黃烷酮3-羥化酶基因表達分析及轉基因大豆新種質創制
2016-09-22 08:16:53崔艷偉李文龍常文鎖李喜煥張彩英
河北農業科學 2016年3期
崔艷偉,李文龍,常文鎖,李喜煥,張彩英
(河北農業大學,教育部華北作物種質資源研究與利用重點實驗室,河北 保定 071001)
黃烷酮3-羥化酶基因表達分析及轉基因大豆新種質創制
崔艷偉,李文龍,常文鎖,李喜煥*,張彩英*
(河北農業大學,教育部華北作物種質資源研究與利用重點實驗室,河北保定071001)
黃烷酮3-羥化酶(F3H)是大豆異黃酮代謝途徑重要酶類。研究F3H在不同大豆品種中的表達差異,并應用轉基因途徑創制新種質是提高大豆異黃酮含量的重要途徑。以高異黃酮含量大豆品種中豆27和低異黃酮含量大豆品種楚秀為試材,采用qPCR技術,分析F3H在大豆不同發育時期、組織部位中的表達差異;并利用花粉管通道轉化技術,獲得轉F3H陽性植株。結果表明:F3H在2個大豆品種R1~R7期葉片中的表達模式不同,中豆27的F3H表達量在R1期最高,而后開始下降,R2~R5期維持較低水平,R6期略有上升,R7期又出現下降;楚秀F3H表達量在R1~R4期較低,R5期出現表達高峰,R6期開始下降。F3H在2個大豆品種R5~R8期籽粒中的表達模式基本相同,表達量均從R5期開始下降,且R6~R8期維持較低水平。基于此,構建F3H RNAi反義載體,并利用花粉管通道技術轉入不同大豆品種,獲得了5個轉基因新材料。
大豆;異黃酮含量;黃烷酮3-羥化酶;基因表達;RNAi
大豆異黃酮合成代謝過程由苯丙烷途徑和異黃酮合成途徑共同組成,主要存在于豆科植物[1~5]。苯丙氨酸和丙二酰輔酶A在苯丙氨酸解氨酶、肉桂酸羥化酶、香豆酸輔酶A連接酶等多種酶的催化作用下,經羥基化、甲氧基化和烷基化等過程形成不同類型的異黃酮化合物[6~9]。在這些異黃酮成分中,大豆甙元、染料木素、黃豆素為該代謝途徑主要分支產物,受到眾多學者重視,其中,染料木素的合成與黃酮及花青素合成具有共同的前體物質——柚皮素。
柚皮素為另一重要次生代謝產物,既可在異黃酮合酶(IFS)催化下合成染料木素,又可在黃烷酮-3-羥化酶(F3H)催化下合成花青素,還可在黃酮合成酶(FNS)作用下生成黃酮。由此可見,通過降低或抑制F3H、FNS表達水平,理論上可達到提高染料木素合成能力的目的[10,11]。夏濤等[12]、馬君蘭等[13]提出由于柚皮素參與多個代謝途徑,因而通過阻斷或降低其它途徑合成,可能提高大豆異黃酮含量。為分析上述途徑是否可行,少數學者進行了嘗試。Liu等[14]將大豆IFS轉入擬南芥F3H缺失體,由于分支途徑被阻斷,轉基因植株染料木素含量提高了5~30倍。Yu等[15]利用玉米C1和R轉錄因子,抑制F3H表達以阻斷花青素合成途徑,使大豆籽粒異黃酮含量升高。因此,進一步探討通過抑制分支途徑來提高異黃酮含量的可行性,對于選育高異黃酮大豆至關重要。
作者前期對213個黃淮海地區大豆的異黃酮含量進行分析,并從中篩選出了高異黃酮含量大豆品種中豆27和低異黃酮含量品種楚秀[16]。以中豆27和楚秀為試材,采用實時定量PCR技術分析F3H在2個品種不同發育時期及組織部位表達特性,明確F3H在高、低異黃酮含量大豆中的表達差異,并依此構建F3H的RNAi載體轉化大豆,獲得轉基因植株,可為高異黃酮含量新品種培育提供基礎材料。
1 材料與方法
1.1試驗材料
試驗材料為籽粒異黃酮含量差異較大的2個大豆品種,其中,中豆27為高異黃酮含量品種,楚秀為低異黃酮含量品種。
1.2試驗方法
1.2.1實時定量PCR分析取樣時期及部位2013年在河北農業大學作物育種中心試驗場種植參試大豆品。采用隨機區組試驗設計,行長2 m、行距0.5 m,2行/區,3次重復,田間管理方法同一般大田。
依據大豆生育期劃分標準[17],分別于始花期(R1)、盛花期(R2)、始莢期(R3)、盛莢期(R4)、始粒期(R5)、鼓粒期(R6)、成熟初期(R7)和完熟期(R8),選擇相同植株部位的葉片或籽粒取樣,置塑封袋后立即進行液氮冷凍,-80℃超低溫冰箱保存,備用。
