牛糞堆肥中反硝化細菌與理化參數的關系
2016-10-21 05:06:15朱麗平許修宏張文浩成利軍李洪濤王紫琪
農業環境科學學報 2016年9期
關鍵詞:結構
朱麗平,許修宏,張文浩,成利軍,李洪濤,王紫琪,賀 寧
(東北農業大學資源與環境學院,哈爾濱 150030)
牛糞堆肥中反硝化細菌與理化參數的關系
朱麗平,許修宏*,張文浩,成利軍,李洪濤,王紫琪,賀寧
(東北農業大學資源與環境學院,哈爾濱 150030)
以nosZ基因為標記,應用聚合酶鏈式反應-變性梯度凝膠電泳法(PCR-DGGE)對牛糞堆肥過程中反硝化細菌群落結構進行研究,通過冗余分析(Redundancy analysis,RDA)法,分析反硝化細菌群落結構變化與堆肥過程中理化參數(堆體溫度、pH、含水率、氨態氮和硝態氮)之間的關系。結果表明,反硝化細菌群落的Shannon-Weaver指數在堆肥0~4 d升高,在7~29 d減小。對優勢條帶進行測序發現Alcaligenes存在于整個堆肥過程中,判定其為此過程中優勢菌屬。反硝化細菌群落結構在堆肥前期(0~4 d)變化較小,而在降溫腐熟期(23~29 d)變化劇烈。皮爾遜相關系數分析表明,反硝化細菌群落多樣性與含水率呈極顯著正相關關系(r= 0.960,P<0.01),與溫度呈顯著正相關關系(r=0.766,P<0.05)。
堆肥;反硝化細菌;理化參數;群落結構;冗余分析
朱麗平,許修宏,張文浩,等.牛糞堆肥中反硝化細菌與理化參數的關系[J].農業環境科學學報,2016,35(9):1781-1787.
ZHU Li-ping,XU Xiu-hong,ZHANG Wen-hao,et al.Correlationship between denitrifying bacteria and physicochemical factors in cow manure composting[J]. Journal of Agro-Environment Science,2016,35(9):1781-1787.
近年來,隨著我國農業和畜牧業的迅速發展,大量畜禽糞便和農業秸稈露天放置,對環境造成了嚴重污染[1]。目前,采取了多種方式對禽畜糞便和農作物秸稈進行資源化處理,如將禽畜糞便加工成有機肥料、將秸稈進行飼料化或材料化等[2],但好氧堆肥法仍然是處理有機固體廢棄物最有效的方式之一[3]。
在堆肥化過程中,反硝化細菌能夠將硝酸鹽轉化成氣態氮(NO、N2O、N2)[4],不僅會造成空氣污染,同時還降低了堆肥品質。參與反硝化過程的還原酶主要有4種[5],其中的nosZ基因催化N2O還原成N2[6],能夠有效減少溫室氣體的排放,因此,在反硝化過程中,以nosZ基因為標記,分析反硝化細菌的群落結構變化,對研究堆肥過程中氮素轉化過程及規律具有重要的意義。近些年來,雖然在研究堆肥化過程中的反硝化菌方面取得了很大的進展[2,7],但對堆肥過程中理化參數與反硝化菌群之間關系的研究尚少。
本研究采用牛糞與稻草秸稈進行好氧堆肥實驗,對堆肥過程中溫度(T)、pH、含水率、氨態氮(NH+4-N)、硝態氮(-N)等理化參數進行參數測定。通過聚合酶鏈式反應-變性梯度凝膠電泳技術(PCRDGGE)來研究以nosZ基因為標記的反硝化細菌的群落動態變化,應用冗余分析(Redundancyanalysis,RDA)方法研究堆肥過程中反硝化細菌群落結構與理化參數之間的相關性,利用皮爾遜相關系數法分析反硝化細菌群落多樣性與理化參數的相關性,并分析影響菌群變化的驅動因子。
