實時熒光定量PCR技術監測腌制麻竹筍中乳酸乳球菌動態變化
2016-11-11 08:15:13夏雪娟葉秀娟吳金松闞建全南大學食品科學學院重慶40075重慶市特色食品工程技術研究中心重慶40075
食品科學 2016年4期
關鍵詞:標準
夏雪娟,鄭 炯,2,,葉秀娟,吳金松,闞建全,2(.西 南大學食品科學學院,重慶 40075;2.重慶市特色食品工程技術研究中心,重慶 40075)
實時熒光定量PCR技術監測腌制麻竹筍中乳酸乳球菌動態變化
夏雪娟1,鄭 炯1,2,*,葉秀娟1,吳金松1,闞建全1,2
(1.西 南大學食品科學學院,重慶 400715;2.重慶市特色食品工程技術研究中心,重慶 400715)
采用實時熒光定量聚合酶鏈式反應(quantitative real-time polymerase chain reaction,qRT-PCR)技術定量監測6 g/100 mL鹽質量濃度腌制麻竹筍中乳酸乳球菌(Lactococcus lactis)的動態變化。經乳酸乳球菌標準菌株基因組DNA提取、標準陽性質粒制備、標準曲線繪制、各時期竹筍腌制發酵液中細菌基因組DNA提取和乳酸乳球菌qRTPCR特異性擴增,對腌制液中乳酸乳球菌進行定量檢測。結果表明,在腌制過程中(0~63 d),隨著腌制時間的延長,乳酸乳球菌含量逐漸升高,在腌制14 d時達到最大值(4.63×108copies/?L),與0 d(2.41×102copies/?L)相比增加了6 個數量級,而后濃度緩慢降低,在腌制63 d時濃度為5.02×106copies/?L。qRT-PCR技術為定量監測腌制麻竹筍中微生物的動態變化提供了一條可靠、快速的有效途徑。
大葉麻竹筍;腌制;實時熒光定量PCR;乳酸乳球菌;動態變化
大葉麻竹筍(Dendrocalamus latiflorus)是著名的高產型竹筍,廣泛分布于我國南亞熱帶和熱帶地區[1]。腌制大葉麻竹筍加工簡易,成本低廉,易于保存,且具有較高的營養價值、獨特的風味,深受消費者的喜愛[2]。在腌制發酵過程中,麻竹筍會形成多種風味化合物,使其具有特殊的風味和營養價值[3]。而在風味形成過程中,除了其本身的風味外,其他的風味形成途徑均與微生物的發酵作用有關[4],因此,了解麻竹筍腌制過程中優勢微生物的動態變化對于研究腌制麻竹筍的風味品質變化具有重要作用。
乳酸菌具有良好的感官特性和防腐特性,在蔬菜發酵過程中發揮著重要作用[5]。其中,乳酸乳球菌(Lactococcus lactis)可形成檸檬酸,參與乙酰風味化合物的合成,并參與發酵初始風味的形成[6]。近年來,國內外諸多學者從腌制發酵蔬菜中分離鑒定出乳酸乳球菌,并對其益生作用進行研究。如付琳琳等[7]采用聚合酶鏈式反應-變性梯度凝膠電泳技術 從不同家庭制作的成熟期的泡菜液中鑒定出乳酸乳球菌。尹軍霞等[8]研究了分離自酸菜汁的乳酸乳球菌的體外去除膽固醇特性。連元元等[9]從泡菜中提取并保存了一株泡菜乳酸乳球菌,并發現其具有較高的耐鎘能力。Seema等[10]從腌制山藥中分離出一株乳酸乳球菌并對其益生特性進行初步研究。Aso等[11]從日本傳統“津田蕪菁”泡菜中分離出一株乳酸乳 球菌,并對其產Nisin Z的能力進行探討。本課題組前期的研究也表明乳酸乳球菌是腌制麻竹筍中的優勢菌群,但其 含量及在腌制加工過程中的動態變化還有待進一步研究[4]。
