黃淮大豆主產區大豆胞囊線蟲生理小種分布調查
2016-10-19 04:14:09云王金社李海朝魏荷李金英武永康雷晨芳
作物學報 2016年10期
關鍵詞:大豆
練 云王金社李海朝魏 荷李金英武永康雷晨芳
張 輝1王樹峰1郭建秋2李月霞2李志輝3靳巧玲3徐淑霞4
張志民4楊彩云5于會勇5耿 臻6舒文濤6盧為國1,*
1河南省農業科學院經濟作物研究所 / 國家大豆改良中心鄭州分中心 / 農業部黃淮海油料作物重點實驗室, 河南鄭州 450002;2洛陽農林科學院, 河南洛陽 471023;3漯河市農業科學院, 河南漯河 462300;4安陽市農業科學院, 河南安陽 455000;5濮陽市農業科學院, 河南濮陽 457000;6周口市農業科學院, 河南周口 466001
黃淮大豆主產區大豆胞囊線蟲生理小種分布調查
練 云1,**王金社1,**李海朝1魏 荷1李金英1武永康1雷晨芳1
張 輝1王樹峰1郭建秋2李月霞2李志輝3靳巧玲3徐淑霞4
張志民4楊彩云5于會勇5耿 臻6舒文濤6盧為國1,*
1河南省農業科學院經濟作物研究所 / 國家大豆改良中心鄭州分中心 / 農業部黃淮海油料作物重點實驗室, 河南鄭州 450002;2洛陽農林科學院, 河南洛陽 471023;3漯河市農業科學院, 河南漯河 462300;4安陽市農業科學院, 河南安陽 455000;5濮陽市農業科學院, 河南濮陽 457000;6周口市農業科學院, 河南周口 466001
大豆胞囊線蟲病(SCN)在黃淮地區普遍發生, 調查病原小種分布情況, 確定優勢小種對抗病育種有重要意義。2012—2015年, 取樣調查黃淮地區6個省份土樣, 利用Riggs模式鑒定生理小種, 繪制黃淮地區SCN生理小種分布圖, 并與文獻報道結果對比, 探討黃淮地區 SCN生理小種類型及其分布規律。結果表明, 該病害在黃淮大豆主產區均有分布, 在采集受SCN感染的322份土樣中, 112份被鑒定出生理小種類型, 包括1號、2號、3號、4號、5號、6號和11號小種。其中, 57份為2號小種, 占樣本總體的50.9%; 26份土樣為5號小種, 占23.2%; 11份土樣為4號小種, 占9.8%, 1號、3號、6號和11號小種分別占總體的4.5%、5.4%、4.5%和1.8%。依據不同生理小種在各省發生頻率由高到低的順序, 河南分布5號、2號、3號、11號小種; 河北分布2號、5號、6號、3號、4號小種; 安徽分布2號、5號、6號、3號小種; 山西分布2號、4號、5號、1號、3號、11號小種; 山東分布2號、3號、5號、1號、6號小種; 江蘇分布2號、5號、1號小種。以上結果表明, 2號小種是目前黃淮地區的優勢小種, 其次是5號小種, 致病力最強的4號小種主要分布在山西省。在黃淮地區, 抗線蟲育種目標應以抗2號生理小種為主, 兼抗5號小種, 部分地區應以兼抗2號和4號小種為主。在黃淮地區3號、6號和11號小種是新發現的小種。與2001—2003年調查結果比較, 黃淮地區大豆胞囊線蟲生理小種組成及分布有一定的改變。
大豆; 大豆胞囊線蟲; 黃淮地區; 生理小種
大豆胞囊線蟲病(soybean cyst nematode, SCN)是一種世界性的大豆病害, 給大豆生產造成嚴重損失[1-2], 培育抗病品種是抵御 SCN最經濟有效的辦法。調查大豆主產區生理小種類型及分布情況, 明確育種目標, 是培育抗病品種的先決條件。
Niblack等[3]于1998—1999年, 在美國密蘇里州采取392份土壤樣品, 在183份土樣的鑒定中, 3號生理小種是優勢小種, 其次是 1號、2號生理小種,這 3個小種占所鑒定出生理小種樣本總體的 86%;Willson等[4]于 1992—1995年在美國俄亥俄州調查667塊大豆田發現, 91塊大豆田受到大豆胞囊線蟲感染, 在其鑒定的 33份樣品中發現 8個生理小種,優勢小種是3號小種, 其次是1號、2號、4號、5號、6號、10號和14號小種; 2005年, Mitchum等[5]在密蘇里州 8個生態區125個田塊采集土樣, 檢測到 49%的樣品受到大豆胞囊線蟲感染, 他們依據HG模式, 確定了其中 45個樣品的生理小種類型。上述研究均是在同一地區采集大量樣本(超過100個)來詳細描述小種的分布規律。
黃淮海流域包括黃河中下游地區、海河流域和淮河以北地區, 是中國大豆的主產區, 在我國大豆生產中占有重要地位[6-7]。何艷琴等[6]和盧為國等[8]研究表明, 大豆胞囊線蟲在河南、河北、北京、安徽、山西和山東均有分布。通過局部取樣調查發現,黃淮地區大豆胞囊線蟲生理小種分布比較復雜, 存在多個生理小種, 包括張磊等[9]在安徽蒙城鑒定的5號小種, 在安徽碭山、濉溪、亳州鑒定的4號小種;陳品三等[10]在山東半島西部的膠縣、濰坊、高密、昌邑以及河南的開封、滑縣、溫縣等地鑒定的 7號小種; 劉漢起等[11]于1983年在山東墾利、山西太谷和安徽碭山鑒定的4號小種; 李衛東等[12]、邢邯等[13]分別在在河南省鑒定的1號、2號、4號、5號小種和山東省鑒定的1號、2號、4號小種等; 李瑩等[14]在太原鑒定的 4號小種; 王衍桐等[15]在北京發現的14號小種和盧為國等[8]在河南發現的9號小種。綜上, 已鑒定的包括1號、2號、4號、5號、7號、9號、14號小種。上述研究多基于對個別省份或個別地點土樣的鑒定, 缺乏系統調查分析, 僅盧為國等[8]于2001—2003年對黃淮地區的32個土樣進行了生理小種鑒定, 并結合前人研究, 繪制了一張黃淮大豆胞囊線蟲生理小種分布圖。
與國外有關研究相比, 國內在大豆胞囊線蟲生理小種分布調查研究方面, 取樣點較為稀疏, 難以反映局部地區存在的不同大豆胞囊線蟲生理小種交叉分布的現象[8]。因此, 較為密集的采樣調查才能充分反映該地區大豆胞囊線蟲小種分布規律; 其次,對黃淮地區大豆胞囊線蟲生理小種的調查研究[8]已過去了十年之久, 在此期間, 該地區種植業結構發生了較大變化, 玉米、花生等同季作物的輪作, 是否對土壤中大豆胞囊線蟲群體結構有影響是本研究關注的另一問題。