1 材料與方法
1.1堆肥實驗與樣品采集
堆肥實驗中的牛糞與水稻秸稈均來自哈爾濱市農場,堆肥材料主要成分見表1。將水稻秸稈剪成小段(3~5 cm),便于與牛糞的充分混勻,秸稈與牛糞按照干重比1∶3進行充分混合,并且將混合物料的含水率調節為60%~65%,置于實驗室自制的堆肥模擬容器中進行發酵,堆肥過程中定期進行翻堆,整個堆肥過程共持續30 d。
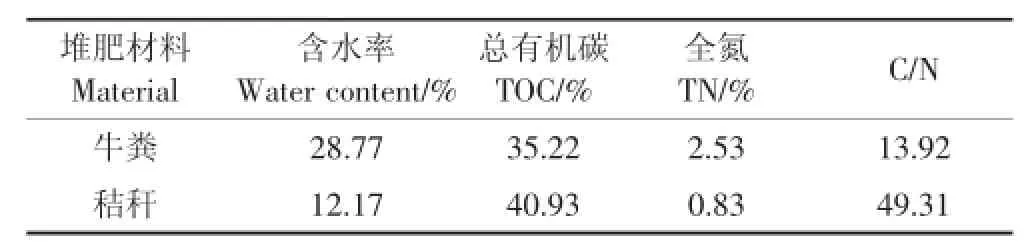
表1 堆肥材料的主要成分Table 1 Properties of raw materials for composting
在堆體的上中下三層的不同位置分別在第0、1、4、7、13、23、29 d取樣并混勻(7個樣品),以堆肥天數為樣品標號。在堆肥第3、5、11、21 d進行翻堆。7個樣品分為兩部分保存,一部分用于理化參數的測定,另一部分放于-80℃用于分子實驗分析。
1.2堆肥理化參數的測定
在堆體上、中、下三層的不同位置,使用精密溫度計分別測其溫度并取平均值,同時記錄下室溫。堆體含水率的測定采用恒重法,將7個堆肥樣品在105℃下烘干24 h,然后進行計算,得出含水率數值。pH的測定是將每個堆肥樣品稱取1 g,按1∶10(質量濃度)加入去離子水,150 r·min-1下振蕩30 min后在10 000 r·min-1進行離心,離心15 min后過濾,濾液用數字pH儀器測定。NH+4-N含量根據鮑士旦[8]的農化分析方法進行測定,-N含量采用酚二磺酸比色法進行測定[9]。
1.3DNA提取、純化
用實驗室改進的CTAB法對堆肥樣品總DNA進行提取[10],使用OMEGA純化試劑盒對粗提取的總DNA進行回收、純化,檢測純化結果后于-20℃保存備用。
1.4PCR擴增
堆肥過程中反硝化細菌的檢測以nosZ基因為標記,選用引物對nosZ-F/nosZ-1622R進行PCR擴增,并且在nosZ-1622R的5′端添加GC夾[11]。PCR擴增,體系為50 μL,其中:10×PCR Buffer 5 μL,dNTP 5 μL,模板1 μL,Taq酶0.6 μL,上下游引物各 0.8 μL,ddH2O補足至50 μL。擴增條件為:預變性5 min(95℃);35個循環中再次變性45 s(95℃),退火45 s(61℃),延伸1 min(72℃);最后再延伸10 min(72℃)[12],于4℃下保存并停止反應。
1.5DGGE及條帶測序
應用DcodeTM系統對PCR產物進行DGGE實驗。使用尿素濃度范圍為35%~70%的8%聚丙烯酰胺凝膠(丙烯酰胺∶雙丙烯酰胺=37.5∶1,m/V)。電泳條件:在60℃恒溫的1×TAE電泳緩沖液中,120 V恒定電壓下電泳12 h[11]。電泳結束后,將DGGE膠進行染色并拍照。切下主要條帶溶于水后,用不帶GC夾子的引物進行PCR擴增,并測序。
測序結果提交至GenBank獲得基因登錄號,用BLAST比對出同源性最高的序列并下載,使用MEGA4.0程序的鄰接法(Neighbor-Joining)構建系統進化樹。
1.6數據處理
