封育年限對高寒草甸群落組分和物種多樣性的影響
2016-10-25 08:02:52劉曉琴張立鋒李英年徐世曉李紅琴馬榮榮高玉葆
生態學報 2016年16期
劉曉琴,張 翔,張立鋒,李英年,趙 亮,徐世曉,李紅琴,馬榮榮,牛 犇,高玉葆,古 松,*
1 南開大學生命科學學院, 天津 300071 2 中國科學院西北高原生物研究所, 西寧 810008
?
封育年限對高寒草甸群落組分和物種多樣性的影響
劉曉琴1,張翔1,張立鋒1,李英年2,趙亮2,徐世曉2,李紅琴2,馬榮榮1,牛犇1,高玉葆1,古松1,*
1 南開大學生命科學學院, 天津300071 2 中國科學院西北高原生物研究所, 西寧810008
高寒草甸在青藏高原畜牧業發展中占有重要地位,然而氣候和人為活動的干擾對原本脆弱的生態系統產生了深刻影響。雖然封育是被廣泛采用的草地恢復有效措施之一,但封育年限對群落組分和多樣性的影響一直是受關注的核心問題,探明這一問題對高寒草甸的可持續發展具有重要意義。研究了封育2、7、17a的高寒矮嵩草(Kobresiahumilis)草甸的群落組分和物種多樣性變化特征,并探討了不同封育年限影響物種多樣性的可能原因。結果表明:1)隨封育年限延長,群落上層禾本科植物的總體重要值明顯增加,其中異針茅(Stipaaliena)最為顯著,其重要值封育2a(13.87%)< 封育7a(21.76%)< 封育17a(23.95%),而豆科植物總體重要值顯著降低,莎草科和雜類草植物變化不明顯;2)α多樣性:隨封育年限延長,高寒草甸群落物種豐富度指數(Patrick和Margalef指數)沒有發生顯著變化,然而物種多樣性指數(Simpson、Shannon-Wiener和種間相遇機率指數)及均勻度指數(Alatalo和Pielou指數)均表現為封育2a > 封育7a > 封育17a,且封育2a與封育7、17a呈顯著性差異(P<0.05);3)β多樣性:Cody指數和Morisita-Horn相似指數的變化表明,封育2a與封育7、17a之間的物種差異較大,而封育7a和17a之間的物種差異較小;4)封育后凋落物的覆蓋時間及其分解引起的土壤養分變化是導致物種多樣性降低的重要原因之一,土壤碳氮比(C/N)與物種均勻度指數(Alatalo指數)和多樣性指數(Simpson指數和種間相遇機率指數)均呈顯著性負相關關系(P<0.05),說明土壤養分元素之間的耦合效應對物種多樣性變化具有重要影響。本研究說明,雖然物種豐富度指數隨封育年限的延長沒有變化,但物種均勻度和多樣性指數均降低,且封育前期對物種多樣性的影響大于后期,就物種多樣性而言,該高寒草甸不宜進行長期封育。該研究以期為高寒草甸生物多樣性的保護和可持續利用及管理提供科學依據。
青藏高原;豐富度指數;均勻度指數;多樣性指數;β多樣性
青藏高原的植被類型多樣、植物種類豐富,是我國生物多樣性最為豐富的地區之一,也是我國生物多樣性的重要保護基地。其中高寒草甸是最主要的植被類型,其生態系統對氣候變化的響應異常敏感[1]。高寒草甸不僅在畜牧業可持續發展中占有十分重要的地位,而且在區域氣候調節、水源涵養、生物多樣性保護和碳源/匯等方面均發揮著不可替代的作用[2]。然而,由于高原生態環境的脆弱性,在氣候變化、超載放牧和生物資源的不合理開發利用等自然和人為因素影響下,導致高寒草甸的退化,并引起群落組分和物種多樣性的改變以及生產力下降等一系列生態問題[3]。據統計,高原上約有0.45×108hm2草地出現了不同程度的退化,占青藏高原草地總面積的1/3,對高寒草甸生態系統的可持續發展造成了嚴重的威脅[4]。因此,保護高寒草甸的生物多樣性、生態系統的穩定性及其服務功能已引起科學界和政府的高度重視。
圍欄封育作為恢復草地生態系統最主要的措施之一而被廣泛采用[5],封育不僅可以改變草地群落的覆蓋度、生物量等數量特征,同時也改變了其土壤理化性質和外界環境特征[6]。物種多樣性是評價草場封育效果的重要指標,通常封育可以避免外界干擾,使退化草場植物在一定時間內得以繁衍,使物種多樣性增加,植物群落逐漸恢復[7]。但是,封育在草地恢復過程中也有不利的一面,因為封育本身也是一種人為干擾,消除了大型食草動物對草地的采食和踐踏,切斷了封育草地與外界生態系統能量與物質交換的通道[8]。因此,探明封育對草地生態系統群落結構和物種多樣性的影響已成為近年的研究熱點,這對于準確評估封育年限效果與草地的可持續利用具有重要意義。
封育年限是影響草地群落組分和物種多樣性變化的重要因子,國內外學者對此做了大量的研究和報道[9- 13],但由于草地類型、氣候、土壤等差異,其結果也不盡相同。對我國溫性草原的研究表明,隨圍封年限的延長物種多樣性呈逐漸增加趨勢[9];但也有研究指出,物種多樣性隨封育年限的延長呈“增加→降低→增加”或“降低→增加→降低”的變化[10, 11]。對溫帶沙化草地的研究結果表明,隨著圍封年限的增加,呼倫貝爾沙化草地的群落結構逐漸趨于合理,物種豐富度、群落均勻性和多樣性均呈增加趨勢[12];而科爾沁沙化草地的物種多樣性指數則隨封育時間延長呈現下降趨勢,即封育不利于物種多樣性的維持[13]。因此,封育年限對草地生態系統群落結構和物種多樣性的影響非常復雜,不僅表現在不同類型草地之間的差異,即使對同一類型草地也有不同的結果。