1.2.2大豆樣品cDNA合成與相關引物設計參照RNA simple Total RNA Kit操作指南,進行大豆葉片和籽粒總RNA提取;參照Prime ScriptRT reagent Kit With gDNA Eraser操作指南,進行cDNA合成。
根據NCBI數據庫中檢索到的黃烷酮3-羥化酶基因F3H序列(GenBank No.AY595420),設計實時定量PCR引物,同時以組成型表達基因Actin11為內參設計引物(表1)。

表1 黃烷酮3-羥化酶基因相關引物及其序列Table 1 Primer sequences of Flavanone 3-hydroxylase gene
1.2.3F3H實時定量PCR分析采用SYBR GreenⅠ熒光染料法,參考Derveaux[18]方法進行。為保證供試基因實時定量PCR結果可靠,試驗設置3次生物學重復和3次技術重復。實時定量PCR結果分析方法為:基因在不同品種葉片相對表達量=2-△△Ct,ΔΔCt=(Ct基因-CtActin11)品種1葉片-(Ct基因-CtActin11)品種2葉片;基因在不同品種籽粒相對表達量=2-△△Ct,ΔΔCt=(Ct基因-CtActin11)品種1籽粒-(Ct基因-CtActin11)品種2籽粒。
1.2.4F3H反義載體構建設計引物F3H-F2和F3H-R2(表1),并在兩端引入SmaⅠ和PmlⅠ酶切位點序列,以高異黃酮大豆cDNA為模板進行擴增。回收PCR產物,并與克隆載體pMD18-T連接,轉化大腸桿菌感受態細胞Top10,通過菌液PCR和酶切鑒定陽性克隆,并將陽性克隆進行測序,篩選序列正確的中間重組子。將中間重組子與植物表達載體pCambia3301分別采用SmaⅠ和PmlⅠ雙酶切,回收目的片段并進行連接,連接產物轉化Top10,菌液PCR與酶切鑒定篩選陽性克隆,并通過測序分析驗證序列正確性。
1.2.5F3H RNAi載體轉化大豆及陽性植株檢測利用大豆花粉管通道遺傳轉化技術,將構建完成的F3H RNAi載體轉化不同大豆品種(東豆99-11、農大豆2號等),并做好標記。待豆莢成熟后收獲種子,然后播種于人工氣候室,利用50 mg/L除草劑篩選抗性植株,提取抗性植株DNA,經PCR擴增與DNA測序,檢測轉基因陽性植株。
2 結果與分析
2.1F3H在不同大豆品種中的表達差異
利用高異黃酮大豆中豆27和低異黃酮大豆楚秀cDNA為模板,采用實時定量引物F3H-F1和F3H-R1分別進行PCR擴增,以檢測引物擴增結果準確性,結果(圖1)顯示,能夠擴增出162 bp目的條帶,且擴增片段特異性高,沒有出現非特異條帶;進一步對擴增片段進行測序分析,發現其與NCBI注冊基因序列一致,證明上述引物擴增結果正確。

圖1 大豆F3H擴增片段瓊脂糖凝膠電泳檢測
在此基礎上,分析F3H在高、低異黃酮含量大豆葉片中的表達差異,結果(圖2)顯示,該基因在中豆27的R1期葉片表達量最高,而后表達量迅速下降,在R2~R5期保持較低水平(其中在R5期表達量最低),隨后在R6期出現第2個表達高峰,在R7期表達量又呈下降趨勢;在楚秀葉片中,F3H表達量在

圖2 F3H在不同大豆品種葉片中的表達差異Fig.2 Differential expression of F3H in different soybean varieties leaves
由此可見,由于F3H是大豆異黃酮合成過程中的一種底物競爭酶類,該基因的高效表達將會提高其競爭能力,不利于異黃酮合成與積累,故F3H在中豆27和楚秀中的表達量均較低,且由于該基因在中豆27的表達量更低,從而有利于中豆27異黃酮合成與R1~R4期較低,在R5期出現表達高峰,在R6~R7期表達量迅速下降。通過分析該基因在參試品種籽粒中的表達差異,發現F3H在2個品種籽粒中的表達趨勢基本相同,在R5~R8期均表現下降趨勢,沒有出現表達高峰(圖3)。積累。同時還發現,楚秀葉片R5期F3H表達量明顯高于其它時期,而中豆27葉片在R6期出現表達高峰,說明F3H參與底物競爭的關鍵時期發生在大豆發育的R5~R6期,且由于中豆27的該競爭反應出現得時間較晚、競爭能力較小,而楚秀的該競爭反應出現得時間較早、競爭能力相對較強,從而使中豆27異黃酮含量較高。因此,通過降低或抑制大豆品種的F3H表達,可實現提高異黃酮含量的目的。