DGGE圖譜分析采用Quantity One 4.6.2軟件,根據公式H=-ΣPiln Pi計算出反硝化細菌的多樣性指數(H);反硝化細菌群落結構變化與理化參數的相關性分析采用Canoco 4.5軟件分析,首先對分析數據進行除趨勢對應分析(Detrended correspondence analysis,DCA),當排序軸的梯度長度相對較短時(<2),選用冗余分析方法進行分析[13],應用Canodraw 4.5程序將生成的數據文件進行作圖分析[14];利用SPSS17.0軟件中皮爾遜相關系數法分析反硝化細菌群落多樣性與理化參數之間的相關性。
2 結果與討論
2.1理化參數的變化
圖1為堆肥過程中溫度變化,根據堆肥過程中溫度的變化可以將堆肥過程分為三個階段:0~3 d為升溫期;4~12d為高溫期;13~29d為降溫腐熟期。堆肥開始后溫度逐漸升高,在第5 d時達到最高溫57.6℃,并且溫度超過55℃的堆肥時間持續了5 d,可在堆肥過程中殺死各種病原菌,滿足基本的堆肥衛生標準[15]。堆體溫度在堆肥第13 d開始迅速下降,且在堆肥后期(13~29 d)溫度變化趨于平穩,堆肥過程結束時溫度接近于室溫。堆肥過程中溫度的變化與堆體內微生物活性息息相關,二者相互影響,微生物的大量活動導致堆體溫度升高,同時高溫也會影響微生物體內的酶活。所以,溫度既是影響堆肥過程中微生物群落變化的重要因素,也是評價堆肥是否達到穩定化的重要條件[16]。
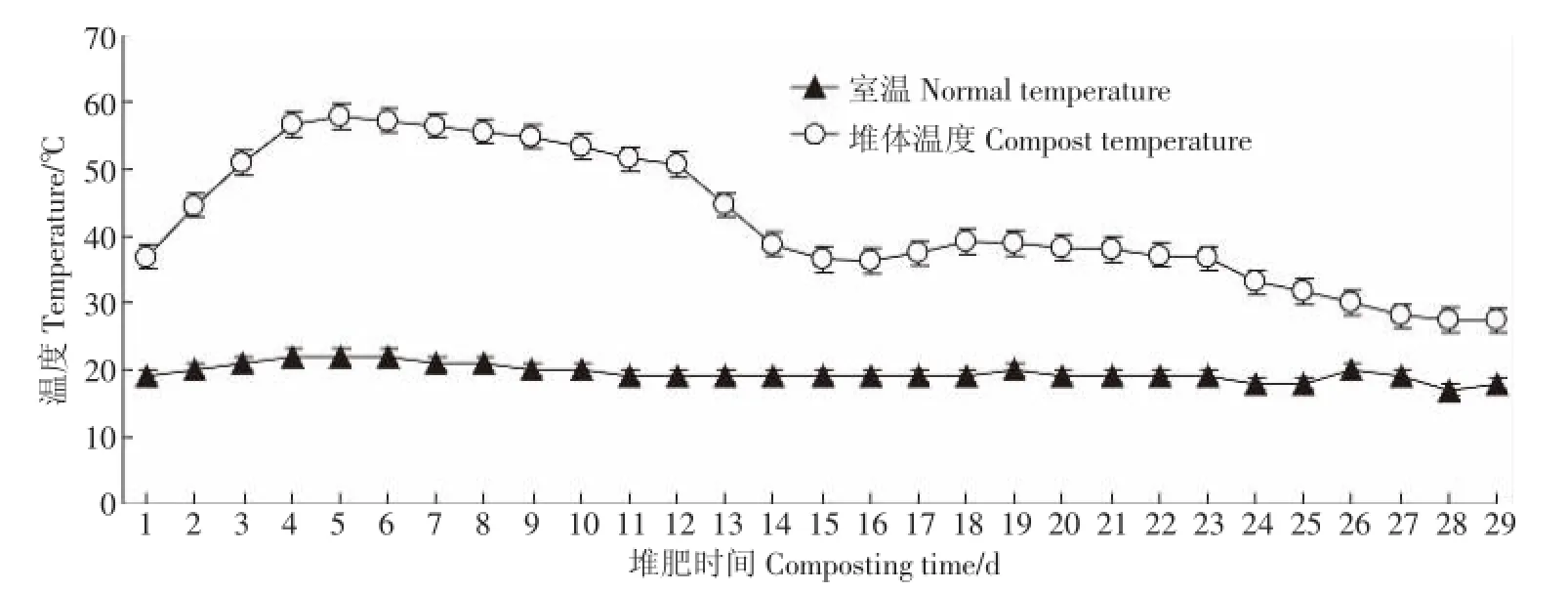
圖1 堆肥過程中溫度變化Figure 1 Changes of temperature during composting process
圖2為堆肥過程中含水率與pH的變化。堆肥過程中含水率的變化始終保持在40%~70%的范圍內,整體呈現下降的趨勢,在堆肥初期(0 d)堆肥含水率最大,達到65.7%,隨著堆體溫度升高,含水率逐漸變小,在堆肥結束時達到最低值42.1%。水分不僅直接影響堆體內微生物的代謝,而且會影響堆肥的腐熟速度[15]。堆肥過程中的pH變化在8.06~8.57之間,在堆肥第1 d降至最低值(8.06),第7 d時達到最大值(8.57),在堆肥后期(13~29 d)逐漸下降。因為在堆肥初期,微生物分解大量有機物的過程中產生有機酸,使得堆體pH值降低;后期有機酸的降解及NH+4-N的大量積累使得堆體pH逐漸升高[8]。
圖3為NH+4-N和-N含量變化。NH+4-N含量在堆肥前期(0~4 d)升高而在堆肥后期(7~29 d)呈迅速降低的變化趨勢,其含量從堆肥起始階段(0 d)的767 mg·kg-1迅速增加到第4 d時的最大值1159 mg· kg-1;隨著堆肥的進行,堆體溫度逐漸升高,大量NH3揮發[17],從而導致NH+4-N的含量迅速減少(4~29 d),堆肥結束時降至最小值453 mg·kg-1。而-N含量在整個堆肥過程中呈現上升趨勢,在堆肥初期-N含量僅為105 mg·kg-1,可能是由于在初期(0~3 d)某些具有硝化作用的微生物的活性受到了高溫以及NH+4-N大量積累的抑制,致使-N的生成速度減緩[18]。在堆肥4 d以后-N含量迅速增加,堆肥結束時達到425 mg·kg-1。
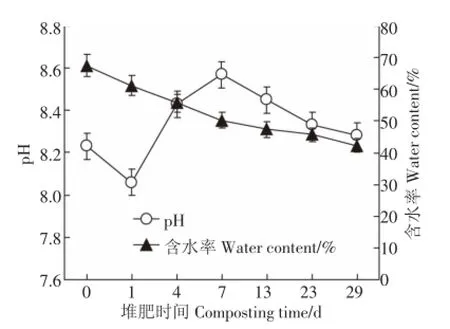