本研究擬在前人及本課題組前期調查的基礎上,在黃淮海各省大豆種植區域密集采取土樣, 調查該地區大豆胞囊線蟲生理小種分布規律與地域分布動態, 明確優勢小種, 為SCN抗性育種提供依據。
1 材料與方法
1.1 供試材料
Riggs模式鑒別寄主[16](Lee、Peking、Pickett、PI88788和 PI90763)由河南省農業科學院經濟作物研究所大豆課題組保存, 2012年提純, 2013年在河南省農業科學院現代農業科技試驗示范基地擴繁,同年10月收獲的成熟籽粒用于本試驗。2012—2013年于大豆生長期, 在河南、河北、安徽北部、山西、山東、江蘇北部的大豆產區調查大豆胞囊線蟲分布情況, 在有大豆胞囊線蟲感染的地塊取土樣322份,每份土樣20~30 kg。
1.2 線蟲繁殖
將采集到的土樣根據土壤粘性不同, 按照病土和無菌細沙以 3.0∶0.5~1.0的比例均勻混合制備成試驗病土, 將子葉出土期(萌發約3~5 d)的Lee幼苗移栽至病土中, 每杯 1株(杯子規格為 6 cm ×12 cm)。于每年5月中旬至9月中下旬, 在河南省農業科學院現代農業科技試驗示范基地大豆胞囊線蟲培養專用塑料大棚里進行試驗。培養條件[17]為自然溫度; 每天澆水 2次, 澆水前/后土壤含水量 4%~7%/ 15%~18%; 自然濕度。約25 d后, 將大豆植株從病土中倒出, 采用淘洗-過篩法從根系中分離胞囊,用研缽輕輕研磨, 把胞囊破碎, 以看不到白色胞囊為準, 然后將釋放出來的卵沖洗入燒杯內, 用清水將蟲卵懸浮備用, 取 10 μL, 在顯微鏡下計數蟲卵密度。
1.3 接種方法
將壤土和沙子以2∶1混合后在150℃烘干2 h,裝在塑料杯中, 每45杯裝在一個67 cm ×45 cm的塑料筐中。將4個鑒別寄主及Lee在蛭石中育苗, 在子葉出土期(萌發 3~5 d), 挑選生長一致的幼苗, 移栽到塑料杯中, 每杯1株, 每個鑒別寄主播種9杯。待長出新根(移栽3~5 d)后, 接種制備好的蟲卵懸浮液, 約每杯4000個卵。接種后25 d左右、胞囊處于顯囊盛期時, 輕輕擠壓塑料杯, 待杯子中的土松動之后, 將植株拔出, 計數根部胞囊數目。
1.4 胞囊計數
利用淘洗-過篩法從根系中分離胞囊, 使其均勻分散到一塊黑布上, 用普通數碼相機照相, 拍攝從每顆植株上沖洗的胞囊照片。利用本課題自主研發的 PDS軟件, 通過計算機掃描、分析, 自動計數每張照片上的大豆胞囊數目, 具體方法見專利2014SR060158[18], 取在各寄主上生長的胞囊平均值,并根據Riggs模式判別生理小種類型。Lee上平均胞囊數目大于 60, 認定此次試驗數據有效, 否則, 將通過再次擴繁該土樣中的胞囊, 重復接種 1次, 如果Lee上平均胞囊數目仍然達不到要求, 舍棄該樣本。
1.5 數據分析及小種分布圖繪制
用“檢出頻率”表示該地區某種生理小種樣本數占該地區所鑒定出生理小種樣本總體的比例。本文定義為, 檢出頻率= (屬某一種生理小種的樣本數目/鑒定出生理小種類型的所有樣本數目)×100%。
將各采樣點鑒定出的小種類型, 利用統計分析軟件R的Maptools包[19]繪制生理小種分布圖。
2 結果與分析
2.1 生理小種的總體分布情況
在河南、河北、安徽、山西、山東和江蘇省采集大豆胞囊線蟲感染的 322份土樣, 大豆胞囊線蟲分布情況見圖1。含 210份因胞囊密度小而未鑒定出生理小種類型的樣本和 112份鑒定出生理小種類型的樣本, 鑒定出的小種類型包括1號、2號、3號、4號、5號、6號和11號, 各小種構成比例見表1。分析有生理小種的112份土樣表明, 2號生理小種最多, 檢出頻率為50.9%; 其次是5號和4號小種, 檢出頻率分別為23.2%、9.8%。其中, 3號、6號和11號小種是首次在黃淮地區發現。河北、安徽、山東發現的6號小種及在河南、山西發現的11號小種為國內初次報道。
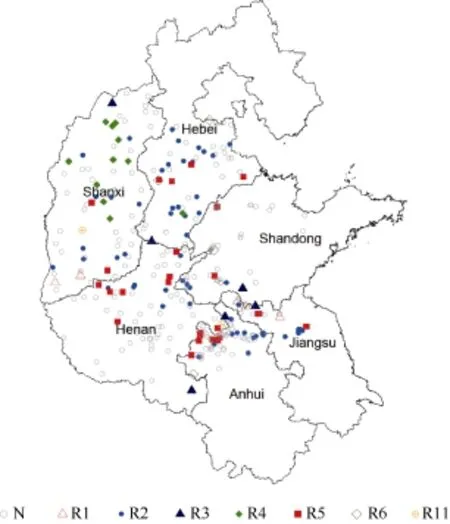
圖1 黃淮地區大豆胞囊線蟲感染地塊及生理小種分布圖Fig.1 Sampling places infected by SCN and race distribution of SCN in Huang-Huai Rivers Valley
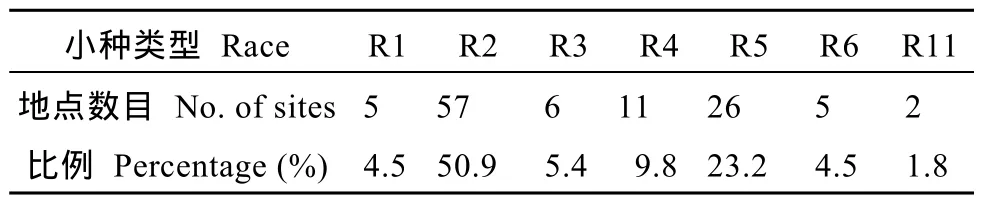
表1 黃淮地區各個生理小種所占比例Table1 Proportion of races in Huang-Huai Rivers Valley
2.2 各生理小種在黃淮大豆主產區的分布情況
從圖1可以看出, 1號生理小種主要分布在山西南部、山東和江蘇北部; 2號和5號生理小種在這6個省份均有分布; 3號生理小種分布在河南、河北、安徽、山西和山東; 4號生理小種主要分布在山西和河北; 6號生理小種主要分布在河北、安徽和山東;在河南和山西發現有11號生理小種。
2 號生理小種在黃淮海大豆主產區均有分布,集中分布在河北中部、南部, 江蘇北部和安徽省北部, 是黃淮海地區的優勢小種。
4 號生理小種集中分布在山西省中部和北部地區, 占所鑒定出4號生理小種樣本的90.9%。在黃淮海其他地區, 僅在河北省邯鄲市平鄉縣鑒定出有 4號生理小種。4號生理小種致病力最強, 抗源相對較少, 但是該小種分布集中。在山西省, 2號和4號生理小種檢出頻率分別是38.5%和38.5%。所以, 在山西省抗病育種應該以兼抗2號和4號生理小種為主要目標。
5 號生理小種在黃淮海各省均有分布。在河南省和安徽省分布廣泛, 在黃淮海地區, 5號生理小種分布范圍僅次于 2號生理小種, 是涉及黃淮海地區抗病育種工作的重點小種之一。