實時熒光定量聚合酶鏈式反應(quantitative real-time polymerase chain reaction,qRT-PCR)技術可定量研究微生物菌落結構組成及數量變化,為全面、快速、準確地分析鑒定食品[12-15]中的各種微生物提供了一種嶄新的技術工具和平臺。近年來,也有部分學者采用qRT-PCR技術對食品中的乳酸乳球菌進行定量研究,如Grattepanche[6]和Achilleos[16]等分別采用qRT-PCR技術對混合發酵乳和芝士中的乳酸乳球菌進行鑒定。而有關麻竹筍腌制過程中乳酸乳球菌的qRT-PCR研究尚未見報道。因此,本實驗采用qRT-PCR技術對腌制麻竹筍中的乳酸乳球菌變化進行定量監測,以期為腌制麻竹筍中風味物質的形成及變化機理研究提供實驗依據。
1 材料與方法
1.1 材料與試劑
腌制麻竹筍:大葉麻竹筍原料采于重慶市北碚區施家梁鎮大葉麻竹筍種植基地,將麻竹筍洗凈、切分、漂燙、瀝干、冷卻后,按傳統加工方法加鹽(6 g/100 mL)在室溫條件下腌制。從腌制當天開始取樣,第3天取一次樣,之后每隔一周取一次樣,共取11 次樣(0、3、7、14、21、28、35、42、49、56 d和63 d),取樣完成時產品已基本成熟。樣品-20 ℃冰箱保存,至最后一次取樣完成后統一進行DNA提取。
乳酸乳球菌乳亞種(Lactococcus lactis subsp. lactis)CGMCC1.1936 中國普通微生物菌種保藏管理中心。
MRS培養基用試劑及配制方法參照文獻[17];基因組DNA提取和克隆用試劑及配制方法參考文獻[18];細菌基因組DNA提取試劑盒(DP302)、溶菌酶、瓊脂糖、GeneGreen核酸染料(RT210)、瓊脂糖凝膠回收試劑盒(DP209)、pGM-T克隆試劑盒(VT202)、DH5α感受態細胞(CB101-03)、質粒小提試劑盒(DP103) 天根生化科技(北京)有限公司;TaKaRa Taq擴增體系套裝(R001A)、SYBR Premix Ex TaqTMⅡ(DRR820A) 寶生物工程(大連)有限公司;毛細管 上海大菱制塑有限公司;Lambda DNA/Hind ⅢMarker(NMW009)、100 bp ladder Ⅱ(NMW004)北京鼎國昌盛生物技術有限責任公司;實驗所用引物由生工生物工程(上海)股份有限公司合成;化學試劑均為分析純。
1.2 儀器與設備
S1000伯樂1000系列高性能PCR儀、164-5050基礎電源電泳儀 美國Bio-Rad公司;Syngene GeneGenius凝膠成像系統 基因科技(上海)有限公司;LightScanner 32熒光定量PCR儀 美國Idaho公司;NanoDrop 2000微量紫外分光光度計 美國Thermo公司;SIM-F140AY65型制冰機 三洋電機國際貿易有限公司;SW-CJ-1F型單人雙面凈化工作臺 蘇州凈化設備有限公司;1-15PK小型臺式離心機 德國Sigma公司;ES-315高壓滅菌鍋日本Tomy公司;HWS-26電熱恒溫水浴鍋、DHP-9272電熱恒溫培養箱 上海齊欣科學儀器有限公司。
1.3 方法
1.3.1 標準品的制備及標準曲線的建立
1.3.1.1 標準菌株模板DNA的制備
將乳酸乳球菌按照標準菌株培養條件接種至MRS液體培養基中,30 ℃恒溫振蕩培養過夜,經2 次活化后采用天根細菌基因組DNA提取試劑盒提取細菌基因組DNA。用溶菌酶對菌體進行破壁處理,提取結束時用80 μL TE buffer將DNA溶出。經0.8%瓊脂糖凝聚電泳,GeneGreen染色檢測后,-20 ℃保存備用,目標片段應在23 kb左右[19]。
1.3.1.2 目的片段的擴增純化
