內蒙古和黑龍江的核盤菌菌絲融合群分化及致病性研究
2016-09-02 00:58:10劉春來夏吉星李新民孟慶林蘇寶華張勻華
植物保護 2016年3期
劉春來, 王 爽, 夏吉星, 楊 帆, 李新民, 孟慶林, 劉 宇, 蘇寶華, 張勻華
(黑龍江省農業科學院植物保護研究所, 哈爾濱 150086)
?
內蒙古和黑龍江的核盤菌菌絲融合群分化及致病性研究
劉春來,王爽,夏吉星,楊帆,李新民, 孟慶林,劉宇,蘇寶華,張勻華*
(黑龍江省農業科學院植物保護研究所, 哈爾濱150086)
為明確核盤菌的遺傳多樣性,對采自內蒙古和黑龍江不同地區的44株核盤菌進行了菌絲融合群確定,并比較了不同菌絲融合群間菌絲生長速度、致病力、草酸和總酸產量的差異。結果表明:供試44個菌株分為25個融合群,其中有14個融合群僅由單一菌株組成,所占比例為56.0%。菌絲融合群內和菌絲融合群間菌絲生長速度、致病力、草酸和總酸產量都表現出顯著差異(P <0.001),并與菌株的地理來源無關。相關分析表明核盤菌菌株的致病力與菌株草酸產量呈正相關(r=0.484, P ≤0.01),與pH呈負相關(r =-0.580, P ≤0.01),與菌株的生長速度無關;草酸產量與pH高低(表示總酸的分泌量)負相關(r=-0.392, P ≤0.01),進一步表明核盤菌菌株產生的總酸中草酸量占了很大的比例。
核盤菌;草酸;致病性;生長速度
核盤菌[Sclerotinia sclerotiorum (Lib.)deBary]是一種具有廣泛寄主范圍和地理分布的病原菌。它至少能夠引起75個科、278個屬和408個種及42個變種或亞種的植物發生病害,其中包括許多重要的蔬菜和經濟作物[1]。該菌的生活周期中90%是以菌核的形式在土壤中度過的,菌絲體和子囊孢子出現的時間都較為短暫,由于菌核抗逆性很強,使得核盤菌成為一種較難防治的病原菌[2]。