其他類型生理小種中, 大豆胞囊線蟲 1號生理小種主要有 3片集中分布區域, 即山西南部的運城市、山東南部的棗莊市和江蘇省西北部的徐州市沛縣和新沂市, 檢出頻率為4.5%; 3號生理小種在河南東南部、山西北部及山東、安徽、江蘇三省交界處有分布, 檢出頻率為5.4%; 6號生理小種在河北石家莊市井陘縣、河北省保定市涿州市、安徽省毫州市渦陽縣、安徽省阜陽市阜南縣、山東省泰安市東平縣有分布, 檢出頻率為4.5%; 11號生理小種在河南省永城市和山西省臨汾市霍州市有零星分布, 檢出頻率為1.8%。
2.3 黃淮地區大豆主產省份大豆胞囊線蟲生理小種構成及分布區域
由表2可見, 在河北、江蘇、安徽、山東、山西和河南, 2號生理小種檢出頻率分別是 68.00%、61.50%、52.40%、45.50%、38.50%和37.50%; 5號生理小種在以上所在省份的檢出頻率分別是16.00%、23.10%、33.30%、18.18%、7.70%和50.00%。這 2個小種所占比例之和, 在河南最高, 達到87.50%; 在最低的山西省, 也達到了46.20%。
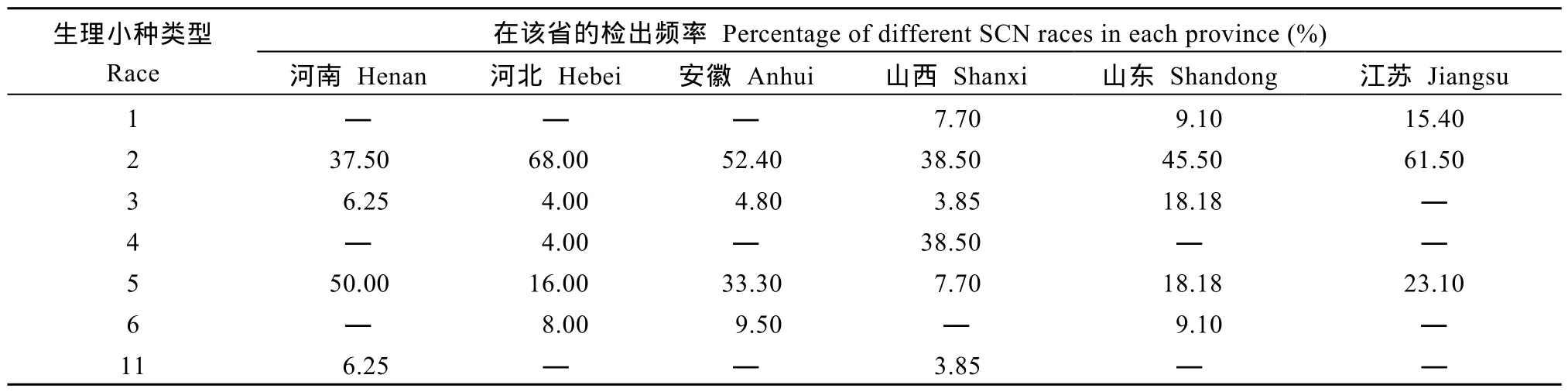
表2 黃淮地區各省大豆胞囊線蟲不同生理小種構成Table2 Proportions of SCN races in the six provinces of Huang-Huai Rivers Valley
圖2表明, 在安徽阜陽和亳州主要分布5號生理小種, 淮北和宿州一帶主要分布有2號生理小種, 部分地區零星分布有3號和6號生理小種; 河北省主要分布有 2號生理小種, 部分地區兼有 5號生理小種,零星分布有3號、4號和6號生理小種; 河南省鄭州、洛陽、濮陽、安陽一帶大豆胞囊線蟲生理小種分布較為復雜, 2號和5號生理小種交叉分布, 局部地區零星分布有3號和11號生理小種; 江蘇省淮安市、宿遷市一帶主要分布有2號生理小種, 在徐州市一帶主要分布有 5號生理小種, 部分地區兼有 1號生理小種;在山東的菏澤、濟寧、棗莊一帶主要分布有2號生理小種, 局部地區零星分布有1號、3號、5號和6號小種; 山西呂梁、太原及其以北地區主要分布有4號生理小種, 兼有2號生理小種, 山西臨汾、長治及其以南主要分布有2號生理小種, 局部地區零星分布有1號、3號、5號和11號生理小種。
表3表明, 在2001—2003年調查研究中, 1號生理小種是優勢小種, 其次是 4號生理小種; 在2012—2015年調查研究中, 2號生理小種是優勢小種, 在黃淮海大豆主產區均有分布, 檢出頻率是50.9%, 集中分布在河北中部、南部, 江蘇北部和安徽北部。
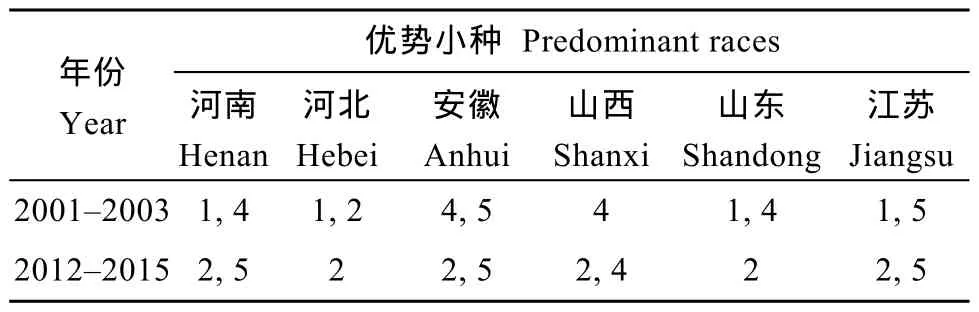
表3 不同年份黃淮地區大豆主產省份優勢小種構成的比較Table3 Composition of predominant races in the main soybean producing area of Huang-Huai Rivers Valley in different years
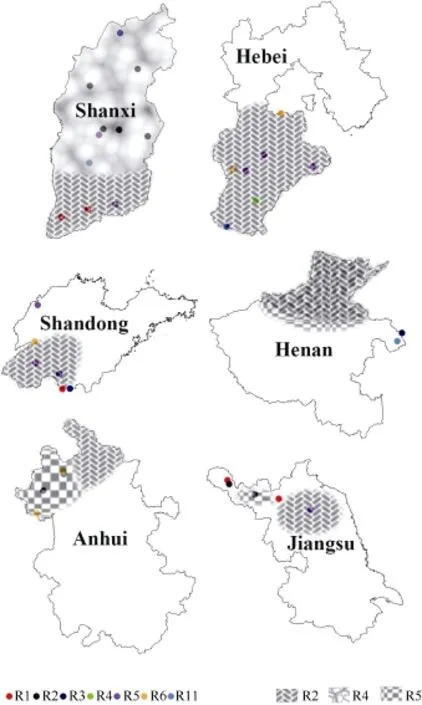
圖2 黃淮地區6省份大豆胞囊線蟲生理小種構成及主要小種分布區域Fig.2 Composition and distribution of the main SCN race in six provinces of Huang-Huai Rivers Valley
3 討論
3.1 大豆胞囊線蟲生理小種鑒別模式選擇
在對大豆胞囊線蟲生理小種類型研究過程中,前人依據致病力將SCN分為不同類型的生理小種,目前通用的鑒別方法有Riggs和Schmitt[16]于1988年建立的16個小種模式以及Niblack等[20]在2002年建立的“HG類型”, 利用7個鑒別寄主將大豆胞囊線蟲分為 128個小種。其目的均是通過鑒別寄主區分致病力不同的線蟲群體, 兩者的不同之處在于HG模式將小種劃分得更細。這兩種劃分方法各有其適宜的使用范疇, 而從抗病育種目標出發,我們認為小種不宜劃分過細, 加之生理小種存在區域間更替現象, 線蟲分布較為復雜, 劃分過細很容易給育種帶來困惑。為了便于同國內不同研究者的調查結果對比, 本研究仍然選用Riggs鑒別模式[16]探討黃淮大豆主產區大豆胞囊線蟲生理小種的分布。
3.2 黃淮地區大豆胞囊線蟲生理小種分布概況
在大豆胞囊線蟲生理小種鑒定研究中, 無論采用Riggs鑒別模式還是HG模式都需要足夠的線蟲群體接種于 4個或 7個鑒別寄主。有研究表明, 接種蟲卵數目需限定在一定范圍內, 否則鑒定結果不穩定[17]。因此, 為能繁殖出夠量的胞囊, 我們采集土樣時要求每個土樣取20~30 kg。但由于土樣采集困難, 因此國內目前已報道的研究, 多局限于對個別省份或個別地點的土樣鑒定。盧為國等[8]的調查較為系統, 但采樣點較為稀疏且沒有標識經緯度。本研究是在其基礎上, 對黃淮大豆主產區大豆胞囊線蟲生理小種分布與地域分布動態做進一步調查研究,此次共采集病土 322份, 其中 112份病土鑒定出生理小種類型, 因此, 本次調查的結果比較系統全面地反映黃淮地區大豆胞囊線蟲生理小種分布的概況,而且每份土樣均有精確的經緯度記錄, 這為今后的進一步研究奠定了基礎。從本次調查結果來看, 黃淮地區大豆胞囊線蟲發生普遍, 而且小種類型多,構成復雜。結合前人文獻報道, 目前在黃淮地區發現的小種類型有 1號、2號、3號、4號、5號、6號、7號、9號、11號、14號等[6,8-15], 2號小種為優勢小種, 3號、6號和11號小種均是首次在黃淮地區發現。
在調查中, 雖然有322個地點發現胞囊線蟲, 但僅有1/3 (112份)鑒定出小種類型。其余210份土樣是因為沒有繁殖到足夠的胞囊而未獲得可靠的結果。因此, 可以認為圖1中彩色點代表了田間胞囊量比較大的地點, 從側面反映了黃淮地區田間種群密度的概況。對比采集病土的取樣點和鑒定結果(圖1)可見, 鑒定出的生理小種集中分布在山西省、河北省和河南黃河沿岸, 這些地區屬于沙性土壤, 土質適宜大豆胞囊線蟲滋生, 因此鑒定出生理小種的樣本較多; 在黃淮區域種植大豆較為集中的安徽阜陽、安徽亳州、河南周口和河南商丘, 采集的病土較為密集, 但由于大豆胞囊線蟲種群密度較小, 因此, 鑒定出生理小種的樣本較少, 推測其原因, 主要是因為這些地區屬于砂姜黑土, 土質黏性較大,不利于大豆胞囊線蟲繁殖、擴散。