采用特異性引物對標準菌株基因組DNA進行常規PCR擴增,用以制備標準陽性質粒[20]。正向引物L. lactis-F:5’-TGA AGA ATT GAT GGA ACT CG-3’,反向引物L. lactis-R:5’-CAT TGT GGT TCA CCG TTC-3’,目標片段約為126 bp[16]。擴增體系為:5 μL DNA模板,正向和反向引物各0.25 μL,10×PCR buffer(Mg2+plus)10 μL,dNTP Mixture 3 μL,Taq DNA聚合酶0.5 μL,滅菌超純水補足至50 μL。反應程序為:94 ℃預變性5 min,94 ℃變性40 s,56 ℃退火40 s,72 ℃延伸1 min,35 個循環,72 ℃延伸7 min,4 ℃保存。擴增產物經3%瓊脂糖凝膠電泳檢測后,用無菌手術刀在紫外燈下切取目標片段,并采用瓊脂糖凝膠回收試劑盒對其進行純化回收。回收產物經瓊脂糖凝膠電泳檢測純度后-20 ℃保存備用。
1.3.1.3 標準陽性質粒的制備[21]
使用天根pGM-T克隆試劑盒將已純化標準菌株目的片段與質粒進行連接,并將4 ?L連接反應物加入至50 ?L DH5α感受態細胞中進行轉化。利用藍白斑篩選法挑選白色陽性克隆菌落后采用質粒小提試劑盒提取質粒,具體操作詳見試劑盒說明書。
為避免質粒假陽性,采用通用引物T7(5’-TAA TAC GAC TCA CTA TAG GG-3’)和SP6(5’-ATT TAG GTG ACA CTA TAG-3’)對提取質粒進行擴增,擴增產物用做熒光定量PCR反應的標準品以繪制標準曲線。擴增體系同1.3.1.2節,擴增程序為:94 ℃預變性5 min,94 ℃變性40 s,55 ℃退火40 s,72 ℃延伸1 min,35 個循環,72 ℃延伸7 min,4 ℃保存。擴增產物用3%的瓊脂糖凝膠電泳和微量紫外分光光度計檢測其濃度與純度后,送至生工生物工程(上海)股份有限公司測序。
1.3.1.4 qRT-PCR條件優化
以L. lactis-F和L. lactis-R為引物,以得到最小的Ct值(熒光信號達到設定閾值時所經歷的循環數)、最大的熒光值和熔解曲線不產生非特異性峰為指標,對乳酸乳球菌qRT-PCR的退火溫度、退火時間、引物濃度以及熒光數據收集溫度等條件進行優化,確定擴增的最佳條件[22]。
1.3.1.5 標準曲線的繪制[12,23]
由微量紫外分光光度計測定的標準陽性質粒PCR產物濃度,結合產物測序得到的片段大小,根據公式(1)計算出PCR產物中所含的初始拷貝數。

稀釋質粒擴增產物,觀察不同濃度模板對擴增效率及熒光吸收強度的影響,從而確定合適的模板濃度范圍。在合適的濃度范圍內以無菌超純水連續10 倍梯度稀釋,將其作為陽性模板,按照優化后的條件進行熒光定量PCR反應,得到不同濃度梯度模板的PCR擴增曲線圖。再以不同濃度陽性模板的拷貝數對數為橫坐標,Ct值為縱坐標繪制出標準曲線,得到標準菌株的線性回歸方程及相關系數(R2)。
1.3.2 各時期竹筍腌制發酵液中乳酸乳球菌含量的qRT-PCR檢測
取3 mL各時期竹筍腌制發酵液,10 000 r/min離心1 min,棄上清液,加入180 ?L 20 mg/mL的溶菌酶,37 ℃處理2 h后采用天根細菌基因組DNA提取試劑盒提取細菌基因組DNA。DNA用80 μL TE buffer溶出后置于-20 ℃冰箱保存備用。采用優化好的qRT-PCR條件和體系對各時期竹筍腌制發酵液細菌基因組DNA中乳酸乳球菌含量進行檢測。
1.4 數據處理
實驗數據采用Origin(Version 8.6)和Excel(Version 2013)軟件進行處理與分析。
2 結果與分析
2.1 實時熒光定量PCR方法的建立
2.1.1 標準菌株模板DNA提取結果
乳酸乳球菌標準菌株基因組DNA電泳結果見圖1,菌株基因組DNA在目標位置處有亮帶,且無拖尾,說明基因組質量較好。
圖1 乳酸乳球菌DNA凝膠電泳圖Fig.1 Agarose gel electrophoresis of L. lactis genomic DNA