菌絲融合群(myceliacompatibilitygroups,MCGs) 檢測是通過真菌多位點基因控制的自我或非自我識別分類系統,是一種基于宏觀和表型的研究[3]。Kohn等鑒定了田間作物核盤菌的克隆種群并表明幾個克隆種群可侵染同一田塊[4]。MCGs是檢測克隆種群的標準方法之一,當配對培養時,同一個菌絲融合群的所有菌株能夠融合形成一個均勻一致的菌落,無反應線產生。同時,這些菌株也共有唯一的DNA指紋圖譜[3,5]。目前,菌絲親和性已被證明是研究核盤菌群體結構及遺傳多樣性的有效方法并得到了廣泛的應用[610]。
草酸(OA)被認為是核盤菌致病的一種重要的決定性因素[11]。目前,研究的熱點也主要集中于核盤菌產生草酸的量及草酸參與發病機制的作用機理:(1)降低了被侵染組織的pH水平,增強了病菌分泌的胞外酶的活性[12];(2)草酸根陰離子螯合了細胞壁Ca2+,軟化了植物細胞壁并削弱了以Ca2+為基礎的防御反應功能[12];(3)對寄主植物產生直接的毒害作用,弱化了植物,有利于病原菌入侵[13];(4)抑制了寄主植物活性氧迸發[14]。所以,草酸在核盤菌的致病過程研究中起到了非常重要的作用。
研究確定不同融合群與主要致病因子之一草酸之間的聯系,將為核盤菌病害的防治提供理論依據。本研究的主要目的是確定來自內蒙古和黑龍江不同地區的44株核盤菌的菌絲融合群,并綜合比較不同融合群間致病性差異與菌株致病力、菌絲生長速度、草酸和總酸產量間的關系,分析這些因子與致病性的相關關系。
1 材料與方法
1.1供試菌株
研究所用44株核盤菌(S.sclerotiorum)菌株主要來源于內蒙古阿榮旗孤山鎮和黑龍江省不同地區(包括哈爾濱、齊齊哈爾、牡丹江、佳木斯、大慶、綏化、黑河、雙鴨山、鶴崗等地區),分離菌株的寄主植物主要為向日葵、大豆、紅小豆和菜豆(見表1)。供試菌株均由不同地區發病植株上的菌核經75%乙醇浸泡30s,再用1‰升汞表面消毒3~5min后,經無菌水洗滌3次,轉移至PDA平板上培養3d,用接種針挑取先端菌絲至少3次,使其成為遺傳穩定菌株。
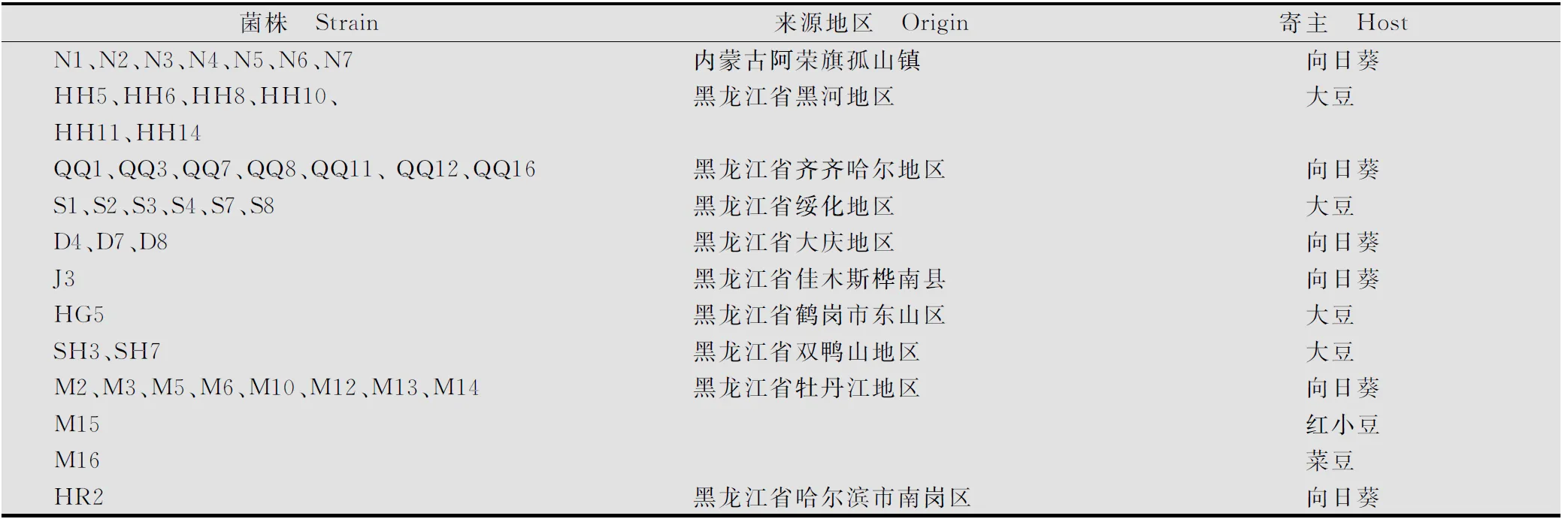
表1 來自不同地區寄主植株上的核盤菌菌株1)
1) 采自內蒙古地區的菌株來源于同一地塊。
StrainswerefromthesamefieldinInnerMongolia.
