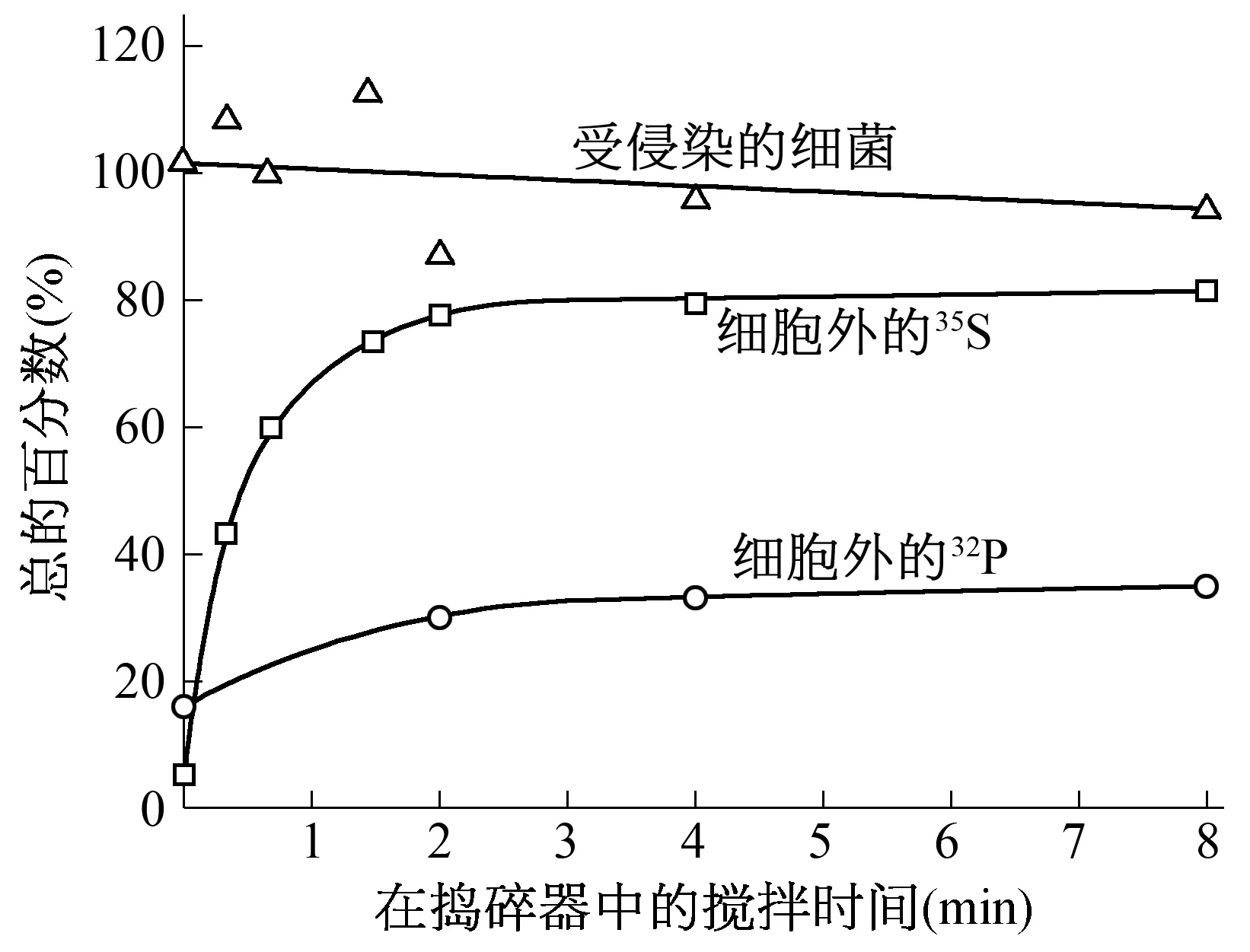
圖1 攪拌中測定的同位素比例及受侵染細菌的存活曲線
從實驗結果可知:在不攪拌的情況下,就有部分35S和32P自發脫落下來;而攪拌可以把75%左右的35S剝落下來;在這些硫釋放的同時,只有35%左右的噬菌體32P釋放出來。赫爾希和蔡斯還做了高侵染倍數實驗(增加噬菌體的感染量做重復實驗),發現高侵染倍數對32P的自發釋放影響不大,卻大大提高35S的自發釋放。這表明,噬菌體在侵染細菌的過程中,其所含的硫(蛋白質)大部分留在細菌的表面;所含的磷(DNA)大部分在吸附細菌后很快進入細胞。
1.7 硫和磷從親代噬菌體向子代噬菌體的傳遞說明含硫蛋白質在噬菌體增殖中不起作用,而DNA起作用 赫爾希和蔡斯用放射性35S標記的噬菌體侵染細菌,產生的子代噬菌體中所含的放射性,還不到親本的1%(可能是吸附在細菌表面存活的親代噬菌體中標記的硫);用32P標記的噬菌體侵染細菌,所產生的子代噬菌體所含的放射性達到親本的30%以上。1952年,Watson和Maalos研究發現:侵染性噬菌體DNA中的磷和腺嘌呤以相當大的量而且同等程度傳遞給子代噬菌體。所以赫爾希和蔡斯推斷:含硫蛋白質在噬菌體增殖中不起作用,起作用的是DNA。
1.8 赫爾希和蔡斯當時留下的討論問題 赫爾希和蔡斯在發表的論文“噬菌體生長過程中蛋白質和核酸各自的功能”中,討論部分明確提出未弄清楚的問題:除了DNA之外,噬菌體還有沒有其他種類的非硫的物質進入細胞?假如有,那么它能傳遞給子代噬菌體嗎?磷傳給子代是直接的還是間接的?[2]
從上可知,赫爾希和蔡斯在前人研究的基礎之上,設計了“噬菌體侵染細菌實驗”,逐步深入研究噬菌體生長過程中蛋白質和核酸各自的功能,其結論是蛋白質擔負著使噬菌體吸附到細菌上并使DNA注入細胞的責任,對胞內噬菌體的增殖沒有作用,而是DNA在起作用。
2 合理處理“噬菌體侵染細菌實驗”的教學建議
2.1 完善教材中的實驗步驟,尊重科學探究史實 高中生物學教材中介紹“噬菌體侵染細菌實驗”的步驟為:標記→侵染→攪拌→離心→檢測。建議根據科學史實將步驟完善為:標記→侵染→離心→培養→攪拌→離心→檢測,侵染后離心的目的是去除未吸附的噬菌體,排除未吸附的噬菌體對實驗結果的干擾。完善實驗步驟一方面尊重科學探究史實;另一方面利于實驗結果的準確分析。