圖2 堆肥過程中pH和含水率變化Figure 2 Changes of pH and water content during composting process
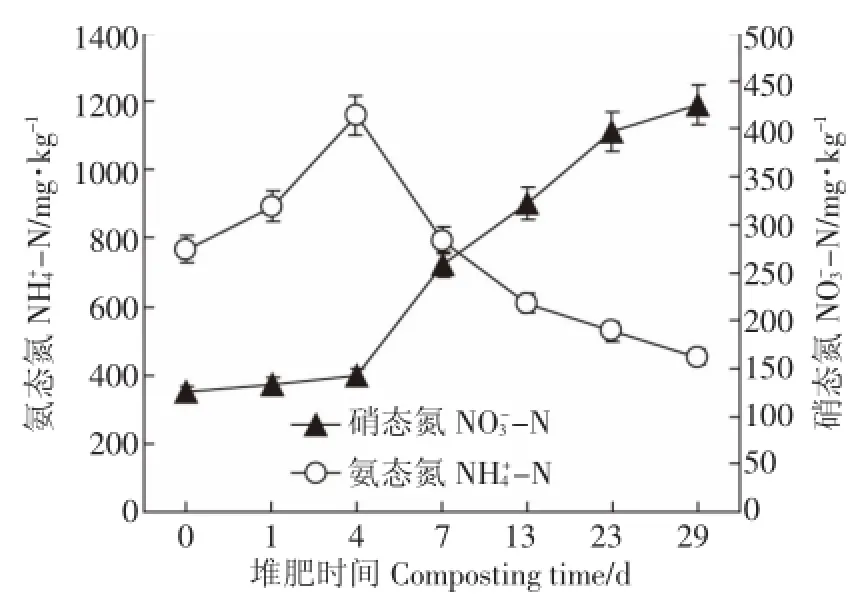
圖3 堆肥過程中NH+-N和NO--N變化43Figure 3 Changes of NH+-N and NO--N during43composting process
2.2 DGGE圖譜分析
不同時期的堆肥樣品PCR產物經DGGE分離,其結果如圖4所示。圖中字母代表條帶編號,在7個泳道上共識別出8條特異性的條帶。堆肥前期(0~7 d)樣品條帶的分布與后期(13~29 d)分布的差異性較大,說明隨著堆肥的進行,反硝化細菌的群落結構發生了明顯的變化。在堆肥過程中,每個時期均有條帶G出現,說明其具有較強的耐受性,為堆肥過程中的優勢種群;條帶B、C和F僅在堆肥高溫期出現,是高溫期的特異性條帶,具有較強的耐高溫性。
2.3多樣性分析
不同時期反硝化細菌群落的多樣性指數應用Shannon-Weaver公式進行計算[19],其結果如表2所示。反硝化細菌的多樣性總體呈現先增加后降低的趨勢。在堆肥開始時,反硝化細菌的多樣性指數呈整個堆肥過程中的最低值1.803,隨著堆肥的進行,反硝化細菌的多樣性逐漸升高,在高溫期(第4 d)達到最大值2.536,在堆肥后期多樣性指數有所下降。這是由于堆肥前期堆體內營養物質含量相對較為豐富,對反硝化細菌的生長較為有利;然而,不同菌屬的反硝化細菌對溫度等理化參數的敏感程度不盡相同,某些菌屬因適應不了高溫或因營養物質的減少而被淘汰,使得反硝化細菌的多樣性呈現高低不同的趨勢[20]。

圖4 堆肥樣品的DGGE圖譜Figure 4 DGGE profile of compost samples
2.4系統發育分析
要想在實際工作中實現對生產規模的提升,就應該進行必要的產業化生產,只有這樣才能在實際工作中實現對農產品成本的顯著控制與降低。而要想達成當前的工作目標,培育和發展新型經營主體是一項十分重要的環節。當前很多新型經營主體對于這一工作還缺乏必要的認識,導致出現了很多問題。
將8個成功測序的序列結果提交給Genbank并獲得登錄號:KU246118-KU246125。使用MEGA 4.0建立系統發育樹,其結果如圖5所示。所有的條帶測序結果與BLAST數據庫中比對的序列同源性菌高于55%。在堆肥過程中,條帶G測序結果與產堿菌屬(Alcaligenes)有較高的相似性,在堆肥各個時期始終出現,是堆肥過程中的優勢菌屬;條帶E和A分別在堆肥初期(1 d)和后期(13 d)被檢測到,經測序與副球菌屬(Paracoccus)和固氮螺菌屬(Azospirillum)有較高的相似性;條帶B、C和F僅在高溫期出現,其測序結果分別與無色桿菌屬(Achromobacter)、未培養細菌和假單胞菌屬(Pseudomonas)有較高的同源性。
2.5理化參數與反硝化細菌群落結構的相關性分析