1.2核盤菌菌絲體融合群測定方法
用打孔器在培養2~3d的菌落邊緣打取生長速度一致的菌餅(d=7mm),每兩個菌株接種到同一個含50mg/L的溴酚藍(bromophenolblue,BPB)馬鈴薯葡萄糖瓊脂(PDA)平板上,兩菌餅相距5cm。其中溴酚藍的作用主要是更加清楚地觀測到菌絲不親和的反應線。作為親和組的對照,每一個菌株本身也進行了成對的比較。接種后的培養皿在23~25℃恒溫下暗培養7d。測定的原則,首先選取來自同一個地區的核盤菌菌株進行培養,測定出它們的融合群。然后,每一個融合群選取一個代表菌株分別與其他融合群的代表菌株進行親和性檢測。當兩個菌落交匯處具有一條黃色分界線或黃色到綠色的反應線說明這兩個菌株不親和,不屬于同一個菌絲融合群;而兩個菌落間沒有明顯的反應線,兩者生長融合為一體的為融合,屬于一個菌絲融合群,每次融合群測定設3個重復,并且對有疑問的親和組再進行重復驗證。
1.3核盤菌菌株生長速度測定方法
菌株經純化接入PDA平板上,待菌株培養2~3d后,于菌落邊緣打取d=7mm的菌碟,轉接至裝有PDA(含50mg/LBPB)平板的中央位置,接種后的培養皿用Parafilm膜封好,放于恒溫培養箱中 23~25℃黑暗培養48h后,測量菌落直徑,各菌株重復4次,以確定不同菌株生長速度的差異。
1.4核盤菌菌株對向日葵致病力測定
取長方形塑料盤,用75%乙醇棉擦拭盤面。在塑料盤底部鋪兩層滅過菌的吸水濾紙,倒入適量無菌水使得吸水濾紙剛好完全濕潤。取苗齡為30d左右的大小較為一致的向日葵(‘豐葵雜1號’)葉片,自來水沖洗干凈,用吸水濾紙吸干葉片表面多余水分,然后將葉片均勻地放于瓷盤內,葉片背面朝上。用打孔器在菌落的邊緣打取菌絲瓊脂塊(d=4mm),再用接菌針挑取單個菌絲瓊脂塊,準確接種在葉片主葉脈的兩側,每個菌株接種5個葉片,每個葉片接種1個菌絲瓊脂塊,以沒有接菌的瓊脂塊作為接種對照。接種后,用保鮮膜封住塑料盤,置于恒溫恒濕培養箱中培養(23~25℃)。48h后觀察發病情況,用十字交叉法量取病斑直徑大小。
1.5核盤菌菌株產草酸(OA)能力和pH測定方法
于生長一致的菌落邊緣打取1個直徑7mm的菌碟,轉至50mL三角瓶中(含30mLPDB,2%葡萄糖和0.4%馬鈴薯液,用蒸餾水配制),室溫靜止培養3d。真空抽濾到已干燥并稱重好的濾紙上,將帶有菌絲體的濾紙在鼓風干燥箱中干燥至恒重,并測得帶有菌絲體濾紙的重量,減去濾紙自身的重量得到干菌絲體的重量。上清液用于草酸和pH(用pH計)的測定。