3.3 大豆胞囊線蟲生理小種的動態變化規律
在相對較長的時間周期內, 優勢生理小種構成是在變化的。從各生理小種在黃淮大豆主產區的分布看, 在2001—2003年調查研究中, 1號生理小種是優勢小種, 檢出頻率是40.0%, 集中分布在山東濟南及以東地區, 河南北部的安陽、滑縣、延津以及河北南部的邯鄲及河南漯河、周口及安徽阜陽一帶[8];在2012—2015年調查研究中, 2號生理小種是優勢小種, 在黃淮海大豆主產區均有分布, 檢出頻率是50.9%, 集中分布在河北中部、南部, 江蘇北部和安徽北部。從黃淮地區大豆主產省份生理小種構成及分布看(表3), 在2001—2003年調查研究中, 河南、山東的優勢小種均是1號和4號小種, 而在2012—2015年調查研究中, 河南的優勢小種變為2號和5號小種,山東為2號小種; 在先后2次調查研究中, 安徽、江蘇的優勢小種分別從4號、5號和1號、5號均演變為優勢小種為2號和5號小種; 河北的優勢小種由1號、2號轉變成2號小種; 山西省優勢小種由4號演變成2號和4號小種。
在一定時期內, 大豆胞囊線蟲生理小種分布是相對穩定的。本研究在2012—2015年, 選擇河南省農業科學院現代農業科技試驗示范基地連續4年監測, 鑒定結果均為2號生理小種, 此研究結果與盧為國等[8]研究結果相同。由于研究條件受限, 盧為國等[8]在2001—2003年的調查研究沒有標識經緯度, 所以很多地點無法一一對應, 但從2001—2003年和2012—2015年這兩個階段的調查研究結果看, 大豆胞囊線蟲生理小種在一定時期內是穩定的。
大豆胞囊線蟲生理小種變化的可能原因, 首先,在黃淮海生態區, 大豆的同期作物有玉米、花生和棉花等, 玉米種植面積在2014年比2002年增加313萬公頃, 而大豆種植面積同比減少50萬公頃(http://zzys.agri.gov.cn/nongqing.aspx), 這些作物的輪作及種植結構的改變, 對大豆胞囊線蟲群體結構有一定的影響; 其次, 農作物育種周期是十年, 在輪回的十年中, 各地抗性育種目標會隨著生產中遇到的病蟲害危害程度不同, 進行不同程度的調整, 這也給胞囊線蟲的生存帶來一定的選擇壓, 對抗性作物敏感的小種逐漸被淘汰, 因此大豆胞囊線蟲的群體結構為了適應新的生存環境也不斷地在變化。其變化周期和更詳盡的原因有待進一步研究, 這也是我們下一階段研究的重點。
3.4 生理小種的區域交叉分布
從上述結果看, 黃淮地區大豆胞囊線蟲生理小種構成復雜, 而且小種分布區域互相交叉。為進一步探討不同小種間交叉分布規律, 我們在較小的范圍內密集取樣, 試圖明確各小種的分布邊界。從河北石家莊市調查取樣的方圓1209 km2看(圖3), 在一定的區域內, 小種分布沒有絕對界限。我們在鹿泉市李村鎮3502農場采集到了2號土樣, 在其東南方向的藁城市堤上村和西南方向的井陘縣吳家窯鄉苗峪均采集到了5號土樣, 而在緊臨苗峪南部3.28 km但仍屬于吳家窯鄉的一塊大豆田采集到了6號土樣;而從保定市調查取樣的方圓5121 km2看, 生理小種分布在一定區域內是相對穩定的, 我們在定州市東亭鎮、保定市博野縣、保定市高陽縣南蒲口鄉北蒲村、保定市清苑縣孫村鄉大候鄉、保定市順平縣南清醒村采集到的均是2號土樣。以上研究結果表明,由于大豆胞囊線蟲是一種土傳的定居性內寄生線蟲, 其傳播速度及傳播范圍具有一定的局限性, 因此具有在局部地區優勢小種分布相對穩定、部分地區存在其他小種的特點。這與盧為國等[8]的研究結果一致。
3.5 黃淮大豆胞囊線蟲生理小種分布特點及其對育種工作的啟示
本研究結果表明, 在黃淮地區大豆胞囊線蟲分布有一定的區域性。從各省大豆胞囊線蟲分布情況看, 在河北省、江蘇省主要分布2號小種, 在河南和安徽主要分布2號和5號小種, 山西以2號和4號為主; 山東兼有2號、3號和5號小種。
國家大豆品種區域試驗在黃淮地區設3個片區[6]。黃淮海夏大豆北組包括河北寧晉、河北石家莊、山東德州、山西北部等地, 主要分布有2號生理小種,其次是4號和5號生理小種; 中組包括山東濰坊、河北邯鄲、山西南部、河南濮陽、河南洛陽等地, 主要分布有2號和5號生理小種, 兼1號和6號生理小種;南組包括江蘇北部、安徽北部、河南鄭州、河南駐馬店、河南周口、山東濟寧、山東菏澤等地, 主要分布2號生理小種, 其次是5號生理小種, 兼1號和6號生理小種。整個黃淮地區以抗2號小種為主要育種目標, 北組需要兼抗4號小種, 中組和南組需要兼抗5號小種。
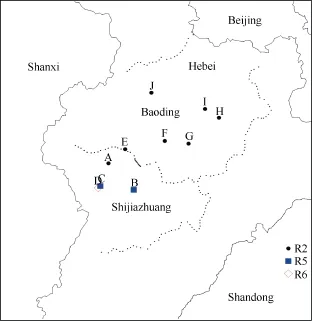
圖3 河北省大豆胞囊線蟲生理小種分布圖Fig.3 Distribution of SCN race in Hebei province
SCN的抗性是受多個位點控制的數量性狀, Rhg位點之間相互作用[21-26], 多篇文獻報道[27-28]: 大豆的第18染色體(G連鎖群)上的rhg1位點與抗病基因緊密連鎖, 控制著對多個大豆胞囊線蟲生理小種的抗性, 因此培育多抗品種是可能的。鑒于生理小種的變化, 在抗線蟲育種中, 我們更需要培育兼抗2個或2個以上生理小種的大豆品種。
4 結論
黃淮海地區大豆胞囊線蟲分布廣泛, 約分布于70%的地區。黃淮地區存在1號、2號、3號、4號、5號、6號和11號生理小種, 其中 2號小種為該地區的優勢小種, 其次是5號生理小種, 而4號生理小種主要集中分布在山西, 3號、6號和11號小種為新發現的小種。大豆胞囊線蟲在多數地區的分布呈連續狀, 組成較為穩定, 有區域性分布的特點, 但在少數的局部地區沒有明顯規律, 在夾縫區域有其他小種存在的特點。整個黃淮海地區大豆育種的目標應以抗2號生理小種為主, 部分地區應以兼抗2號和5號生理小種為主, 而在山西應以兼抗2號和抗4號生理小種為主。