堆肥過程中反硝化細菌群落結構與理化參數的冗余分析結果如圖6。前2個排序軸分別解釋了57.5%和21.1%的反硝化細菌種群變化,說明這些理化參數對堆肥過程中反硝化細菌群落結構的變化有顯著的影響。在RDA二維排序結果中,帶箭頭的矢量線表示不同的理化參數,圓圈代表在不同堆肥天數時取樣樣品。箭頭的矢量線長度可以表示不同理化參數與物種之間的相關性,其矢量線越長,相關性就越大,反之則越小[21]。從RDA排序圖中可以看出,含水率、硝態氮和氨態氮三個參數的連線相對較長,說明在堆肥過程中這些因素對反硝化細菌群落結構變化的影響較大。在RDA分析中,圓圈之間的距離表示菌落結構的差異性,圓圈間連線越短,差異性越小[13]。由此可知,反硝化細菌的群落結構在堆肥前期(0~4 d)變化較小。在降溫腐熟期(23~29 d)變化則較為劇烈。
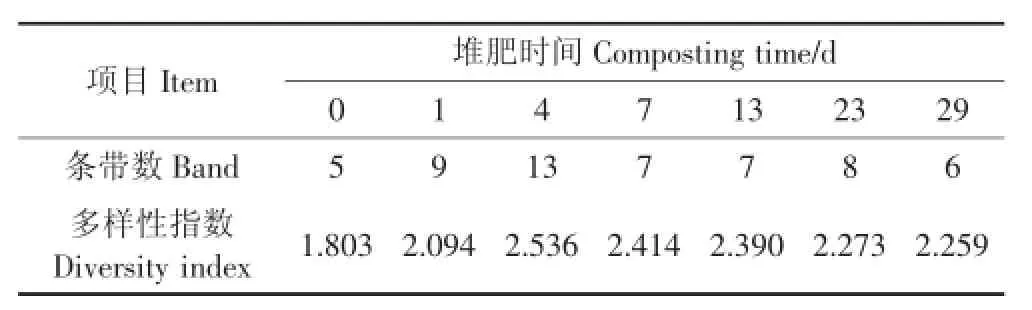
表2 反硝化細菌的多樣性指數Table 2 Diversity index of denitrifying bacteria

圖5 反硝化細菌的系統進化樹Figure 5 Phylogenetic tree of denitrifying bacteria
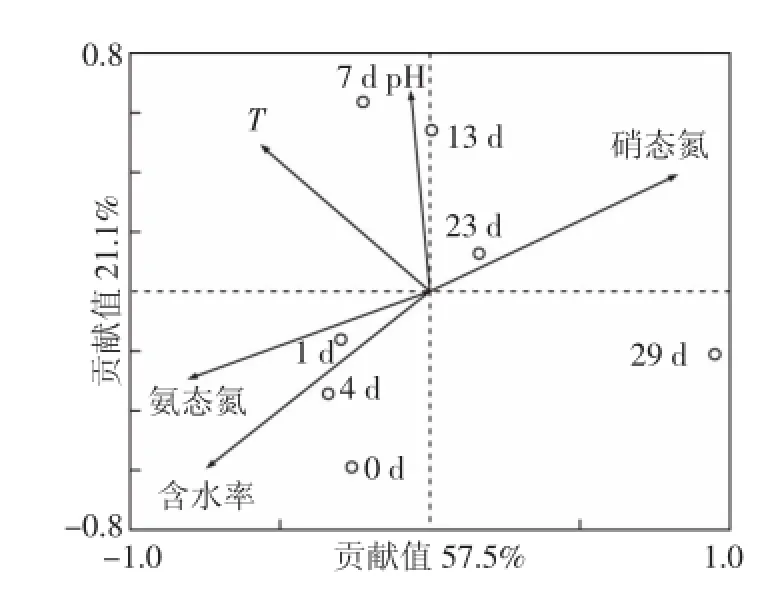
圖6 反硝化細菌種群變化與理化參數的RDA圖Figure 6 RDA of denitrifying bacteria community and physic-chemical parameters
堆肥過程中理化參數(含水率、硝態氮和氨態氮)對反硝化細菌群落結構變化有不同程度的影響。含水率與堆肥中溶解氧的濃度密切相關,當含水率較大時,堆體中溶解氧濃度較低[22],利于反硝化菌的生長,因為多數反硝化菌屬厭氧微生物。在堆肥初始階段(0~3 d),堆體含水率較大,在一定程度上使得堆體內成厭氧或缺氧的狀態,因而反硝化菌的生長較為穩定,其群落結構變化較小;在堆肥后期(4~29 d),含水率降低使得溶氧量相對較高,使得堆體內的反硝化細菌群落結構發生了較大的變化。
過去的研究中,有學者發現,硝態氮含量可以影響反硝化速率與菌群的群落結構變化[23]。在本文中,硝態氮與反硝化細菌群落結構有一定的相關性,也有學者研究發現,反硝化細菌的變化與硝態氮濃度并無顯著的相關性[24]。這些結果表明,在不同的環境條件下,硝態氮濃度對反硝化細菌群落變化的影響存在顯著性的差異。堆肥的前期(0~4 d),堆肥中反硝化細菌的群落結構變化較小,在此階段中,供反硝化細菌作用的營養物質相對較多,反硝化細菌作用時相對穩定,所以,群落結構變化不大,其硝態氮含量逐漸積累增加;但在堆肥后期(23~29 d),反硝化細菌群落結構變化卻很大,其原因可能是因為堆肥后期營養物質的減少以及大量硝態氮的積累,對反硝化細菌的作用產生了一定程度的影響。所以,反硝化細菌群落結構變化與理化因子之間的關系還需更多的研究。氨態氮表面上雖然與反硝化細菌的作用并無直接聯系,但在氨態氮氧化生成硝態氮的過程中,會產生大量N2O氣體,此氣體作為反硝化功能基因(nosZ)的底物,其含量多少會對反硝化細菌的作用產生影響。胡春曉等[25]也在對農業廢棄物好氧堆肥過程的研究中發現,氨態氮濃度與nosZ基因呈顯著正相關關系。