草酸的測定參考Durman等[15]的方法,反應液包含:0.2mL的樣品液(或標準草酸溶液),0.11mL的BPB(1mmol/L),0.198mL的硫酸(1mol/L),0.176mL(100mmol/L)的重鉻酸鉀,4.8mL蒸餾水。將混合物放入60℃的水浴鍋中反應10min后,加入0.5mL(1.0mol/L)氫氧化鈉溶液終止反應。然后在分光光度計(T6新世紀紫外可見分光光度計)中測定600nm的吸光值,PDB溶液作為對照。菌株產草酸的量通過與標準曲線比較計算出,并以μg草酸/mg干菌絲來表示。各菌株總酸的產量通過pH計(賽多利斯PB-10)測定溶液的pH來表示。試驗設3次重復。
1.6數據分析
確定的25個菌絲融合群菌株的菌絲生長速度、致病力、pH及草酸產量的數據采用單一變量方差分析,最小顯著差異測定(OnewayANOVA:LSDtest)。不同菌絲融合群的菌絲體生長速度、致病力、pH及草酸產量的平均數用于分析4個因素之間的相關關系。所有試驗數據均采用SPSS12.0軟件分析完成。
2 結果與分析
2.1菌絲體融合群的確定
由表2可知,44株供試菌株共分成25個融合群。其中有14個融合群僅由單一菌株組成,所占比例為56.0%。最大的融合群為MCG3,分別由來自內蒙古,黑龍江雙鴨山、綏化3個地區的5個菌株組成;其他的融合群分別由2~4個菌株組成。通過對44株供試菌株進行菌絲融合群測定表明,采集自內蒙古和黑龍江省內的核盤菌菌株存在著豐富的遺傳多樣性,表現在同一地區內的菌株可被分成不同的融合群,同一地區同一田塊采集的菌株也被分在了不同的融合群(如內蒙古地區菌株)。

表2 菌絲融合群分組結果
2.2菌株生長速度測定結果
供試菌株在PDA(加BPB)培養基上都生長迅速。培養48h后測定菌落直徑范圍,最小為2.30cm(M13菌株),最大直徑為8.62cm(QQ7菌株)。菌株生長速度表現出顯著差異,并與菌株地理來源和菌絲融和群無關(數據未給出)。當利用菌絲融合群方法對供試菌株進行分組,培養48h后菌落平均直徑變化范圍為最小2.32cm(MCG23)到最大8.56cm(MCG11)(表3),融合群間菌絲的生長速度達極顯著差異(P<0.001)(表4)。 這種極顯著差異不僅表現在來源于不同地區融合群間,如來源于齊齊哈爾的MCG11、來源于牡丹江的MCG23和來源于綏化的MCG14,而且表現為來源于同一地區的融合群間,如來源于牡丹江的MCG20菌絲平均生長速度比MCG22和MCG23快(表2~3)。
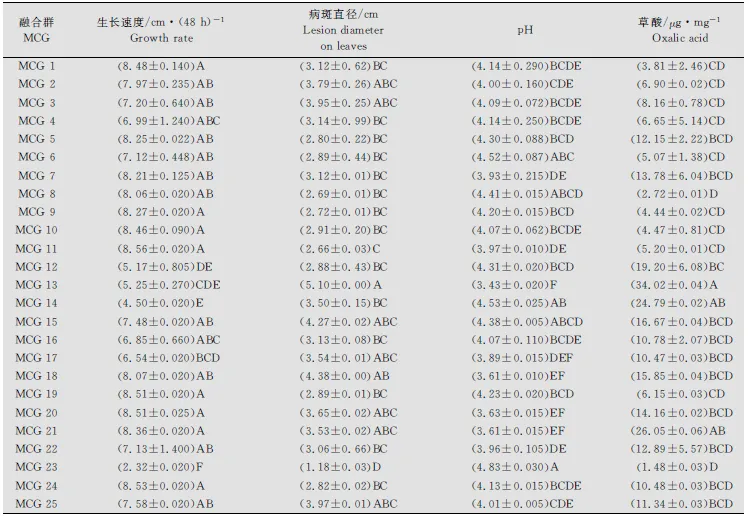
表3 核盤菌不同菌絲融合群生長速度、病斑直徑、pH及草酸產量1)
1) 按鄧肯新復極差測驗(P=0.01),同列后附不同字母的均值有極顯著差異。
DatawithinsamecolumnfollowedbydifferentletterswereextremelysignificantdifferentaccordingtoDuncan’smultiplerangetestat0.01probabilitylevel.
2.3核盤菌對向日葵品種致病力測定結果