在高原極端環境影響下形成的高寒草甸生態系統極其脆弱,對氣候變化和人為干擾非常敏感[1]。然而,就封育年限對高寒草甸群落組分和物種多樣性影響的研究相對匱乏,雖然近年有一些關于圍封對高寒草地群落影響的報道[14- 17],但其中大多以短期封育為主。青藏高原幅員遼闊,草地和氣候類型迥異,結果也不盡相同,更缺少長期封育對高寒草甸生物多樣性影響的研究。本研究在位于青藏高原東北隅的中國科學院高寒草甸生態系統定位站,對封育2、7、17a的高寒草甸群落結構、物種多樣性、凋落物和土壤養分等進行了調查與測定,其主要目的是:1)揭示不同封育年限下的高寒草甸群落組分及物種多樣性狀況;2)探討封育年限對高寒草甸物種多樣性的影響及其主要原因,以期為保護高寒草甸在圍欄封育過程中的生物多樣性以及畜牧業的可持續發展提供科學依據。
1 材料與方法
1.1研究區概況
實驗地位于中國科學院海北高寒草甸生態系統定位站(簡稱海北站),地理位置為37°29′—37°45′N,101°12′—101°23′E,海拔3200—3600 m,地處青藏高原東北隅祁連山北支冷龍嶺東段南麓坡地的大通河河谷。該區位于亞洲大陸腹地,屬明顯的高原大陸性氣候,東南季風及西南季風微弱。受高海拔條件影響,該區輻射強,氣溫低,無明顯四季之分,僅有冷暖季之別,干濕季分明。年平均氣溫 -1.7 ℃,降水量約560 mm,其中降水量的80%以上集中在植物生長季的5—9月,年均實際日照時數為2467.7 h,可照日照時數為4434.7 h[18]。
實驗區地勢開闊平坦,植被類型為高寒矮嵩草草甸,物種分布均勻,在垂直結構上一般分上下兩層[19],上層包括禾草類和較高類雜草,優勢種有異針茅(Stipaaliena)、山地早熟禾(Poaorinosa)、羊茅(Festucaovina)等;下層包括莎草類和較低類雜草,優勢種為矮嵩草(Kobresiahumilis)、麻花艽(Gentianastraminea)等,土壤為草氈寒凍雛形土(Mat-Cryic Cambisols)。
1.2研究方法
在海北站高寒矮嵩草草甸選取3塊試驗樣地進行封育,至2013年底,封育年限分別為2、7、17a,面積分別為650、6000 m2和900 m2,樣地之間相隔約300 m。于2013年8月底,在不同封育年限樣地的觀測區內隨機選取5個50 cm × 50 cm的樣方,將樣方分為100個5 cm × 5 cm小方格,采用針刺法測量樣方內各植物種的高度和蓋度,再將樣方內植物分種齊地面剪掉,并收集各樣方內凋落物,分別放入紙袋,帶回實驗室烘干稱重,統計各物種的生物量。然后清除樣方內表層土,使用直徑1.5 cm土鉆分別對不同封育年限樣地0—10 cm土壤進行取樣(每個樣地5個重復),放入自封袋保存,帶回實驗室風干,分別采用重鉻酸鉀容量法、半微量凱氏定氮法和硫酸-高氯酸消煮法測定土壤有機碳(SOC)、全氮(STN)和全磷(STP)含量。
1.3計算方法
本文計算物種重要值、α和β多樣性,其中α多樣性包括豐富度指數、多樣性指數和均勻度指數,β多樣性包括Whittaker 指數、Cody指數和Morisita-Horn相似指數,具體計算公式如下:
(1) 物種重要值[20]
IV=(RB+RC+RH)×100/3
式中,RB為相對地上生物量,即單種植物地上生物量占所有物種地上生物量和的比例;RC為相對蓋度,即單種植物蓋度占所有物種蓋度和的比例;RH為相對高度,即單種植物自然高度占所有物種自然高度和的比例。
(2) α多樣性指數[21-22]
1) 豐富度指數
Patrick 指數(R)
R = S′
Margalef 指數(Ma)
Ma =(S′-1)/ln(B)
2) 多樣性指數
Simpson 指數(D)
Shannon-Wiener 指數(H′)
種間相遇機率指數(PIE)
3) 均勻度指數
Pielou 指數(Jsw)
Jsw= H′/ln(S′)
Alatalo 指數(Ea)
式中,S′為樣地樣方的平均物種數;B為群落的總生物量;pi為第i種物種的相對重要值;Ni為物種i的重要值;N為物種i所在樣方的各個種的重要值之和。
(3)β多樣性指數[23]
1)Whittaker指數(βws)
βws= S/ma - 1
2)Cody指數(βc)
βc= [ g(H) + I(H)]/2
3) 經Wolda改進的Morisita-Horn指數(CMH)
式中,S為各封育樣地出現的物種總數,ma為各樣方的平均物種數;g(H)是沿生境梯度H而增加的物種數,I(H) 是沿生境梯度H而失去的物種數;pia和pib為a和b樣地中第i種植物的相對重要值,Sa和Sb為a和b樣地出現的物種總數。
1.4數據分析
采用SPSS17.0統計軟件對數據進行統計分析。在P= 0.05顯著水平上,One-way ANOVA和LSD分析方法進行差異性檢驗,Pearson法進行相關性檢驗。
2 結果與分析
2.1不同封育年限的群落組分變化
不同封育年限高寒草甸的植物群落結構和組成發生了明顯變化(表1和圖1)。封育2a樣地共有44種植物,分屬于39個屬、17個科;封育7a樣地有44種植物,分屬于39個屬、15個科;封育17a樣地則41種植物,分屬于35個屬、15個科。從功能群組成來看,在封育2、7、17a樣地的群落上層,禾本科植物占絕對優勢,總體重要值分別為35.02%、34.23%和40.99%(圖1)。封育2a與7a之間的群落物種數和禾本科植物的重要值均未發生明顯變化;但與封育2a和7a相比,封育17a群落物種數降低了6.81%,禾本科植物重要值分別增加了17.05%和19.75%。值得注意的是,在封育2a的樣地中,異針茅(Stipaaliena)與山地早熟禾(Poaorinosa)的重要值分別為13.87% 和10.53%,而在封育7a和17a樣地中,異針茅的重要值分別增加到21.76%(P<0.05)和23.95%(P<0.05),山地早熟禾則分別降低到3.92%(P<0.05)和5.70%(P<0.05),即隨封育年限延長,群落上層逐漸由共優勢種的異針茅和山地早熟禾變為單優勢種的異針茅。