圖3 F3H在不同大豆品種籽粒中的表達差異Fig.3 Differential expression of F3H in different soybean varieties seeds
2.2F3H RNAi載體構建
利用引物F3H-F2和F3H-R2擴增F3H相應序列,瓊脂糖凝膠電泳檢測結果顯示,目的片段大小約為1 140 bp,與預期結果基本一致;回收該目的條帶,與T載體進行連接,形成中間重組子pMD-F3H,將其轉化大腸桿菌感受態細胞,經菌液PCR擴增獲得陽性克隆;將陽性克隆進行測序分析發現,與F3H相應序列一致,可用于RNAi載體構建。
利用核酸限制性內切酶SmaⅠ和PmlⅠ,分別酶切中間重組子pMD-F3H與植物表達載體pCambia3301,獲得酶切后的目的片段;回收目的片段,并進行連接、轉化以及菌液PCR擴增與酶切鑒定,獲得重組子pCambia3301-ASF3H(圖4);將重組子進行測序分析發現序列正確無誤,表明RNAi載體pCambia3301-ASF3H構建成功。
2.3F3H RNAi載體轉化大豆及轉基因材料獲得
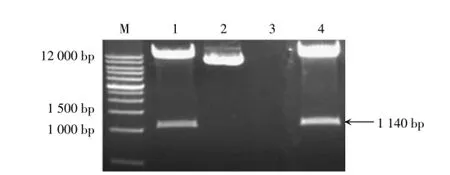
圖4RNAi載體pCambia3301-ASF3H酶切產物的鑒定
利用花粉管通道轉化技術,將RNAi載體pCambia3301-ASF3H轉入不同大豆品種,收獲轉化后的T0種子,并播種于人工氣候室;出苗后,噴施除草劑初步篩選陽性植株,野生型對照由于不抗除草劑出現葉片失綠現象,而轉基因抗性植株葉片顏色基本不變(圖5)。進一步提取抗性植株葉片DNA,進行目的基因PCR檢測,結果顯示,可以擴增出目的條帶;回收目的條帶測序證明其序列正確,說明RNAi載體pCambia3301-ASF3H已轉入大豆基因組(圖6);將陽性植株自交,收獲種子,獲得了轉F3H的T1大豆新材料5個(表2)。

圖5 利用除草劑鑒定技術檢測轉基因大豆葉片顏色變化Fig.5 Leaf colour identification results of transgenic soybean by herbicide

圖6轉基因陽性植株F3H測序結果
3 結論與討論
在大豆異黃酮合成途徑中涉及到的酶類有十幾種,目前一般采用2種策略來提高異黃酮含量[10]:第1種策略是通過分析該途徑的限速步驟,克隆或外源合成限速步驟相關酶類或基因,并導入細胞增強限速酶的表達量,打破關鍵步驟的合成瓶頸,進而提高異黃酮含量;第2種策略是通過分析異黃酮代謝途徑中的分支途徑,利用RNAi技術減少或阻塞非目標代謝物的合成,從而減少分支代謝途徑的流向,為異黃酮合成提供更加充足的合成前體[19]。其中,第1種策略研究報道較多,但由于涉及的酶類或基因數量較多,故單一基因的超表達轉化研究進展不盡人意,目前有學者提出采用“多基因”轉化的思路來提高大豆異黃酮含量[20~22]。關于第2種提高異黃酮含量的策略,目前已有學者開始嘗試并取得部分研究進展,如,張艷等[23]通過構建異黃酮代謝分支途徑F3H反義載體,抑制大豆花青素合成通路,以提高異黃酮合成能力;江曼等[24]將GhF3H RNAi載體轉入棉花,獲得了GhF3H表達水平顯著降低的棉花資源,證實通過抑制GhF3H表達水平可以改變棉花類黃酮合成;Liu等[14]研究發現,在擬南芥tt6/tt3突變體中,F3H表達受到抑制,導致該突變體異黃酮含量增加;柳青等[25]利用RNAi技術構建F3H反義載體,得到了4株轉基因植株,其異黃酮含量均表現明顯增加。作者通過分析F3H在高、低異黃酮含量大豆品種中的表達差異,同樣發現,該基因高效表達不利于異黃酮合成與積累。基于此,構建了大豆F3H的RNAi載體,并將其轉入異黃酮含量低但豐產性好、抗病的大豆品種,獲得了轉基因T1材料,為高異黃酮含量轉基因大豆新品種培育提供了新的種質資源。