利用菌碟貼接離體向日葵葉片,經保濕培養,12h后在葉片上即可出現明顯的水浸狀病斑,隨保濕時間的延長病斑生長迅速。48h測定病斑直徑的大小從1.20cm(M13菌株)到5.10cm(S3菌株)。從融合群間看,病斑直徑的差異達到了極顯著性水平(P<0.001)(見表4),病斑直徑變化的范圍為1.18cm(MCG23)到5.10cm(MCG13)(見表3)。不同菌株間及親合群間致病力的顯著性差異未發現與菌株的地理來源有關。
2.4核盤菌產草酸(OA)能力和pH測定結果
不同的融合群之間,菌株產草酸的量變化范圍為1.48 (MCG23)到34.02(MCG13)μg/mg干菌絲(見表3)。融合群間草酸產生能力差異很大,并達到了極顯著差異(P<0.001)(見表4)。同時,研究發現這種極顯著差異不僅表現在所劃分的所有不同融合群間,也表現在同一地區的不同融合群間,例如來源于牡丹江地區的不同融合群間,MCG21比MCG22、MCG23和MCG24產生草酸的量都多。
菌株在液體培養的過程中除了產生草酸外,還可能產生其他的未知酸類物質,致使培養液的pH產生很大的變化。測定結果顯示未接入菌碟前原始培養液(CK)的平均pH為5.03,接入菌后MCG13培養液的pH最小,達到了3.43(表3)。當以pH(表示總酸的分泌量)變化比較各融合群間總酸產量差異時,各融合群間表現出極顯著差異(P<0.001)(表4)。MCG13產生的總酸量最大(pH=3.43),而MCG23產生的總酸量最小(pH=4.83)(表3)。
2.5與致病力有關的4個因子間的相關性分析
通過對菌株生長速度、病斑直徑、草酸產量及培養液pH進行相關性分析(表5)。結果表明,菌株的致病力與菌株產生的草酸量呈正相關(r=0.484, P≤0.01),與產生的總酸(pH)之間呈負相關(r=-0.580, P ≤0.01),與菌株的生長速度無關;而生長速度與菌株產草酸量、總酸及致病力都無關;草酸產量與菌株產生總酸(pH)呈負相關(r=-0.392, P≤0.01),也就說明了總酸的產量與草酸的釋放間存在明顯的相關關系,進一步表明核盤菌菌株產生的總酸中草酸量占了很大的比例。
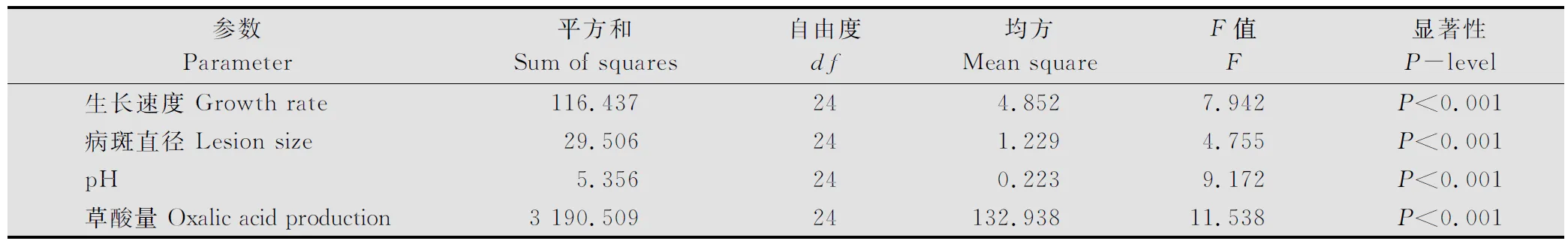
表4 核盤菌不同融合群間生長速度、病斑直徑、pH及草酸產量方差分析表
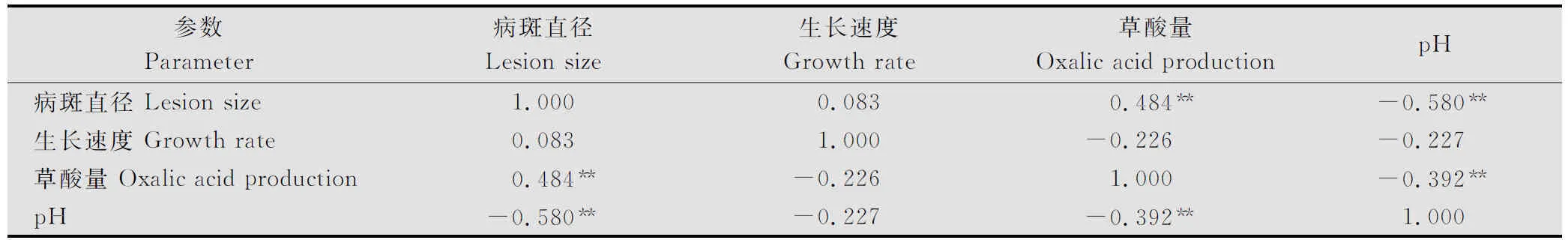
表5 菌絲體融合群間菌絲生長速度、病斑直徑、草酸產量和總酸產量的相關性分析1)
1) 表中數據為相關系數(r),**代表相關顯著性水平P≤0.01。
Datainthetablearecorrelationcoefficient(r),**indicatecorrelationissignificantat0.01level.