表1 不同封育年限高寒草甸群落物種組成和重要值
表中“-”表示該種植物不存在

圖1 不同封育年限高寒草甸群落功能群 Fig.1 Community functional group for different exclosure duration (year) in alpine meadow圖中字母不同表示差異顯著(P<0.05)
莎草科植物的總體重要值隨封育年限延長而降低,封育2、7、17a樣地分別為10.32%、10.11%和8.36%(圖1),沒有顯著變化。下層優勢種矮嵩草(Kobresiahumilis)的重要值封育2、7、17a分別是6.71%、6.43%和7.06%,未發生顯著變化,即不受封育年限的影響。
豆科植物的總體重要值在封育2、7、17a樣地分別是5.77%、4.82%和3.23%,封育7a和17a分別比封育2a樣地降低了16.46%和44.02%(P<0.05)(圖1)。
雜類草植物的總體重要值封育2、7、17a分別是47.96%、51.49%和49.53%(圖1),沒有發生顯著變化。但其中的幾種植物重要值卻發生了較大變化。例如,鵝絨委陵菜(Potentillaanserina)的重要值為封育2a(5.23%)> 封育17a(2.10%)> 封育7a(0.51%);摩苓草(Morinachinensis)重要值為封育17a(6.87%)> 封育7a(4.03%)> 封育2a(2.76%);黃帚橐吾(Ligulariavirgaurea)在封育2a樣地中的重要值為2.71%,但沒有出現在封育7a和17a樣地中;箭葉橐吾(Ligulariasagitta)的重要值為封育2a(3.40%)> 封育7a(2.68%)> 封育17a(0.21%);西伯利亞蓼(Polygonumsibiricum)的重要值在封育2a和17a樣地中分別為1.87%和1.13%,而沒有出現在封育7a樣地;美麗風毛菊(Saussureapulchra)重要值為封育7a(5.22%)> 封育17a(2.07%)> 封育2a(0.60%);線葉龍膽(Gentianafarreri)的重要值是封育17a(1.94%)> 封育7a(0.62%)> 封育2a(0.01%)。該結果表明,雖然雜類草植物總體重要值隨封育年限延長未出現顯著性變化,但封育對其群落內部物種的組成及重要值產生了較大影響。

圖2 不同封育年限高寒草甸群落物種重要值排序 Fig.2 Ranking of species important values for different exclosure duration (year) in alpine meadow
由物種重要值的大小排序可以發現(圖2),不同封育年限的物種重要值變化大致從第10個物種開始趨于重合,而前9個物種占整體重要值的比例較大,封育2、7、17a樣地分別為58.02%、62.15%和65.92%。此外,不同封育年限之間的變化趨勢有所不同:封育2a樣地降低速率最為緩慢,且趨勢線最平滑;封育7a樣地降低速率加快,但波動很小;而封育17a樣地不僅降低速率加快且趨勢線波動最大,其主要原因是隨封育年限增加異針茅重要值迅速增加,而山地早熟禾則呈相反變化。
隨封育年限的延長,凋落物量呈下降趨勢,但差異不顯著(圖3);群落上層重要值最大的異針茅相對高度變化由封育2a的6.11%增加到封育7a和17a的8.13%和7.46%(P<0.05)(7a與17a之間不顯著)(圖3)。
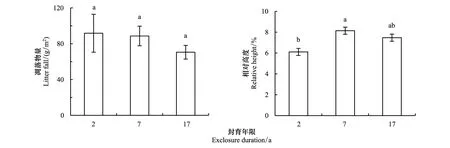
圖3 不同封育年限高寒草甸凋落物量和異針茅相對高度Fig.3 Litter fall and relative height of Stipa aliena for exclosure duration (year)圖中字母不同表示差異顯著(P<0.05)
2.2不同封育年限的物種多樣性2.2.1不同封育年限的α多樣性
在不同封育年限樣地中,表示物種豐富度的Patrick指數(R)和Margalef指數(Ma)(圖4)均未發生顯著性變化,3個封育年限樣地的R和Ma分別在34和7.7左右,說明封育年限對物種豐富度并未造成明顯影響。然而,表示物種均勻度的Pielou指數(Jsw)和Alatalo 指數(Ea)(圖4)均為封育2a > 封育7a > 封育17a。其中,封育2a與封育17a樣地之間的Jsw差異顯著(P<0.05),而封育7a與另外兩樣地的差異不顯著;Ea則隨封育年限延長呈顯著性降低(P<0.05)。

圖4 不同封育年限高寒草甸群落α多樣性Fig.4 α diversity of alpine meadow for different exclosure duration (year)圖中小寫字母不同表示差異顯著(P<0.05)
表示物種多樣性的Simpson指數(D)、Shannon-Wiener 指數(H′)(圖4)和種間相遇機率指數(PIE)(圖4)均為封育2a> 封育7a> 封育17a,而封育2a樣地顯著高于封育7a和17a樣地(P<0.05),但封育7a和17a樣地之間的差異不顯著。
2.2.2不同封育年限的β多樣性