2.2 準確分析實驗結果,校正現行分析偏差35S標記噬菌體侵染細菌實驗中,離心后沉淀物中也有較弱放射性原因的分析,一些資料、試題認為是攪拌不徹底,部分吸附的噬菌體外殼未脫落下來導致的。其實不是攪拌不徹底,而是在保證細菌結構穩定的前提下,攪拌不可能將吸附在細菌表面的噬菌體全部剝落下來(大約剝落75%),導致部分吸附在細菌表面的噬菌體外殼隨細菌沉降,使沉淀物中也有較弱的放射性。
32P標記噬菌體侵染細菌實驗中,離心后上清液中也有較弱放射性原因的分析,一些資料、試題認為是培養時間控制不當導致的,培養時間過短導致一些噬菌體還沒有吸附細菌,就離心到上清液中;或培養時間過長導致細菌裂解釋放的子代噬菌體處于上清液中,這樣的解釋是不科學的。赫爾希和蔡斯實驗的實際步驟是:標記→侵染→離心→培養→攪拌→離心→檢測。侵染后離心的目的就是去除未吸附的噬菌體,所以上清液中的放射性不是未吸附細菌的噬菌體攜帶的。在噬菌體一步生長曲線的基礎上,赫爾希和蔡斯嚴格控制實驗時間在20 min內。離心前細菌處于潛伏期,尚未裂解。
所以,上清液中的放射性也不是細菌裂解后釋放的子代噬菌體攜帶的,而是吸附細菌后自發脫落的完整噬菌體(未注入DNA)所攜帶的。
2.3 牢牢抓住證據力點,理清實驗遞進關系 1944年艾弗里就已證明DNA是使R型細菌轉化為S菌的轉化因子,即DNA控制細菌莢膜的形成,證明了DNA能控制生物性狀,從而證明DNA是S菌的遺傳物質。“噬菌體侵染細菌實驗”證明親代DNA能傳給子代,在“格里菲斯體外轉化實驗”的基礎上,證明了DNA在親子間的傳遞性,成為DNA是遺傳物質的更有力證據。赫爾希和蔡斯的實驗結果中,細菌裂解釋放的子代噬菌體顯示,32P標記的一組實驗中可以檢測到32P標記的DNA,35S標記的一組實驗中卻不能檢測到35S標記的蛋白質,正說明噬菌體DNA在親子代間的可傳遞性,是證明DNA是遺傳物質的更有力證據的力點,也是“噬菌體侵染細菌實驗”和“格里菲斯體外轉化實驗”呈遞進關系的承接點,分析時一定要牢牢抓住。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