表2 轉pCambia3301-ASF3H大豆新材料 (個)Table 2 Transgenic soybean germplasms with pCambia3301-ASF3H
利用高異黃酮含量大豆品種中豆27和低異黃酮含量大豆品種楚秀,分析F3H在不同大豆品種葉片和籽粒中的表達差異,證實該基因高水平表達不利于異黃酮積累。基于此,構建F3H RNAi載體并轉入低異黃酮含量大豆,獲得了轉基因材料,為進一步選育高異黃酮含量大豆品種奠定了物質基礎。Engineering,2009,11(6):355-366.
[1]Trantas Emmanouil,Panopoulos Nicklas,Ververidis Filippos.Metabolicengineeringofthecompletepathway leading to heterologous biosynthesis of various flavonoids and stilbenoids in Saccharomyces cerevisiae[J].Metabolic
[2]Richard A Dixon,Fang Chen,Dianjing Guo,Kota Parvathi.Thebiosynthesis of monolignols:a“metabolic grid”,or independent pathways to guaiacyl and syringyl units?[J].Phytochemistry,2001,57(7):1069-1084.
[3]陳宣欽,張樂,徐慧妮,陳麗梅,李昆志.大豆異黃酮生物合成關鍵酶及其代謝工程研究進展[J].中國生物工程雜志,2012,32(7):133-138.
[4]郭遠,楊柳,王園清,薛樹鵬,張嬌,崔岳,陳學珍,謝皓.大豆種子發育過程中異黃酮含量的動態分析[J].農學學報,2012,2(11):1-5,36.
[5]寧海龍,金雙義,王繼安,孫德生,李文濱,洛育,李修平,魯振明,朱秀清,孫樹坤.大豆子粒中異黃酮含量的快速測定[J].大豆科學,2004,23(1):45-49.
[6]孫君明,韓粉霞.植物次生代謝產物異黃酮的調控機理[J].西南農業學報,2005,18(5):663-667.
[7]陳封政,王雄清,李書華.大豆異黃酮的研究進展[J].樂山師范學院學報,2002,17(4):59-61.
[8]董李平.大豆異黃酮代謝機理的研究[D].重慶:西南農業大學,2003.
[9]楊俊波,楊漢奇,李德銖,李洪濤.真雙子葉基部類群十種植物的CHS基因家族成員分析[J].云南植物研究,2006,28(4):352-358.
[10]姜伊娜.大豆黃酮合成酶基因的克隆與RNA干擾調控異黃酮含量的研究[D].上海:上海交通大學,2013.
[11]Cheng H,Yang H,Zhang D,Gai JY,Yu DY.Polymorphisms of soybean isoflavone synthase and flavanone 3-hydroxylase genes are associated with soybean mosaic virus resistance[J].Molecular Breeding,2010,25(1):13-24.
[12]夏濤,高麗萍.類黃酮及茶兒茶素生物合成途徑及其調控研究進展[J].中國農業科學,2009,42(8):2899-2908.
[13]馬君蘭,趙越.外源茉莉酸甲酯(MeJA)對大豆異黃酮合成途徑的影響[J].東北農業大學學報,2011,42(5):14-18.
[14]Chang-Jun Liu,Jack W Blount,Christopher L Steele,Richard A Dixon.Bottlenecks for metabolic engineering of isoflavone glycoconjugates in Arabidopsis[J].Proceedings of the National Academy of Sciences,2002,99(22):14578-14583.