不同封育年限的Morisita-Horn(CMH)指數和Cody(βc)指數表明(表2),封育2a和7a樣地的CMH值居中(0.85),βc值最大(5.0),封育7a較封育2a增加和減少的物種數均為5;封育2a和17a的CMH值最小(0.83),βc值居中(4.5),封育17a較封育2a增加和減少的物種數分別為3和6;封育7和17aCMH值最大(0.94),βc值最小(3.5),封育17a較封育7a增加和減少的物種數分別為2和5,說明封育2a與7、17a樣地之間的物種差異較大,而封育7a和17a樣地之間的物種差異相對較小。
封育2、7、17a樣地的Whittaker(βws)指數分別為0.26、0.33和0.21(圖5),說明封育2a樣地群落內物種更替程度居中,封育7a樣地物種更替程度最大,樣方間種類組成相似性最小;封育17a則物種更替程度最小,樣方間種類組成相似性最大。β多樣性指數綜合結果說明,物種多樣性對封育年限的響應程度在封育前期(封育2和7a之間)大于封育后期(封育7和17a之間),這與α多樣性的結果相一致。

表2 不同封育年限之間高寒草甸Cody指數(βc)和Morisita-Horn(CMH)相似指數

圖5 不同封育年限高寒草甸Whittaker指數(βws) Fig.5 Whittaker index for different exclosure duration in alpine meadow
2.2.3不同封育年限的物種多樣性相關性
豐富度指數(Ma)與Whittaker指數(βws)呈顯著正相關關系(P<0.05)(圖6),多樣性指數(D、H′和PIE)隨均勻度指數(Jsw和Ea)的增加呈顯著性上升趨勢(P<0.05)(圖6)。
2.3不同封育年限的土壤養分與物種多樣性相關性
封育2、7、17a的SOC、STN、STP含量以及土壤氮磷比(N/P)均表現為封育17a > 封育2a > 封育7a,土壤碳氮比(C/N)沒有發生顯著變化(表3)。SOC、STN、STP含量與α和β多樣性指數的相關性均不顯著,土壤C/N與多樣性指數和均勻度指數呈負相關關系,其中均勻度指數(Ea)和多樣性指數(D和PIE)與C/N均達到顯著關系(P<0.05)(圖7),物種均勻度和多樣性指數隨土壤N/P的增加呈降低趨勢(圖7)。
3 討論與結論
3.1凋落物對群落組分和物種多樣性的影響
凋落物的積累與分解是影響植被群落組分和生態系統物種多樣性的重要因子,包括凋落物的量和覆蓋時間兩方面[24-25]。凋落物量增加是草地封育初期最顯著的特征之一,但隨封育年限延長,凋落物量呈下降趨勢[26-27],然而也有相反結果的報道[28]。本研究與左萬慶等[26]和康博文等[27]研究結果一致,即隨封育年限的增加凋落物量呈下降趨勢(圖3),但由于3個不同封育年限樣地之間的凋落物量差異并不顯著,故認為不同封育年限群落組分和物種多樣性的改變與凋落物量無明顯關系。Xiong等[25]也指出凋落物覆蓋的時間越長,對物種多樣性的負效應越大,而凋落物量的影響并不顯著。本研究中,凋落物覆蓋對不同功能群植物的影響不同,莎草科植物受影響最小,豆科植物則最大,這與Wang等[29]的研究結果一致。本研究結果亦表明,均勻度指數和多樣性指數隨封育年限延長而下降,其主要原因可能與凋落物覆蓋時間有關。通常,覆蓋的時間越長,凋落物阻止種子到達土壤表面的可能性越大,從而抑制種子萌發和幼苗建植[30],降低物種多樣性。另外,凋落物覆蓋極大地消弱了群落底層的光照強度,導致一些喜光植物隨封育年限延長,其生存空間逐漸縮小而被淘汰,而光競爭能力強或耐蔭的物種得以存留。因此,由于凋落物覆蓋時間的負效應影響,最終導致群落組分發生變化、物種均勻度指數和多樣性指數隨封育年限延長而降低。

圖6 不同封育年限高寒草甸群落物種多樣性相關性Fig.6 Correlations of species diversity for different exclosure duration (year) in alpine meadow
封育年限/aExclosureduration土壤養分含量Soilnutrientcontent/(g/kg)土壤有機碳(SOC)Soilorganiccarbon土壤全氮(STN)Soiltotalnitrogen土壤全磷(STP)Soiltotalphosphorus土壤碳氮比(C/N)SoilC/Nratio土壤氮磷比(N/P)SoilN/Pratio254.50±0.90b4.77±0.13a0.79±0.03a11.45±0.25a6.60±0.19b750.46±1.25b4.29±0.20b0.69±0.02b11.78±0.32a6.48±0.23b1771.34±1.25a5.95±0.18a0.80±0.02a12.01±0.29a7.89±0.24a
表中字母不同表示差異顯著(P<0.05)

圖7 不同封育年限高寒草甸群落土壤養分比與多樣性的關系Fig.7 Relationship between soil nutrient ratio and species diversity for different exclosure duration in alpine meadow圖中沒有標明P值表示相關性不顯著(P>0.05)