致謝: 本文在調查方案制定、論文寫作等方面得到南京農業大學蓋鈞鎰院士的悉心指導, 在此深表謝意。
References
[1] Wrather J A, Koenning S R.Estimates of disease effects on soybean yields in the United States 2003 to 2005.J Nematol, 2006,38: 173-180
[2] Niblack T L, Tylka G L, Riggs R D.Nematode pathogens of soybean.Wilcox J R.Soybeans: Improvement, Production, and Uses.Madison, WI, USA: American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America,2004.pp 821-851
[3] Niblack T L, Wrather J A, Heinz R D, Donald P A.Distribution and virulence phenotypes of Heterodera glycines in Missouri.Plant Dis, 2003, 87: 929-932
[4] Willson H R, Riedel R M, Eisley J B, Young C E, Jasinski J R,Wheeler T A, Kauffman P H, Pierson P E, Stuart M C.Distribution and virulence phenotypes of Heterodera glycines in Missouri.J Nematol, 1996, 28: 599-603
[5] Mitchum M G, Wrather J A, Heinz R D, Shannon J G, Danekas G.Variability in distribution and virulence phenotypes of Heterodera glycinesin Missouri during 2005.Plant Dis, 2007, 91:1473-1476
[6] 何艷琴, 閆曉燕, 吳存祥, 楊中路, 年海.中國大豆新品種動態.北京: 中國農業科學技術出版社, 2013 He Y Q, Yan X Y, Wu C X, Yang Z L, Nian H.The dynamic of new soybean variety in China.Beijing: China Agricultural Science and Technology Press, 2013
[7] 練云, 盧為國.大豆抗 SCN機制及抗病相關基因研究進展.中國油料作物學報, 2013, 35: 727-732 Lian Y, Lu W G.Advances on resistance mechanism and gene to SCN in soybean.Chin J Oil Crop Sci, 2013, 35: 727-732 (in Chinese with English abstract)
[8] 盧為國, 蓋鈞鎰, 李衛東.黃淮地區大豆胞囊線蟲(Heterodera glycines Ichinohe)生理小種的抽樣調查與研究.中國農業科學, 2006, 39: 306-312 Lu W G, Gai J Y, Li W D.Sampling survey and identification of races of soybean cyst nematode (Heterodera glycines Ichinohe) in Huang-Huai Valleys.Sci Agric Sin, 2006, 39:306-312 (in Chinese with English abstract)
[9] 張磊.安徽淮北地區大豆胞囊線蟲生理小種研究初報.大豆科學, 1988, 7: 251-254 Zhang L.Preliminary research on races of soybean cyst nematode (Heterodera glycines Ichinohe) in Huaibei area of Anhui province.Soybean Sci, 1988, 7: 251-254
[10] 陳品三, 張東生, 陳森玉.大豆胞囊線蟲7號生理小種的研究初報.中國農業科學, 1987, 20(2): 94 Chen P S, Zhang D S, Chen S Y.First report on a new physiological race (race 7) of soybean cyst nematode (Heterodera glycines).Sci Agric Sin, 1987, 20(2): 94 (in Chinese with English abstract)
[11] 劉漢起, 商邵剛, 甄鴻杰, 霍虹, 吳和禮, 姚振純, 李秀蘭.大豆胞囊線蟲生理小種研究初報.大豆科學, 1985, 4:131-136 Liu H Q, Shang S G, Zhen H J, Huo H, Wu H L, Yao Z C, Li X L.Study on physiological races of soybean cyst nematode (Heterodera glycines).Soybean Sci, 1985, 4: 131-136 (in Chinese with English abstract)