2.1.2 標準陽性質粒制備結果

圖2 乳酸乳球菌常規PCR及陽性質粒PCR產物電泳圖Fig.2 Agarose gel electrophoresis of PCR amplified products of L. lactis and its positive plasmids
以提取的乳酸乳球菌基因組DNA為模板,經特異性引物常規PCR擴增后,對目的片段進行連接、轉化、克隆,再采用通用引物T7和SP6對質粒進行擴增。常規PCR產物和質粒PCR產物的瓊脂糖凝膠電泳檢測結果如圖2所示。
由圖2可知,常規PCR和質粒PCR擴增產物條帶均清晰,無雜帶、無拖尾,且常規PCR產物片段大小在126 bp左右,與預期結果一致。由于質粒擴增得到的片段是目的片段與pGM-T載體的部分序列(約145 bp)的連接產物,因此菌株的陽性質粒擴增產物大小約為271 bp,與圖中一致。
經紫外分光光度計檢測,菌株陽性質粒擴增產物的OD260nm為5.266±0.220,OD280nm為2.846±0.072,O D260nm/O D280nm為1.8 5±0.0 3,質量濃度為(263.3±11.0) ng/μL,質量較好。經序列分析,除去兩端質粒片段,得到127 bp的目的片段,與預期結果相一致。且經BLAST比對(http://blast.ncbi.nlm.nih.gov/Blast. cgi)所得序列與乳酸乳球菌乳亞種(Lactococcus lactis subsp. lactis)的核苷酸序列同源性為100%,說明所構建的重組質粒含有目的片段,可作為標準品用于qRT-PCR中。2.1.3 qRT-PCR反應條件優化結果經優化,擴增采用25 ?L的反應體系,反應體系見表1。

表1 qRT-PCR反應體系Table 1 Reaction system for qRT-PCR amplification
利用SYBR Green Ⅰ染料法進行實時監測,選擇兩步法PCR擴增程序進行擴增。反應程序為:第一步:95 ℃預變性30 s,20 ℃/s,1 個循環,不采集信號;第二步:95 ℃變性5 s,20 ℃/s,60 ℃退火20 s,20 ℃/s,每個循環采集熒光信號,50 個循環。熔解程序為:95 ℃、0 s,20 ℃/s;65 ℃、15 s,20 ℃/s;95 ℃、0 s,0.1 ℃/s,持續采集信號;50 ℃冷卻30 s,20 ℃/s。
2.1.4 標準曲線的建立
對陽性標準品進行系列稀釋(10-2、10-4、10-6、10-8、10-10),采用優化的條件進行qRT-PCR,不同濃度梯度陽性模板的PCR擴增熒光曲線圖如圖3A所示,其熔解曲線見圖3B。由圖3可知,各濃度擴增產物均在(78.7±0.4) ℃出現熒光峰值,無雜峰,說明產物均一性良好,無非特異擴增[24]。以相對拷貝數的對數為橫坐標,以Ct值為縱坐標,得出菌株標準曲線。回歸方程為:y=-3.868x+45.142,R2=0.998 5,線性關系良好。
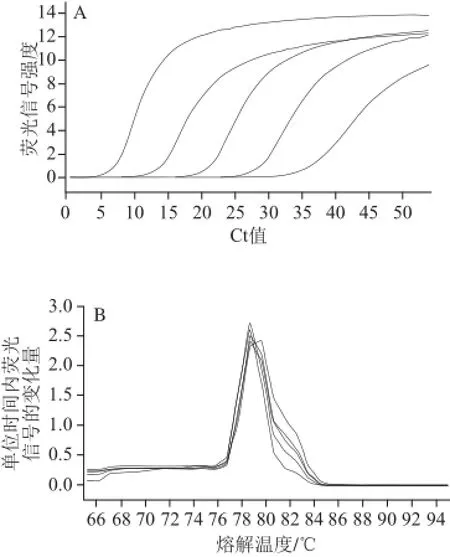
圖3 乳酸乳球菌熒光定量PCR熒光曲線(A)與熔解曲線(B)Fig.3 Quantitative PCR fluorescence and melting curves for Lactococcus lactis
2.2 各時期竹筍腌制發酵液中乳酸乳球菌含量檢測結果