然而,物種豐富度指數并沒有隨封育年限增加發生顯著變化,這可能與該高寒草甸存在一定的物種冗余有關,因為該群落存在許多重要值特別低(表1)且對初級生產力貢獻很小的物種[31],并且研究樣地在0.25 m2樣方內平均物種數目是內蒙古貝加爾針茅群落、大針茅群落、克氏針茅群落和小針茅群落物種數[32]的1.88—4.67倍。根據“冗余種假說”,該生態系統可能存在一些物種能夠適應因封育而改變的微環境。例如,群落下層優勢種矮嵩草,不同封育年限之間并沒有顯著變化;另外,美麗風毛菊較耐蔭,單位面積葉片的葉綠素含量和PSⅡ有效光化學量子產量要高于其它植物[33],葉片在地表伸張使其在與其它植物的競爭中占據更多的地表空間,其重要值封育7a和17a均顯著大于封育2a(P<0.05)。
3.2光競爭對群落組分和物種多樣性的影響
光照條件對草地植物群落的生長有重要作用[34],物種對光資源的競爭是影響植被群落結構和多樣性變化的因素之一[25]。根據Tilman[35]的資源比例假說,草地植物群落結構的改變與透光率有關,低透光率導致低的物種存活率。雖然本研究沒有對群落內的光照強度進行觀測,然而植株高度是反映植物對光資源競爭能力重要的代表性指標之一[31]。在本研究中,隨封育年限的延長,群落上層重要值最大的異針茅相對高度變化由封育2a的6.11%增加到封育7a和17a的8.13%和7.46%(P<0.05)(7a與17a之間不顯著)(圖3B)。由于異針茅的光競爭能力較強[36],其重要值(表1)和相對高度隨封育年限延長而增加,減弱其群落下層的光照,遮蔽了其他物種,其最終結果導致即長不高又不耐蔭的物種被排除掉,使物種多樣性下降,這與Chapin等[37]報道的結果一致。
3.3土壤養分對群落組分和物種多樣性的影響
土壤養分是維系地上和地下生物活動最基本的營養元素,也是影響群落組分和生物多樣性的重要因子[38],而封育后改變了地上植物與土壤之間的相互作用過程。多數研究表明,土壤養分隨封育年限的延長而增加[39,40],然而文海燕等[41]則指出,不同封育年限之間的土壤有機質和全氮的差異均不顯著。本研究中,封育2、7、17a SOC、STN和STP含量均表現為封育17a > 封育2a > 封育7a(表3),這與邱莉萍等[42]報道的半干旱地區的結果相似,其原因可能是草地封育后隨外界干擾的消失,植被加速了對原有土壤營養的利用,導致土壤養分降低[42],但隨封育時間的延長,由于凋落物的累積與分解導致土壤養分含量逐漸增加,因為土壤養分供應和養分循環的正反饋作用是通過凋落物的累積而加強[43]。
然而,土壤養分與群落組分和物種多樣性之間的關系較為復雜,因為不同功能群植物對土壤養分變化的響應不同[44]。楊曉霞等[36]和沈振西等[45]的研究表明,N和P元素均是高寒草甸植物生長重要的限制性營養元素,因此植物對土壤養分資源的競爭強度大,不同封育年限間土壤養分的變化必定引起群落組分的改變。本研究不同功能群總體重要值的變化表明,禾本科植物變化較大,如異針茅、羊茅對土壤養分變化的響應較為敏感(表1),這可能是由于它們對養分利用率較高的原因[45-46];莎草科和雜類草植物變化不明顯;而豆科植物卻顯著降低。土壤養分對物種多樣性的影響亦存在爭議,有研究表明隨土壤養分的增加,物種多樣性降低,但也有先增加后降低,或二者之間沒有相關性的報道[44]。本研究表明, SOC、STN、STP含量與α和β多樣性指數的相關性均不顯著,與左小安[47]的研究結果相同。然而,土壤C/N與多樣性指數和均勻度指數呈負相關關系,其中均勻度指數(Ea)和多樣性指數(D和PIE)與C/N均達到顯著關系(P<0.05)(圖7),說明物種多樣性主要是通過養分因子之間的耦合效應起作用,可能與土壤礦化作用有關,這與Small[48]的研究結果一致。另外,本研究表明物種均勻度和多樣性指數隨土壤N/P的增加呈降低趨勢(圖7),可能原因是N元素的增加導致優勢植物快速生長,進而對其它物種競爭排斥,導致物種多樣性降低,這與Roem[49]的研究結果一致。
3.4其他因素的可能影響
影響群落物種多樣性變化的因子非常復雜,是生物多樣性研究的主要內容。這些影響因子主要分為生物因子和環境因子,如光照、溫度、水分、養分、干擾以及物種之間的相互關系, 而群落生境差異可能是影響多樣性的主要原因,當前研究最多的環境因子是土壤因子和光照因子[50]。圍封年限的不同對其生境的改變亦不相同,對于天然草地,圍封后凋落物量、覆蓋時間及其分解對土壤因子和光照因子的改變是影響群落生境最顯著的因素,前文對此行了探討。然而,圍封引起的其他因子變化也可能對生物多樣性產生影響,如土壤水分、溫度以及物種間的相互作用等。
水分是影響植物生長發育及生物多樣性的重要因子,尤其是在干旱地區[51]。本研究3個圍封樣地相鄰,降水量相同,但封育樣地中的凋落物覆蓋可能通過截留部分降水、或減少土壤蒸發而改變水分收支和土壤水分[30]。然而,該研究樣地年均降水量達560 mm,特別是生長季中水分相對豐富,有研究指出水分不是該生態系統的限制因素[52]。另外,3個樣地間凋落物量無顯著差異(圖3),故可認為凋落物對降水的截留不會成為影響生物多樣性的直接原因。
溫度是控制植物生理活動和生化反應最重要的生態因子之一,能限制植物的分布,從而影響生物多樣性。氣候變暖對全球的生態系統和生物多樣性產生著顯著影響[53],特別是受低溫條件控制的高山生態系統對氣候變暖的響應尤為敏感[54],相關研究主要集中在全球尺度或較大的區域尺度上。圍封后由于凋落物覆蓋而改變群落底部溫度等微環境,進而影響生物多樣性,但目前尚未見到相關研究報道。本研究的3個樣地溫度背景相同,且樣地間凋落物量無顯著差異(圖3),但對于不同封育年限,凋落物覆蓋是否可導致樣地間的溫度差異及其對生物多樣性產生的影響仍有待于進一步探討。