[12] 李衛東, 賀春林, 田保明.河南省大豆胞囊線蟲分布及生理小種鑒定.華北農學報, 1991, 6(增刊1): 111-114 Li W D, He C L, Tian B M.Sampling survey and identification of races of soybean cystnematode (Heterodera glycines Ichinohe) in the province of Henan.Acta Agric Boreali-Sin,1991, 6(S1): 111-114 (in Chinese with English abstract)
[13] 邢邯, 趙經榮, 戰明奎, 李佩廷, 蓋鈞鎰.山東省大豆胞囊線蟲生理小種的鑒定.中國油料作物學報, 1997, 19(4): 61-65 Xing H, Zhao J R, Zhan M K, Li P T, Gai J Y.Identification of races of soybean cystnematode (Heterodera glycines Ichinohe)from Shandong province.Oil Crops China, 1997, 19(4): 61-65 (in Chinese with English abstract)
[14] 李瑩, 王志, 焦廣音, 常汝鎮.中國大豆遺傳資源對大豆胞囊線蟲 4號生理小種的抗性鑒定研究.中國農業科學, 1991,24(5): 64-69 Li Y, Wang Z, Jiao G Y, Chang R Z.Studies on resistance of soybean germplasm resources to race 4 of soybean cyst nematode.Sci Agric Sin, 1991, 24(5): 64-69 (in Chinese with English abstract)
[15] 王衍桐, 彭德良, 陳受宜.灰布支黑豆對大豆孢囊線蟲(Heterodera glycines) 14號小種的抗性遺傳.遺傳學報, 2000,27: 146-150 Wang Y T, Peng D L, Chen S Y.Inheritance of resistance to Heterodera glycines race 14 in Huibuzhi black bean.Acta Genet Sin, 2000, 27: 146-150 (in Chinese with English abstract)
[16] Riggs R D, Schmitt D P.Complete characterization of the race scheme for Heterodera glycines.J Nemato, 1988, 20: 392-395
[17] 練云, 魏荷, 王金社, 雷晨芳, 李海朝, 武永康, 盧為國.影響大豆胞囊線蟲生理小種鑒定因素探討.分子植物育種,2015, 13: 1259-1264 Lian Y, Wei H, Wang J S, Lei C F, Li H C, Wu Y K, Lu W G.A study on the factors that influence the race-identification of soybean cyst nematode.Mol Plant Breed, 2015, 13:1259-1264 (in Chinese with English abstract)
[18] 王金社, 盧為國, 李金英, 練云, 魏荷, 李海朝, 雷晨芳.植物病蟲害表型數據采集系統.專利號: 2014SR060158,中國, 2014 Wang J S, Lu W G, Li J Y, Lian Y, Wei H, Li H C, Lei C F.The data acquisition system on the phenotype of plant diseases and insect pests.Patent number: 2014SR060158,China, 2014
[19] Team R C.R: a language and environment for statistical computing.R Found Statist Comput, 2014
[20] Niblack T L, Arelli P R, Noel G R, Opperman C H, Orf J H,Schmitt D P, Shannon J G, Tylka G L.A revised classification scheme for genetically diverse populations of Heterodera glycines.J Nematol, 2002, 34: 279-288
[21] Afzal A J, Srour A, Saini N, Hemmati N, El Shemy H A,Lightfoot D A.Recombination suppression at the dominant Rhg1/Rfs2 locus underlying soybean resistance to the cyst nematode.Theor Appl Genet, 2011, 124: 1027-1039
[22] Yuan C P, Li Y H, Liu Z X, Guan R X, Chang R Z, Qiu L J.DNA sequence polymorphism of the Rhg4 candidate gene conferring resistance to soybean cyst nematode in Chinese domesticated and wild soybeans.Mol Breed, 2012, 30: 1155-1162