反硝化細菌的變化,不僅體現在其群落結構與理化參數的相關性,還有反硝化細菌群落多樣性與理化參數之間的相關性。理化參數與反硝化細菌群落多樣性的皮爾遜相關性分析結果如表3所示。堆肥過程中反硝化細菌群落多樣性與含水率呈極顯著正相關關系(r=0.960,P<0.01),說明隨著含水率的變大,反硝化細菌的群落多樣性呈現顯著性的增加,反之則減少;溫度與反硝化細菌群落多樣性呈顯著正相關關系(r= 0.766,P<0.05),說明隨著溫度的增加,反硝化細菌多樣性也增加,反之則呈下降的趨勢。但溫度卻不是影響反硝化細菌多樣性的最主要因素,溫度的變化通過對溶氧量及含水率等因素的影響,從而對反硝化細菌的多樣性有一定程度的影響[26-28]。含水率對反硝化細菌群落多樣性的影響呈現了極顯著的相關性,說明反硝化細菌群落多樣性對堆體內含水率的大小敏感程度很大,其細微的變化就能引起多樣性的改變。

表3 理化參數與反硝化細菌群落多樣性的皮爾遜相關分析Table 3 Pearson correlations between physic-chemical parameters and diversity index of denitrifying bacteria
溫度也可以對其他理化參數產生影響,從而間接地影響微生物的代謝活動。在堆肥過程中,溫度的變化使得堆體內含水率有所不同,其含水率的大小與溶氧濃度存在著一定的相關性。堆體內含水率較大時,溶氧濃度相對較低,反之溶氧量含量較高[29]。有大量的研究表明,當溶解氧濃度在一定的范圍內時,反硝化細菌的活性不會受到溶解氧濃度的影響[30];當溶解氧的濃度低于某個值的時候,反硝化細菌的活性隨著溶解氧濃度的降低而升高。這就說明影響反硝化細菌活性的溶解氧濃度存在著一個極限值,也稱為閾值[31]。堆肥過程溶氧的變化可能受堆肥水分的影響,因此堆肥過程中反硝化細菌群落多樣性隨含水率的變大而升高。
3 結論
(1)在牛糞堆肥過程中,反硝化細菌群落結構在堆肥前期(0~4 d)變化較小,在堆肥后期(23~29 d)變化較為劇烈;反硝化細菌群落多樣性總體呈現先升高(0~4 d)后降低的趨勢(23~29 d);
(2)Alcaligenes(條帶G)存在于整個堆肥過程中,有較強的耐受性,是堆肥過程中反硝化細菌的優勢種屬;Achromobacter(條帶B)、未培養細菌(條帶C)和Pseudomonas(條帶F)僅出現在堆肥高溫期,具有較強的耐高溫性,是高溫階段的優勢性菌屬。
(3)堆體含水率、硝態氮和氨態氮對群落結構變化有著不同程度的影響。含水率與反硝化細菌群落多樣性呈極顯著正相關關系,溫度與反硝化細菌群落多樣性呈顯著正相關關系。
[1]Jusoh M L C,Manaf L A,Latiff P A.Composting of rice straw with effective microorganisms(EM)and its influence on compost quality[J].I-ranian Journal of Environmental Health Sciences and Enineering,2013,10(1):1-9.
[2]張玉芹,劉開啟,王革,等.反硝化細菌的篩選及培養條件的研究[J].農業環境科學學報,2005,24(1):18-25.
ZHANG Yu-qin,LIU Kai-qi,WANG Ge,et al.Screening of denitrifying bacteria and determination of the culture conditions[J].Journal of Agro-Environment Science,2005,24(1):18-25.