由于3 mL腌制發酵液中細菌較少,細菌基因組DNA條帶較暗,故未提供其電泳結果。采用優化好的qRTPCR條件和體系對各時期竹筍腌制發酵液中乳酸乳球菌含量進行檢測,根據熔解曲線分析PCR產物的特異性,根據樣品的擴增曲線可計算得出Ct值,再帶入標準曲線方程中,可得各樣品中乳酸乳球菌的拷貝數對數值(lgC)見圖4。使用Power函數對縱坐標進行換算,得出各時期竹筍腌制發酵液中乳酸乳球菌的拷貝數,在腌制0~3 d時,乳酸乳球菌含量略有上升,但濃度較低(分別為2.41×102copies/?L和1.78×103copies/?L)。隨著腌制時間的延長,乳酸乳球菌濃度迅速升高,在腌制7 d時濃度為3.47×105copies/?L,而在腌制14 d時達到最大值,含量為4.63×108copies/?L,與腌制0 d相比增加了6 個數量級。之后濃度緩慢下降,在腌制21~63 d期間,乳酸乳球菌濃度從1.40×108copies/?L波動下降至5.02×106copies/?L。

圖4 麻竹筍腌制液中乳酸乳球菌的動態變化Fig.4 Changes in of Lactococcus lactis concentration in pickled ma bamboo shoots during pickling
3 討論與結論
本研究采用qRT-PCR技術對6 g/100 mL鹽質量濃度腌制麻竹筍發酵液中乳酸乳球菌含量的動態變化進行監控。經標準菌株基因組DNA提取、擴增、克隆、制備出標準陽性質粒,再經擴增獲得陽性模板,再用優化后的qRT-PCR技術制備出標準曲線。提取各時期竹筍腌制發酵液中細菌基因組DNA,對其中乳酸乳球菌的含量進行qRT-PCR定量檢測。結果表明,隨著腌制時間的延長,乳酸乳球菌的含量逐漸升高,在腌制14 d時達到最大值(4.63×108copies/?L),而后濃度緩慢降低,在腌制63 d時濃度為5.02×106copies/?L。
乳酸乳球菌(L. l a c t i s)歸屬于硬壁菌門(F i r m i c u t e s)桿菌綱(B a c i l l i)乳桿菌目(Lactobacillales)鏈球菌科(Streptococcaceae)乳球菌屬(Lactococcus)[25]。Aso[11]和Harri[26]等研究表明乳酸乳球菌能在發酵初期產生細菌素Nisin,抑制存在于鹽水中的一些食源性致病菌和一些革蘭氏陰性菌生長,且乳酸乳球菌為同型發酵,通過糖酵解途徑產生乳酸。Xiong Tao等[27]研究表明,乳酸乳球菌在酸菜腌制過程中生長速度快,且產酸能力強,但耐酸能力差。本課題組之前的研究也表明,隨著發酵時間的延長,麻竹筍發酵液pH值逐漸降低[2]。因此,可推測本研究中乳酸乳球菌濃度先升高再降低的原因是由于發酵開始時乳酸乳球菌產酸、產細菌素,抑制了其他細菌的生長,使得它的濃度迅速升高,而隨著發酵時間的延長,發酵液pH值降低,乳酸乳球菌活力下降。Xiong Tao等[28]研究表明,在泡菜發酵過程中,乳酸乳球菌在發酵初期慢慢出現,在后期死亡,與本研究相一致。
綜上所述,qRT-PCR技術為腌制麻竹筍中的微生物動態變化的定量監測提供了一條可靠、快速的有效途徑。本實驗僅對腌制大葉麻竹筍中乳酸乳球菌的定量變化進行了檢測,進一步的研究可對腌制麻竹筍中不同微生物進行定量檢測,以探討腌制發酵液中微生物的交互作用對其風味物質的形成作用,為麻竹筍的風味形成機理和腌制工藝的改進提供一定的參考依據。
[1] SANTOSH S, LALIT M B, POONAM S, et al. Bamboo shoot processing: food quality and safety aspect[J]. Trends in Food Science and Technology, 2010, 21(4): 181-189. DOI:10.1016/ j.tifs.2009.11.002.