[23] Glover K D, Wang D, Arelli P R, Carlson S R, Cianzio S R,Diers B W.Near isogenic lines confirm a soybean cyst nematode resistance gene from PI 88788 on linkage group.J Crop Sci, 2004, 44: 936-941
[24] Vuong T D, Sleper D A, Shannon J G, Nguyen H T.Novel quantitative trait loci for broad-based resistance to soybean cyst nematode (Heterodera glycines Ichinohe) in soybean PI 567516C.Thoer Appl Genet, 2010, 121: 1253-1266
[25] Liu X, Liu S, Jamai A, Bendahmane A, Lightfoot D A,Mitchum M G, Meksem K.Soybean cyst nematode resistance in soybean is independent of the Rhg4 locus LRR-RLK gene.Funct Integr Genomics, 2011, 11: 539-549
[26] 盧為國, 蓋鈞鎰, 鄭永戰, 李衛東.大豆遺傳圖譜的構建和抗胞囊線蟲(Heterodera glycines Ichinohe)的QTL分析.作物學報, 2006, 32: 1272-1279 Lu W G, Gai J Y, Zheng Y Z, Li W D.Construction of a soybean genetic linkage map and mapping QTLs resistant to soybean cyst nematode (Heterodera glycines Ichinohe).Acta Agron Sin, 2006,32: 1272-1279 (in Chinese with English abstract)
[27] Concibido V C, Diers B W, Arelli P R.A decade of QTL mapping for cyst nematode resistance in soybean.Crop Sci,2004, 44: 1121-1131
[28] Afzal A J, Natarajan A, Saini N, Iqbal M J, Geisler M, El Shemy H A, Mungur R, Willmitzer L, Lightfoot D A.The nematode resistance allele at the rhg1 locus alters the proteome and primary metabolism of soybean roots.Plant Physiol, 2009, 151: 1264-1280
Race Distribution of Soybean Cyst Nematode in the Main Soybean Producing Area of Huang-Huai Rivers Valley
LIAN Yun1,**, WANG Jin-She1,**, LI Hai-Chao1, WEI He1, LI Jin-Ying1, WU Yong-Kang1, LEI Chen-Fang1,ZHANG Hui1, WANG Shu-Feng1, GUO Jian-Qiu2, LI Yue-Xia2, LI Zhi-Hui3, JIN Qiao-Ling3, XU Shu-Xia4,ZHANG Zhi-Min4, YANG Cai-Yun5, YU Hui-Yong5, GENG Zhen6, SHU Wen-Tao6, and LU Wei-Guo1,*
1Zhengzhou Subcenter of National Soybean Improvement Center / Key Laboratory of Oil Crops in Huang-Huai Valleys of Ministry of Agriculture / Institute of Industrial Crops, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China;2Luoyang Academy of Agriculture and Forestry Sciences, Luoyang 471023, China;3Luohe Academy of Agricultural Sciences, Luohe 462300, China;4Anyang Academy of Agricultural Sciences,Anyang 455000, China;5Puyang Academy of Agricultural Sciences, Puyang 457000, China;6Zhoukou Academy of Agricultural Sciences, Zhoukou 466001, China