[3]Jiang T,Schucharde F,Li G X,et al.Effect of C/N ratio,aeration rate and moisture content on ammonia and greenhouse gas emission during the composting[J].Journal of Environmental Science,2011,23(10):1754-1760.
[4]王海濤,鄭天凌,楊小茹.土壤反硝化的分子生態學研究進展及其影響因素[J].農業環境科學學報,2013,32(10):1915-1924.
WANG Hai-tao,ZHENG Tian-ling,YANG Xiao-ru.Molecular ecology research for soil denitrification and research status for its influencing factors[J].Journal of Agro-Environment Science,2013,32(10):1915-1924.
[5]Wang Q L,Hu N,Ding D X,et al.Removal of-N from polluted groundwater in decommissioned mining area in an in-situ leach uranium mine by denitrifying bacteria bioreactor[J].Atomic Energy Science and Technology,2013(8):7-13.
[6]Barrett M,Jahanigir M M,Lee C,et al.Abundance of denitrification genes under different peizometer depths in four Irish agricultural groundwater sites[J].Environmental Science and Pollution Research,2013,20(9):6646-6657.
[7]Wei L,Ma F,Su J F,et al.Rapid quantitative detection of denitrifying bacteria in terms of nirS gene[J].Journal of Harbin Institute of Technology,2008,40(10):12-18.
[8]鮑士旦.土壤農化分析[M].北京:中國農業出版社,2000.
BAO Shi-dan.Soil agricultural chemistry analysis[M].Beijing:China Agriculture Press,2000.
[9]Tago K,Okubo T,Shimomura Y,et al.Environmental factors shaping the community structure of ammonia-oxidizing bacteria and archaea in sugarcane field soil[J].Microbes Environmental,2015,30(1):21-28.
[10]Su J F,Ma F,Gao S S,et al.Aerobic denitrifiers dispose the wastewater of NO3-N and the analysis of the diversity of microbial species[J].Journal of Basic Science and Engineering,2008,16(1):45-50.
[11]王曉君,陳少華,張兆基,等.利用氧化亞氮還原酶基因(nosZ)評價人工濕地系統中的反硝化菌[J].環境科學,2012,33(4):1306-1311.
WANG Xiao-jun,CHEN Shao-hua,ZHANG Zhao-ji,et al.Denitrifying bacteria of constructed wetland system based on the nitrous oxide reductase gene(nosZ)[J].Environmental Science,2012,33(4):1306-1311.
[12]Angnes G,Nicoloso R S,Sliva M L B,et al.Correlation denitrifying catabolic genes with N2O and N2emissions from swine slurry composting[J].Bioresource Technology,2013,6:368-375.
[13]Valentín-Vargas A,Root R A,Neilson J W,et al.Environmental factors influencing the structural dynamics of soil microbial communities during assisted phytostabilization of acid-generating mine tailings:A mesocosm experiment[J].Science of the Total Environment,2014,500:314-324.
[14]龍健,廖洪凱,李娟,等.基于冗余分析的典型喀斯特山區土壤-石漠化關系研究[J].環境科學,2012,33(6):2132-2137.
LONGJian,LIAOHong-kai,LIJuan,etal.Relationshipsbetweensoiland rockydesertificationintypicalkarstmountainareabased on redundancy analysis[J].Environmental Science,2012,33(6):2132-2137.
[15]唐珠,沈其榮,余光輝.豬糞堆肥的理化特征及腐熟度評價研究[D].南京:南京農業大學,2011:85.
TANG Zhu,SHEN Qi-rong,YU Guang-hui.Characterization and indentification of maturity of pig manure composting[D].Nanjing:Nanjing Agricultural University,2011:85.
[16]柴曉利,張華,趙由才.固體廢物堆肥原理與技術[M].北京:化學工業出版社,2005:83.
CHAI Xiao-li,ZHANG Hua,ZHAO You-cai.The principle and tech-nology of soild waste composting[M].Beijing:Chemical Industry Press,2005:83.
[17]Liu S T,Wang G F,Xie X F,et al.Effect of matrix on denitrification efficiency and distribution of nitrifying and denitrifying bacteria in constructed wetlands[J].Journal of Southeast University(Natural Science Edition),2011,41(2):15-18.
[18]Li H B,Lian J,Li M,et al.Isolation and characteristics of denitrifying bacterium strain GW1[J].Journal of Hebei University of Science and Technology,2012,33(2):20-24.
[19]董西洋.反硝化細菌對絡合脫硝液中Fe(Ⅲ)EDTA和Fe(Ⅱ)EDTA-NO的還原[D].大連:大連理工大學,2013:19-23.
DONG Xi-yang.Reduction of Fe(Ⅲ)EDTA and Fe(Ⅱ)EDTA-NO by dinitrifying bacteria in a NOx scrubber liquou[D].Dalian:Dalian University of Technology,2013:19-23.