極限相似性理論認為物種間相互作用可引起群落性狀趨異的構建模式[55],從而影響群落物種多樣性。封育后,凋落物的增加,牲畜采食、踐踏等干擾的排除,都是引起群落結構和土壤理化性質發生改變的重要因素,很多研究也報道了圍封對群落結構和土壤理化性質的影響[6,9- 15]。總之,圍封引起群落結構和土壤性質的改變使原有種間關系發生變化,也改變了群落中共存物種對資源的利用狀況,最終可能導致物種多樣性的改變。然而,物種間的相互作用關系非常復雜,欲探明其種間關系對生物多樣性的影響還需長期的、更深入的實驗數據提供依據。
3.5結論
本文探討了2、7、17a不同封育年限對青藏高原東北部高寒草甸群落組分及物種多樣性的影響,結果表明,隨封育年限的延長,禾本科植物的總體重要值增加最明顯,特別是異針茅最為顯著,成為17a封育樣地中群落上層的單優勢種;而豆科植物總體重要值顯著降低;莎草科和雜類草植物變化不明顯,說明封育對群落的物種組成及重要值具有較大影響。物種多樣性指數和均勻度指數均隨封育年限的延長呈下降趨勢,且封育前期(2a與7a之間)變化大于后期(7a與17a之間)。凋落物的存在影響植物的種子萌發、幼苗定居以及物種間的相互關系,但3個樣地間凋落物量的差異并不顯著,說明不同封育年限樣地間物種多樣性的差異與凋落物量無關,而與凋落物覆蓋時間有關。隨封育年限的延長,凋落物分解參與調節的土壤養分含量及其比例發生改變,其中土壤C/N和N/P與群落物種多樣性呈負相關關系,說明土壤養分之間的耦合作用影響高寒草甸群落組分及其物種多樣性。本研究說明,封育雖然是草地恢復的有效措施之一,但就封育年限對物種多樣性的影響而言,該地區高寒草甸生態系統不宜進行長期封育,可采取適度的放牧或輪牧管理措施,保護其物種多樣性。由于影響物種多樣性變化的因子非常復雜,在以后的研究中還需進一步加強研究封育年限對群落結構及其生物多樣性影響的生態學機制,同時加強封育年限及其生物多樣性變化對生態系統功能影響的研究。
[1]徐玲玲, 張憲洲, 石培禮, 于貴瑞. 青藏高原高寒草甸生態系統表觀量子產額和表觀最大光合速率的確定. 中國科學(D輯: 地球科學), 2004, 34(S2): 125- 130.
[2]孫鴻烈, 鄭度, 姚檀棟, 張鐿鋰. 青藏高原國家生態安全屏障保護與建設. 地理學報, 2012, 67(1): 3- 12.
[3]王長庭, 龍瑞軍, 王啟蘭, 曹廣民, 施建軍, 杜巖功. 放牧擾動下高寒草甸植物多樣性、生產力對土壤養分條件變化的響應. 生態學報, 2008, 28(9): 4144- 4152.
[4]馬玉壽, 郎百寧, 王啟基.“黑土型”退化草地研究工作的回顧與展望. 草業科學, 1999, 16(2): 5- 9.
[5]韓龍, 郭彥軍, 韓建國, 郭蕓江, 唐華. 不同刈割強度下羊草草甸草原生物量與植物群落多樣性研究. 草業學報, 2010, 19(3): 70- 75.
[6]喬春連, 王基恒, 葛世棟, 陳懂懂, 趙亮, 李英年, 徐世曉. 圍封和放牧條件下高寒矮嵩草草甸土壤性質的比較. 草業科學, 2012, 29(3): 341- 345.
[7]姜世成, 周道瑋. 過牧、深翻及封育三種方式對退化羊草草地的影響. 中國草地, 2002, 24(5): 5- 9.
[8]江小蕾, 張衛國, 楊振宇, 王剛. 不同干擾類型對高寒草甸群落結構和植物多樣性的影響. 西北植物學報, 2003, 23(9): 1479- 1485.
[9]吳建波, 包曉影, 李潔, 趙念席, 高玉葆. 不同圍封年限對典型草原群落及大針茅種群特征的影響. 草地學報, 2010, 18(4): 490- 495.
[10]單桂蓮, 徐柱, 寧發, 馬玉寶, 李臨杭. 圍封年限對典型草原群落結構及物種多樣性的影響. 草業學報, 2008, 17(6): 1- 8.
[11]閆玉春, 唐海萍, 常瑞英, 劉亮. 典型草原群落不同圍封時間下植被、土壤差異研究. 干旱區資源與環境, 2008, 22(2): 145- 151.
[12]呂世海, 馮長松, 高吉喜, 盧欣石. 呼倫貝爾沙化草地圍封效應及生物多樣性變化研究. 草地學報, 2008, 16(5): 442- 447.
[13]苗仁輝, 蔣德明, 王永翠. 科爾沁沙質草地封育過程中的植被變化及其機制. 干旱區研究, 2013, 30(2): 264- 270.
[14]晁增國, 汪詩平, 徐廣平, 胡宜剛, 張振華. 圍封對退化矮嵩草草甸群落結構和主要種群空間分布格局的影響. 西北植物學報, 2009, 28(11): 2320- 2326.
[15]蘇淑蘭, 李洋, 王立亞, 郭丁, 康海軍, 李旭東, 傅華. 圍封與放牧對青藏高原草地生物量與功能群結構的影響. 西北植物學報, 2014, 34(8): 1652- 1657.
[16]趙景學, 曲廣鵬, 多吉頓珠, 尚占環. 藏北高寒植被群落物種多樣性與土壤環境因子的關系. 干旱區資源與環境, 2011, 25(6): 105- 108.
[17]苗福泓, 郭雅婧, 繆鵬飛, 郭正剛, 沈禹穎. 青藏高原東北邊緣地區高寒草甸群落特征對封育的響應. 草業學報, 2012, 21(3): 11- 16.
[18]李英年, 趙新全, 曹廣民, 趙亮, 王勤學. 海北高寒草甸生態系統定位站氣候、植被生產力背景的分析. 高原氣象, 2004, 23(4): 558- 567.