3 討論
本研究對采集于內蒙古一個地區和黑龍江省不同地區的核盤菌種群進行了融合群分析,44個菌株被分為25個融合群,其中有14個融合群僅由單一菌株組成,所占比例為56.0%。初步說明供試菌株存在較大的異質性,這與國內外相關研究報道的結果相符。Osofee等[9]對25個菌株進行親和性測定得到的17個菌絲融合群(MCGs)中,13個是由單個僅與自身親和的菌株組成;Li等[10]對205個菌株進行菌絲融合性測定得到的39個菌絲融合群(MCGs)中,61%是由單個僅與自身親和的菌株組成;Barari等[16]進行菌絲融合性測定得到的39個菌絲融合群(MCGs)中,26個是由單個僅與自身親和的菌株組成;李偉[17]對采集自江蘇省范圍內的145株油菜菌核病菌菌株進行了菌絲融合性測定,共鑒定出了131個菌絲融合群,其中114個是由單個僅與自身親和的菌株組成; 李沛利等[18]對四川不同生態環境和寄主來源的80個核盤菌菌株進行了菌絲融合性測定,得到63個菌絲體融合群(MCGs),其中52個菌絲融合群(MCGs)是由單個僅與自身親和的菌株組成;楊丹[19]對來自湖北省10個縣市的60個油菜核盤菌進行菌絲融合性的測定,分為27個融合群,其中12個僅由單個菌株組成,占融合群的44.4%。對于菌絲融合群與菌株的地理來源的關系,多數學者認為核盤菌的菌絲融合群與菌株的地理來源并無明顯的相關性。本研究的結果也如此,具體表現為不同地區來源的菌株可能屬于同一菌絲融合群,也可能屬于不同的菌絲融合群,同一地區來源的菌株表現相同的結果。
許多研究已經表明核盤菌存在致病性分化的現象,表現在不同融合群菌株的致病力會明顯不同,即使是同一融合群內也存在致病性明顯不同的菌株,而且這種致病力分化的現象與菌株的地理來源無關。所以,在評價某一地區的核盤菌菌株的致病性或者進行抗病育種篩選時,要盡可能多地收集菌株,才能得到更好的結果。
草酸一直以來被認為與核盤菌的致病性密切相關,作為核盤菌重要的致病因素已引起愈來愈多的關注。Godoy等[20]發現核盤菌的突變體會失去合成草酸的能力,而變為非致病菌株,當恢復突變后,菌株會重新恢復合成草酸的能力,變為正常的具有致病力的菌株。本研究證實核盤菌融合群間致病力存在顯著差異,并與產生的草酸量成正相關,與融合群的地理來源無相關性。同Durman等[15]和Li等[10]的研究結果一致。
[1]BolandGJ,HallR.IndexofplanthostsofSclerotinia sclerotiorum[J].CanadianJournalofPlantPathology,1994,16:93108.
[2]AdamsPB,AyersWA.EcologyofSclerotiniaspecies[J].Phytopathology,1979,69:896899.
[3]CarboneI,AndersonJB,KohnLM.Patternsofdescentinclonallineagesandtheirmultilocusfingerprintsareresolvedwithcombinedgenegenealogies[J].Evolution,1999,53:1121.
[4]KohnLM.Theclonaldynamicinwildandagriculturalplantpathogenpopulations[J].CanadianJournalofBotany,1994,73:S1231S1240.
[5]KohnLM,StasovskiE,CarboneI,etal.MycelialincompatibilityandmolecularmarkersidentifygeneticvariabilityinfieldpopulationsofSclerotinia sclerotiorum [J].Phytopathology,1991,81:480485.
[6]王玉杰. 向日葵菌核病病原菌遺傳多樣性及致病力分化的研究[D]. 呼和浩特:內蒙古農業大學,2011.
[7]張吉清,張學雷,肖炎,等.黑龍江省大豆菌核病菌生長特性及菌絲體親和型測定[J].中國油料作物學報,2013,35(3):307312.
[8]李易初.黑龍江大豆菌核病菌生物學特性、融合群及遺傳多樣性研究[D]. 哈爾濱:東北農業大學,2014.
[9]OsofeeH,HameedKM,MahasnedhA,etal.RelatednessamongindigenousmembersofSclerotinia sclerotiorumbymycelialcompatibilityandRAPDanalysisintheJordanValley[J].ThePlantPathologyJournal,2005,21(2):106110.