[20]Yi N,Di P P,Wang Y,et al.Diel vertical variability analysis of denitrifying bacteria abundance in oxygen-enriched irrigation pond[J].Transactions of the Chinese Society of Agricultural Engineering,2015,30 (15):22-26.
[21]Zhu L,Yin L H,Pu Y P,et al.Optimization and application of fluorescence in situ hybridization assay for detecting nitrifying bacteria in environmental samples[J].Journal of Southeast University(Natural Science Edition),2005,35(2):21-23.
[22]Su J F,Ma F,Wang J H,et al.Nitrification and denitrification characteristics of new heterotrophic nitrifying bacteria[J].Journal of Tianjin University,2007,40(10):7-14.
[23]Miao Y,Liao R H,Zhang X X,et al.Metagenomic insights into salinity effect on diversity and abundance of denitrifying bacteria and genes in an expanded granular sludge bed reactor treating high-nitratewastewater[J].ChemicalEngineeringJournal,2015,10:116-123.
[24]Ligi T,Truu M,Turr J,et al.Effects of soil chemical characteristics and water regime on denitrifiction genes(nirS,nirK,and nosZ)abundances in a created riverine wetland complex[J].Ecological Engineering,2014, 11:47-55.
[25]胡春曉,陳耀寧,張嘉超,等.農業廢物好氧堆肥中環境因子對nirK、nirK和nosZ數量的影響[J].環境科學,2013,34(3):1196-1302.
HU Chun-xiao,CHEN Yao-ning,ZHANG Jia-chao,et al.Effects of physico-chemical parameters on the abundance of the denitrification associated genes nirK,nirS and nosZ during agricultural waste composting[J].Environmental Science,2013,34(3):1196-1302.
[26]Zhang L H,Dou C L,Shi H F,et al.Experimental study on the nitrification process in the bio-cathode of microbial fuel cells[J].Environmental Pollution and Control,2012,34(9):26-30.
[27]Chen Q Y,Jiang L F,Chen J Q,et al.Isolation and NH+4-N removal characteristics of a high-efficient heterotrophic nitrifying bacterium alcaligenesfaecalisNi3-1[J].EnvironmentalEngineering,2015,33(5):3-7.
[28]李衛芬,鄭佳佳,張小平,等.反硝化酶及其環境影響因子的研究進展[J].水生生物學報,2014(1):166-170.
LI Wei-fen,ZHENG Jia-jia,ZHANG Xiao-ping,et al.Progress in studies on denitrification enzymes and environmental impact factors[J]. Acta Hydrobiologica Sinica,2014(1):166-170.
[29]Michael-Kordatou I,Michael C,Duan X D,et al.Dissolved effluent organic matter:Characteristics and potential implications in wastewater treatment and reuse applications[J].Water Research,2015,6:213-248.
[30]張雪辰.禽畜糞便堆肥化過程碳氮轉化規律及其受調控措施的影響[D].楊凌:西北農林科技大學,2014.
ZHANG Xue-chen.The transformation of organic carbon and nitrogen with different regulation technology during the livestock manure composting process[D].Yangling:Northwest A&F University,2014.
[31]Niu T H,Zhou Z,Shen X L,et al.Effects of dissolved oxygen on performance and microbial community structure in a micro-aerobic hydrolysis sludge in situ reduction process[J].Water Research,2016,5:369-377.
Correlationship between denitrifying bacteria and physicochemical factors in cow manure composting
ZHU Li-ping,XU Xiu-hong*,ZHANG Wen-hao,CHENG Li-jun,LI Hong-tao,WANG Zi-qi,HE Ning
(School of Resources and Environmental Sciences,Northeast Agricultural University,Harbin 150030,China)
In the present paper,the community structure of denitrifying bacteria during cow manure composting process was analyzed by polymerase chain reaction-denaturing gradient gel electrophoresis(PCR-DGGE)based on the nosZ gene.The relationship between the changesofdenitrifyingbacteria community structure and the physic-chemical parameters(temperature,pH,water content was evaluated by redundancy analysis(RDA).Results showed that the Shannon-Weaver index of denitrifying bacteria increased during 0~4 d but decreased during 7~29 d of the composting.Based on the sequencing of dominant bands,Alcaligenes were the dominant species during composting.Redundancy analysis(RDA)showed that the community structure of dinitrifying bacteria changed little at the early stage of composting(0~4 d),while it changed dramatically in the maturity stage of composting(23~29 d).Pearson correlation analysis showed that denitrifying bacteria community diversity was extremely significantly positively correlated with water content(r=0.960,P≤0.01),and significantly positively correlated with temperature(r=0.766,P≤0.05).
compost;denitrifier;physico-chemical parameter;bacterial community structure;redundancy analysis
X713
A
1672-2043(2016)09-1781-07doi:10.11654/jaes.2016-0250
2016-03-01
國家自然科學基金項目(31272484,31372351)
朱麗平(1990—),女,碩士,研究方向為微生物。E-mail:502348499@qq.com
許修宏E-mail:xuxiuhong@neau.edu.cn
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