[15]Yu O,Shi J,Hession A O,Maxwell C A,McGonigle B, OdellJT.Metabolicengineeringtoincrease isoflavone biosynthesis in soybean seed[J].Phytochemistry,2003,63(7):753-763.
[16]崔艷偉,李喜煥,李文龍,常文鎖,張彩英.黃淮海大豆異黃酮含量分析與特異種質遴選[J].植物遺傳資源學報,2013,14(6):1167-1172.
[17]王連錚,郭慶元.現代中國大豆[M].北京:金盾出版社,2007.
[18]Derveaux S,Vandesompele J,Hellemans J.How to do successful gene expression analysis using real-time PCR[J].Methods,2010,50(4):227-230.
[19]姜伊娜.大豆黃烷酮-3-羥化酶基因(f3h)RNA干擾型載體的構建及功能驗證[D].上海:上海交通大學,2009.
[20]錢丹丹,龔德順,焦麗,翟瑩,張海軍,李景文,王英,王慶鈺.大豆異黃酮合成關鍵酶基因的克隆及表達分析[J].大豆科學,2011,30(5):743-748.
[21]夏循禮,楊廣笑,何光源.大豆異黃酮生物合成串聯基因簇及其表達載體的構建[J].大豆科學,2009,28(1):7-10,20.
[22]郝佳,馬會勤,代茹,陳尚武.大豆異黃酮代謝途徑在大腸桿菌中的構建及表達[J].生物工程學報,2007,23(6):1022-1028.
[23]張艷.大豆異黃酮測定方法的優化及大豆F3H基因沉默表達載體的構建[D].吉林:吉林大學,2011.
[24]江曼.下調GhDFR和GhF3H基因對棉花生長發育的影響[D].重慶:西南大學,2013.
[25]柳青,李欣達,王玉民,孫春玉,張美萍,王義. RNAi干擾的F3H基因轉化大豆的研究[J].北京農業,2010,(15):7-11.
Expression Analysis of Flavanone 3-Hydroxylase Gene and Innovation of Transgenic Elite Germplasms in Soybean
CUI Yan-wei,LI Wen-long,CHANG Wen-suo,LI Xi-huan*,ZHANG Cai-ying*
(North China Key Laboratory of Crop Germplasm Resources of Education Ministry of China,Agriculture University of Hebei,Baoding 071001,China)
Flavanone 3-hydroxylase(F3H)is an important enzyme involved in the metabolism pathway of soybean isoflavones.Analysis of the expression pattern of F3H in different soybean varieties and cultivation of the transgenic germplasms with F3H are important methods to enhance the isoflavone of soybean.In this study,the expression patterns of F3H at different developmental stages and organs in both high isoflavone content soybean variety Zhongdou 27 and low isoflavone content germplasm Chuxiu were analyzed by qPCR and transgenic elite germplasms with RNA interference vector of F3H were obtained.The results indicated that F3H showed quite different expression patterns between the two soybean varieties in the leaf.The expression level of F3H in Zhongdou 27 began falling at the R1 stage,maintained lower expression level at R2-R5 stage,increased a little bit at R6 stage and got down again at R7 stage.While in the leaf of Chuxiu,F3H showed lower expression level at R1-R4 stages,highest level at R5 stage and falling expression level at R6 stage.The expression patternsofF3Hwerealmostthesameinthe seeds of these two soybean varieties,beginning falling at stage R5 and maintaining the lower level at stages R6-R8.Based on these results,the RNA interference vector of F3H was constructed and introduced into soybean varieties through the pollentube pathway transgenic technology,and five transgenic elite germplasms were obtained by now.
Soybean;Isoflavonescontent;Flavanone 3-hydroxylase;Geneexpression;RNAinterference
S565.1
A
1008-1631(2016)02-0062-05
2015-08-11
河北省自然科學基金項目(C2013204020)
崔艷偉(1987-),女,河北衡水人,碩士,主要從事大豆分子生物學與轉基因研究。E-mail:cuiyanwei07@126. com。
李喜煥(1974-),女,河北保定人,副教授,博士,主要從事大豆遺傳育種與轉基因研究。Tel:0312-7528122;E-mail:lixihuan@hebau.edu.cn。
通訊作者:張彩英(1960-),女,河北深澤人,研究員,博士,主要從事大豆遺傳育種與轉基因研究。電話:0312-7521558;E-mail:zhangcaiying@hebau.edu.cn。
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
中國衛生(2014年12期)2014-11-12 13:12:52