[10]LiZiqin,ZhangMin,WangYingchun,etal.MycelialcompatibilitygroupandpathogenicityvariationofSclerotinia sclerotiorumpopulationinsunflowerfromChina,CanadaandEngland[J].PlantPathologyJournal,2008,7(2):131139.
[11]DickmanMB,MitraA. Arabidopsis thalianaasamodelforstudyingSclerotinia sclerotiorumpathogenesis[J].PhysiologicalandMolecularPlantPathology,1992,41:255263.
[12]BatemanDF,BeerSV.SimultaneousproductionandsynergisticactionofoxalicacidandpolygalacturonaseduringpathogenesisbySclerotium rolfsii [J].Phytopathology,1965,58:204211.
[13]NoyesRD,HancockJG.RoleofoxalicacidintheSclerotinia sclerotiorumwiltofsunflower[J].PhysiologicalPlantPathology,1981,18(2):123132.
[14]CessnaSG,SearsVE,DickmanMB,etal.Oxalicacid,apathogenicityfatorforSclerotinia sclerotiorum,suppressestheoxidativeburstofthehostplant[J].PlantCell,2000,12:21912199.
[15]DurmanSB,MenendezAB,GodeasAM.VariationinoxalicacidproductionandmyceliacompatibilitywithinfieldpopulationsofSclerotinia sclerotiorum [J].SoilBiologyandBiochemistry,2005,37:21802184.
[16]BarariH,AlaviV,BadalyanSM.GeneticandmorphologicaldiversitiesinSclerotinia sclerotiorumisolatesinnorthernpartsofIran[J].WorldAppliedSciencesJournal,2010,8(3):326333.
[17]李偉. 江蘇省油菜菌核病菌群體遺傳結構分析[D]. 南京:南京農業大學,2007.
[18]李沛利,秦蕓,嚴吉明,等. 四川省核盤菌的營養體親和性[J]. 四川農業大學學報,2010,28(3):324327.
[19]楊丹. 湖北省油菜菌核病病菌多樣性研究[D]. 武漢:華中農業大學,2010.
[20]GodoyG,SteadmanJR,DickmanMB,etal.UseofmutantstodemonstratetheroleofoxalicacidinpathogenicityofSclerotinia sclerotiorumonPhaseolus vulgaris [J].PhysiologicalandMolecularPlantPathology, 1990,37:179191.
(責任編輯:田喆)
MycelialcompatibilitygroupandpathogenicityvariationofSclerotinia sclerotiorumpopulationsfromInnerMongoliaandHeilongjiang
LiuChunlai,WangShuang,XiaJixing,YangFan,LiXinmin,MengQinglin,LiuYu,SuBaohua,ZhangYunhua
(PlantProtectionInstituteofHeilongjiangAcademyofAgriculturalSciences,Harbin150086,China)
ToidentifythegeneticdiversityofSclerotinia sclerotiorum, 44isolatesfromInnerMongoliaandHeilongjiangwereascertainedbymycelialcompatibilitygroups(MCGs)screeningmethods,andthedifferencesamongMCGswereobservedbycomparingtheirdifferencesinmyceliagrowthrate,pathogenicityandproductionofoxalicacidandtotalacids.Theresultsshowedthatthe44isolatesweregroupedin25MCGs,amongwhich14MCGsconsistedofonlyoneisolate,withaproportionof56.0%.Significantdifferenceswerefoundinmyceliagrowthrate,pathogenicityandproductionofoxalicacidandtotalacidswithinandamongMCGs(P<0.001)regardlessoftheirgeographicorigins.CorrelationanalysisindicatedthatS. sclerotiorumpathogenicitywaspositivelyrelatedtooxalicacidproduction(r=0.484, P ≤0.01)andnegativelyrelatedtopH(r=-0.580, P≤0.01),butnotrelatedtothemyceliagrowthrate.TherewasanegativerelationshipbetweenpHvaluesandoxalicacidproduction(r=-0.392, P ≤0.01),suggestingthatoxalicacidproductioncontributedmosttothetotalproductionofacidsreleasedbyS. sclerotiorum.
Sclerotinia sclerotiorum;oxalicacid;pathogenicity;growthrate
20150602
20150817
國家現代農業產業技術體系(CARS-16)
E-mail:yhzhang9603@126.com
Q935
ADOI:10.3969/j.issn.05291542.2016.03.025
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