[2] 陳光靜, 汪莉莎, 鄭炯, 等. 食鹽質量濃度對大葉麻竹筍腌制過程中品質特性的影響[J]. 食品科學, 2013, 34(15): 48-52. DOI:10.7506/ spkx1002-6630-201315010.
[3] 鄭炯, 宋家芯, 陳光靜, 等. 頂空-固相微萃取-氣質聯用法分析腌制麻竹筍揮發性成分[J]. 食品科學, 2013, 34(18): 193-196. DOI:10.7506/spkx1002-6630-201318039.
[4] 鄭炯, 夏雪娟, 葉秀娟, 等. PCR-DGGE技術分析腌制麻竹筍中微生物多樣性[J]. 食品科學, 2014, 35(21): 170-174. DOI:10.7506/ spkx1002-6630-201421033.
[5] HOLZAPFEL W. Use of starter cultures in fermentation on a household scale[J]. Food Control, 1997, 8(5): 241-258. DOI:10.1016/ S0956-7135(97)00017-0.
[6] GRATTEPANCHE F, LACROIX C, AUDET P, et al. Quantification by real-time PCR of Lactococcus lactis subsp cremoris in milk fermented by a mixed culture[J]. Applied Microbiology and Biotechnology, 2005, 66(4): 414-421. DOI:10.1007/s00253-004-1822-0.
[7] 付琳琳, 曹郁生, 李海星, 等. 應用PCR-DGGE技術分析泡菜中乳酸菌的多樣性[J]. 食品與發酵工業, 2005, 31(12): 103-105. DOI:10.3321/j.issn:0253-990X.2005.12.027.
[8] 尹軍霞, 沈國娟, 謝亞芳. 分離自酸菜汁的乳酸乳球菌體外去除膽固醇特性[J]. 食品與生物技術學報, 2009, 28(6): 850-853. DOI:10.3321/j.issn:1673-1689.2009.06.025.
[9] 連元元, 王穎, 李晨, 等. 鎘脅迫下泡菜乳酸乳球菌的形態變化[J]. 食品科學, 2015, 36(1): 124-127. DOI:10.7506/spkx1002-6630-201501024.
[10] SEEMA B, ARASHDEEP S, ABHIJIT G. Probiotic characterization of potential hydrolases producing Lactococcus lactis subsp. lactis isolated from pickled yam[J]. International Journal of Food Sciences and Nutrition, 2014, 65(1): 53-61. DOI:10.3109/09637486.2013.832175.
[11] ASO Y, TAKEDA A, SATO M, et al. Characterization of lactic acid bacteria coexisting with a nisin Z producer in Tsuda-turnip pickles[J]. Current Microbiology, 2008, 57(1): 89-94. DOI:10.1007/s00284-008-9161-5.