[19]周華坤, 周立, 劉偉, 王啟基, 趙偉, 周彥艷. 封育措施對退化與未退化矮嵩草草甸的影響. 中國草地, 2003, 25(5): 17- 22.
[20]周杰, 趙洪, 袁曉霞, 楊玉婷, 羅開嘉, 郭正剛. 青藏高原邊緣區高寒草甸植物群落的特征. 草業科學, 2013, 30(7): 1066- 1070.
[21]陳芙蓉, 程積民, 劉偉, 朱仁斌, 楊曉梅, 趙新宇, 蘇紀帥. 不同干擾對黃土區典型草原物種多樣性和生物量的影響. 生態學報, 2013, 33(9): 2856- 2866.
[22]劉維暐, 王杰, 王勇, 楊帆. 三峽水庫消落區不同海拔高度的植物群落多樣性差異. 生態學報, 2012, 32(17): 5454- 5466.
[23]白永飛, 邢雪榮, 許志信, 李德新. 內蒙古高原針茅草原群落β多樣性研究. 應用生態學報, 2000, 11(3): 408- 412.
[24]Wardle D A, Bonner K I, Nicholson K S. Biodiversity and plant litter: experimental evidence which does not support the view that enhanced species richness improves ecosystem function. Oikos, 1997, 79(2): 247- 258.
[25]Xiong S J, Nilsson C. The effects of plant litter on vegetation: a meta-analysis. Journal of Ecology, 1999, 87(6): 984- 994.
[26]左萬慶, 王玉輝, 王風玉, 師廣旭. 圍欄封育措施對退化羊草草原植物群落特征影響研究. 草業學報, 2009, 18(3): 12- 19.
[27]康博文, 劉建軍, 侯琳, 李勝功. 蒙古克氏針茅草原生物量圍欄封育效應研究. 西北植物學報, 2006, 26(12): 2540- 2546.
[28]闞雨晨, 王堃, 王宇通, 黃欣穎, 邵新慶. 典型草原封育過程中植物凋落物的變化動態. 草原與草坪, 2011, 31(4): 25- 29.
[29]Wang C T, Long R J, Wang Q L, Liu W, Jing Z C, Zhang L. Fertilization and litter effects on the functional group biomass, species diversity of plants, microbial biomass, and enzyme activity of two alpine meadow communities. Plant and Soil, 2010, 331(1/2): 377- 389.
[30]李強, 周道瑋, 陳笑瑩. 地上枯落物的累積、分解及其在陸地生態系統中的作用. 生態學報, 2014, 34(14): 3807- 3819.
[31]李曉剛. 模擬放牧和土壤資源獲得性對高寒草甸物種多樣性、功能多樣性與初級生產力關系的影響[D]. 西安: 陜西師范大學, 2012.
[32]白永飛, 許志信, 李德新. 內蒙古高原針茅草原群落α多樣性研究. 生物多樣性, 2000, 8(4): 353- 360.
[33]師生波, 李惠梅, 王學英, 岳向國, 徐文華, 陳桂琛. 青藏高原幾種典型高山植物的光合特性比較. 植物生態學報, 2006, 30(1): 40- 46.
[34]Tilman D. Species richness of experimental productivity gradients: how important is colonization limitation? Ecology, 1993, 74(8): 2179- 2191.
[35]Tilman D. The resource-ratio hypothesis of plant succession. The American Naturalist, 1985, 125(6): 827- 852.
[36]楊曉霞, 任飛, 周華坤, 賀金生. 青藏高原高寒草甸植物群落生物量對氮、磷添加的響應. 植物生態學報, 2014, 38(2): 159- 166.
[37]Chapin III F S, Shaver G R, Giblin A E, Nadelhoffer K J, Laundre J A. Responses of arctic tundra to experimental and observed changes in climate. Ecology, 1995, 76(3): 694- 711.
[38]Huang P M, Wang S L, Tzou Y M, Huang Y B, Weng B Q, Zhuang S Y, Wang M K. Physicochemical and biological interfacial interactions: impacts on soil ecosystem and biodiversity. Environmental Earth Sciences, 2013, 68(8): 2199- 2209.
[39]白于. 黃土區不同封育時期天然草地根系與土壤特性[D]. 楊凌: 西北農林科技大學, 2014.
[40]何玉惠, 趙哈林, 劉新平, 趙學勇, 李玉強. 封育對沙質草甸土壤理化性狀的影響. 水土保持學報, 2008, 22(2): 159- 161, 181- 181.
[41]文海燕, 趙哈林, 傅華. 開墾和封育年限對退化沙質草地土壤性狀的影響. 草業學報, 2005, 14(1): 31- 37.
[42]邱莉萍, 張興昌, 程積民. 不同封育年限草地土壤有機質組分及其碳庫管理指數. 植物營養與肥料學報, 2011, 17(5): 1166- 1171.
[43]Aerts R, de Caluwe H. Nutritional and plant-mediated controls on leaf litter decomposition ofCarexspecies. Ecology, 1997, 78(1): 244- 260.
[44]王長庭, 龍瑞軍, 劉偉, 王啟蘭, 張莉, 吳鵬飛. 高寒草甸不同群落類型土壤碳分布與物種多樣性、生物量關系. 資源科學, 2010, 32(10): 2022- 2029.
[45]沈振西, 周興民, 陳佐忠, 周華坤. 高寒矮嵩草草甸植物類群對模擬降水和施氮的響應. 植物生態學報, 2002, 26(3): 288- 294.
[46]曹廣民, 杜巖功, 梁東營, 王啟蘭, 王長庭. 高寒嵩草草甸的被動與主動退化分異特征及其發生機理. 山地學報, 2008, 25(6): 641- 648.
[47]左小安, 趙學勇, 趙哈林, 李玉強, 郭軼瑞, 趙玉萍. 科爾沁沙質草地群落物種多樣性、生產力與土壤特性的關系. 環境科學, 2007, 28(5): 945- 951.