Soybean cyst nematode (SCN) disease, caused by Heterodera glycines, occurs widely in the Huang-Huai Rivers Valley.Resistant cultivar is the most important solution to ocntrol this disease and reduce yield loss.Knowledge of SCN race distribution in soybean producing area is basis for SCN resistant breeding.A four-year survey from 2012 to 2015 was carried out to understand the diversity and distribution of Heterodera glycines races in the Huang-Huai Rivers Valley.A distribution map of races was constructed based on Riggs model.A total of 322 soil samples with SCN infection were collected.Seven H.glycines races were identified in 112 (34.8%) samples from six provinces, including five races described previously.Race 2 was the leading race, accounting for 50.9% in the 112 soil samples; race 5 was next to race 2 with the ratio of 23.2%; and the remaining types were races 4, 3, 1, 6, and 11, accounting for 9.8%, 5.4%, 4.5%, 4.5%, and 1.8%, respectively.The distribution of races varied greatly among the six provinces.According to frequencies from high to low, they were races 5, 2, 3, and 11 in Henan, races 2, 5, 6, 3, and 4 in Hebei, races 2, 5, 6, and 3 in Anhui, races 2, 4, 5, 1, 3, and 11 in Shanxi, races 2, 3, 5, 1, and 6 in Shandong, and races 2, 5, and 1 in Jiangsu.These results indicate that race 2 is the predominant race in the Huang-Huai Rivers Valley, followed by race 5, and the most virulent race 4 is centered in Shanxi province.Therefore, the SCN resistant breeding in this region should be mainly targeted towards race 2 with consideration of race 5 simultaneously, and race 4 also needs attention in some areas.In this study, races 3, 6,and 11 were newly detected in the Huang-Huai Rivers Valley.We also found that the race components and their distributions varied compared to the survey conducted in the same region in 2001-2003.
Soybean; Soybean cyst nematode; Huang-Huai Rivers Valley; Race
10.3724/SP.J.1006.2016.01479
本研究由國家自然科學基金項目(31371652), 河南省農業科學院科研發展專項資金項目(20137905), 國家現代農業產業技術體系建設專項(CARS-004)和河南省科技攻關項目(162102110141)資助。
This study was supported by the National Natural Science Foundation of China (31371652), the Special Fund for Scientific Research and Development of Henan Academy of Agricultural Sciences (20137905), the China Agriculture Research System (CARS-004), and the Science and Technology Project of Henan Province (162102110141).
(Corresponding author): 盧為國, E-mail: 123bean@163.com**同等貢獻(Contributed equally to this work)
聯系方式: 練云, E-mail: lianyun262@126.com; 王金社, E-mail: wjs33314@126.com
Received(): 2016-03-01; Accepted(接受日期): 2016-06-20; Published online(網絡出版日期): 2016-07-04.
URL: http://www.cnki.net/kcms/detail/11.1809.S.20160704.0826.008.html
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