[12] XU W, HUANG Z Y, ZHANG X J, et al. Monitoring the microbial community during solid-state acetic acid fermentation of Zhenjiang aromatic vinegar[J]. Food Microbiology, 2011, 28(6): 1175-1181. DOI:10.1016/j.fm.2011.03.011.
[13] ZHANG W J, CAI Q, GUAN X, et al. Detection of peanut (Arachis hypogaea) allergen by real-time PCR method with internal amplification control[J]. Food Chemistry, 2015, 174: 547-552. DOI:10.1016/j.foodchem.2014.11.091.
[14] ZHAO Y W, WU Z F, SHEN X Q, et al. Bacteria community analysis by quantitative real-time PCR of fermenting wax gourd and its changes of organic acids[J]. Journal of Food Processing and Preservation, 2014, 38(4): 1653-1659. DOI:10.1111/jfpp.12127.
[15] ROSANNA T, MARIA S, GIORGIA P, et al. Development and application of a real-time PCR-based assay to enumerate total yeasts and Pichia anomala, Pichia guillermondii and Pichia kluyveri in fermented table olives[J]. Food Control, 2012, 23(2): 356-362. DOI:10.1016/j.foodcont.2011.07.032.
[16] ACHILLEOS C, BERTHIER F. Quantitative PCR for the specific quantification of Lactococcus lactis and Lactobacillus paracasei and its interest for Lactococcus lactis in cheese samples[J]. Food Microbiology, 2013, 36(2): 286-295. DOI:10.1016/j.fm.2013.06.024.
[17] 李平蘭, 賀稚非. 食品微生物學實驗原理與技術[M]. 北京: 中國農業出版社, 2005: 264.
[18] 薩姆布魯克 J, 拉塞爾 D W. 分子克隆實驗指南[M]. 2版. 黃培堂,譯. 北京: 科學出版社, 2002: 1564-1594.
[19] 夏雪娟, 陳芝蘭, 陳宗道, 等. 16S rDNA序列分析法快速鑒定西藏地區傳統乳制品中的乳酸菌[J]. 食品科學, 2013, 34(14): 245-249. DOI:10.7506/spkx1002-6630-201314050.
[20] XU W T, LI L Y, LU J, et al. Analysis of caecal microbiota in rats fed with genetically modified rice by real-time quantitative PCR[J]. Journal of Food Science, 2011, 76(1): M88-M93. DOI:10.1111/1750-3841.12604.
[21] LIU J X, FENG Y M, CHEN Q S. The distribution of somatostatin precursor 1 and preproenkephalin mRNA in chicken intestinal nerve of Remak[J]. Acta Zoologica Sinica, 2007, 53(2): 294-302. DOI:10.3969/ j.issn.1674-5507.2007.02.014.
[22] 趙雪濤. 熒光定量PCR測定食品中臘樣芽孢桿菌[D]. 上海: 復旦大學, 2006.
[23] KAMILA O S, OLGA K R, ADLEY C C. Development of realtime pcr assays for detection and quantification of Bacillus cereus group species: differentiation of B. weihenstephanensis and rhizoid B. pseudomycoides isolates from milk[J]. Applied and Environmental Microbiology, 2011, 77(1): 80-88. DOI:10.1128/AEM.01581-10.
[24] 騫宇. 抗消化淀粉對實驗鼠腸道生理環境和胃腸功能性作用的影響[D]. 重慶: 西南大學, 2013.
[25] 張家超, 王芳, 徐海燕, 等. 6 種區分乳酸乳球菌乳酸亞種和乳酸乳球菌乳脂亞種的分子生物學方法比較[J]. 微生物學報, 2010, 50(12): 1670-1676. DOI:10.13343/j.cnki.wsxb.2010.12.007.
[26] HARRI L J, FLEMING H P, KLAENHAMMER T R. Characterization of 2 nisin-producing Lactococcus-lactis subsp. lactis strains isolated from a commercial sauerkraut fermentation[J]. Applied and Environmental Microbiology, 1992, 58(5): 1477-1483.