[48]Small C J, McCarthy B C. Relationship of understory diversity to soil nitrogen, topographic variation, and stand age in an eastern oak forest, USA. Forest Ecology and Management, 2005, 217(2/3): 229- 243.
[49]Roem W J, Berendse F. Soil acidity and nutrient supply ratio as possible factors determining changes in plant species diversity in grassland and heathland communities. Biological Conservation, 2000, 92(2): 151- 161.
[50]汪殿蓓, 暨淑儀, 陳飛鵬. 植物群落物種多樣性研究綜述. 生態學雜志, 2001, 20(4): 55- 60.
[51]李新榮, 張景光, 劉立超, 陳懷順, 石慶輝. 我國干旱沙漠地區人工植被與環境演變過程中植物多樣性的研究. 植物生態學報, 2000, 24(3): 257- 261.
[52]李紅琴, 李英年, 張法偉, 劉曉琴, 吳啟華, 毛紹娟. 高寒草甸植被生產量年際變化及水分利用率狀況. 冰川凍土, 2013, 35(2): 475- 482.
[53]Walther G R, Post E, Convey P, Menzel A, Parmesan C, Menzel A, Parmesan C, Beebee T J, Fromentin J M, Guldberg O H, Bairlein F. Ecological responses to recent climate change. Nature, 2002, 416(6879): 389- 395.
[54]K?rner C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlín & Heidelberg: Springer Verlag, 1999: 297- 298.
[55]Pillar V D, da S Duarte L, Sosinski E E, Joner F. Discriminating trait-convergence and trait-divergence assembly patterns in ecological community gradients. Journal of Vegetation Science, 2009, 20(2): 334- 348.
Effects of exclosure duration on the community structure and species diversity of an alpine meadow in the Qinghai-Tibet Plateau
LIU Xiaoqin1, ZHANG Xiang1, ZHANG Lifeng1, LI Yingnian2, ZHAO Liang2, XU Shixiao2, LI Hongqin2, MA Rongrong1, NIU Ben1, GAO Yubao1, GU Song1,*
1CollegeofLifeSciences,NankaiUniversity,Tianjin300071,China2NorthwestPlateauInstituteofBiology,ChineseAcademyofSciences,Xining810008,China
The Qinghai-Tibet Plateau is one of the richest areas in terms of biodiversity in China. The alpine meadow is the most widespread vegetation type on this plateau and plays an important role in the carbon cycle, conservation of biodiversity, and livestock husbandry. However, climate change and human activities have greatly affected this already fragile ecosystem. Although exclosure, one of the effective measures, has been widely applied in grassland restoration, the impact of exclosure duration on community structure and species diversity is of particular interest. Therefore, determining the influence of exclosure duration on the alpine meadow is important for the development of sustainable livestock husbandry. We examined the community structure and species diversity in patches ofKobresiahumilisalpine meadow, which had been enclosed for 2, 7, or 17 years. We have discussed possible reasons for the changes in community structure and species diversity. The results indicated the following: 1) With increase in exclosure duration, the percentage of grass species in the upper community increased significantly, especially ofStipaaliena. The percent change was 13.87% for 2-year, 21.76% for 7-year, and 23.95% for 17-year exclosure. The percent change of leguminous species significantly decreased, whereas sedge and forbs species exhibited no obvious change in exclosure duration. 2) The α-biodiversity, Patrick index, and Margalef index of the plant communities indicated no significant changes in exclosure duration. In contrast, the Simpson index, Shannon-Wiener index, probability of interspecific encounter index, Alatalo index, and Pielou index decreased in exclosure duration (2-year > 7-year > 17-year enclosures); there were significant differences between the 2-year and 7-year exclosures and between the 2-year and 17-year exclosures (P<0.05). 3) Theβ-biodiversity, Cody index, and Morisita-Horn similarity index indicated that differences inspecies were greater between 2-year and 7-year exclosures, as well as 2-year and 17-year exclosures, than that between 7-year and 17-year exclosures. 4) The amount of plant litter decreased in exclosure duration, but differences were not significant. Litter coverage time appeared to be one of the reasons for the decrease in species diversity because the litter layer could prevent the seeds from falling to the ground. 5) Soil organic carbon (SOC), soil total nitrogen (STN), and soil total phosphorus (STP) increased in exclosure duration due to litter decomposition, but we observed no effect on species diversity. We found that the ratio of soil carbon to nitrogen (C/N) showed significantly negative correlation with the Alatalo index, Simpson index, and probability of interspecific encounter index (P<0.05), indicating that the coupling relationship between soil carbon and nitrogen was possibly the most important factor influencing species diversity. Our results suggested that species diversity decreased in exclosure duration because of the effects of litter accumulation, litter decomposition, and plant competition for light. The influence of exclosure on species diversity in the early stage was greater than that in the later stage, and thus, long-term exclosure was unnecessary for species diversity in this alpine meadow. We hope that our study will provide a scientific basis for biodiversity protection, sustainable utilization, and management of alpine meadow ecosystems.
Qinghai-Tibet Plateau; richness index; evenness index; diversity index;βdiversity
國家自然科學基金項目(31070433, 31570478, 31270523, 31300385)
2015- 01- 30; 網絡出版日期:2015- 12- 03
Corresponding author.E-mail: songgu@nankai.edu.cn
10.5846/stxb201501300251
劉曉琴,張翔,張立鋒,李英年,趙亮,徐世曉,李紅琴,馬榮榮,牛犇,高玉葆,古松.封育年限對高寒草甸群落組分和物種多樣性的影響.生態學報,2016,36(16):5150- 5162.
Liu X Q, Zhang X, Zhang L F, Li Y N, Zhao L, Xu S X, Li H Q, Ma R R, Niu B, Gao Y B, Gu S.Effects of exclosure duration on the community structure and species diversity of an alpine meadow in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2016,36(16):5150- 5162.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