[27] XIONG T, LI X, GUAN Q Q, et al. Starter culture fermentation of Chinese sauerkraut: growth, acidification and metabolic analyses[J]. Food Control, 2014, 41: 122-127. DOI:10.1016/ j.foodcont.2013.12.033.
[28] XIONG T, GUAN Q Q, SONG S H, et al. Dynamic changes of lactic acid bacteria flora during Chinese sauerkraut fermentation[J]. Food Control, 2012, 26(1): 178-181. DOI:10.1016/j.foodcont.2012.01.027.
Detection of Lactococcus lactis in Pickled Ma Bamboo Shoots by Quantitative Real-time PCR
XIA Xuejuan1, ZHENG Jiong1,2,*, YE Xiujuan1, WU Jinsong1, KAN Jianquan1,2
(1. College of Food Science, Southwest University, Chongqing 400715, China; 2. Chongqing Engineering Research Center of Regional Food, Chongqing 400715, China)
The dynamic changes of Lactococcus lactis in pickled Ma bamboo shoots with 6 g/100 mL salt concentration were studied by quanti tative real-time polymerase chain reaction (qRT-PCR). After the DNA extraction from standard Lactococcus lactis, preparation of positive plasmids, protraction of standard curve, DNA extraction from samples at different fermentation stages and qRT-PCR, and quantitative detection were performed. The results showed that during fermentation (0–63 days), the concentration of Lactococcus lactis increased gradually from 2.41 × 102copies/?L (day 0) to the maximum value of 4.63 × 108copies/?L (day 14), an increase by six orders of magnitude, followed by a slow decrease to 5.02 × 106copies/?L at day 63. In conclusion, qRT-PCR technology provides a reliable, fast and effective way for the quantitative analysis of bacteria in pickled Ma bamboo shoots.
Ma bamboo shoots (Dendrocalamus latiflorus); pickling; quantitative real-time polymerase chain reaction (qRT-PCR); Lactococcus lactis; dynamic changes
10.7506/spkx1002-6630-201604016
TS201.3
A
1002-6630(2016)04-0088-05
夏雪娟, 鄭炯, 葉秀娟, 等. 實時熒光定量PCR技術監測腌制麻竹筍中乳酸乳球菌動態變化[J]. 食品科學, 2016, 37(4): 88-92. DOI:10.7506/spkx1002-6630-201604016. http://www.spkx.net.cn
XIA Xuejuan, ZHENG Jiong, YE Xiujuan, et al. Detection of Lactococcus lactis in pickled ma bamboo shoots by quantitative real-time PCR[J]. Food Science, 2016, 37(4): 88-92. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201604016. http://www.spkx.net.cn
2015-03-29
中央高校基本科研業務費專項(XDJK2013C131);重慶市特色食品工程技術研究中心能力提升項目(cstc2014pt-gc8001)
夏雪娟(1988—),女,博士研究生,研究方向為食品化學與營養學。E-mail:xiaxuej1989@163.com
*通信作者:鄭炯(1982—),男,講師,博士,研究方向為果蔬加工與質量控制。E-mail:zhengjiong_swu@126.com
猜你喜歡
城市道橋與防洪(2022年4期)2022-07-01 06:04:12
當代陜西(2019年8期)2019-05-09 02:22:48
上海建材(2019年1期)2019-04-25 06:30:48
動漫星空(興趣百科)(2019年3期)2019-03-07 07:23:10
家庭影院技術(2018年4期)2018-05-09 07:07:52
專用汽車(2016年4期)2016-03-01 04:13:43
質量與標準化(2015年9期)2015-12-31 11:41:40
中國質量與標準導報(2014年4期)2014-03-11 19:54:25
中國質量與標準導報(2014年10期)2014-02-28 22:25:47
中國質量與標準導報(2014年7期)2014-02-28 22:24:39
