基于線(xiàn)粒體Cyt b基因的中國(guó)南方唇魚(yú)骨(Hermibarbuslabeo)和間魚(yú)骨(Hermibarbusmedius)種群分化及親緣地理研究
2016-11-28 09:17:33藍(lán)昭軍范明君黃小林
生態(tài)學(xué)報(bào) 2016年19期
藍(lán)昭軍,范明君,黃小林,趙 俊
1 廣東省水產(chǎn)健康安全養(yǎng)殖重點(diǎn)實(shí)驗(yàn)室,廣東省高等學(xué)校生態(tài)與環(huán)境科學(xué)重點(diǎn)實(shí)驗(yàn)室,廣州市亞熱帶生物多樣性與環(huán)境生物監(jiān)測(cè)重點(diǎn)實(shí)驗(yàn)室,華南師范大學(xué)生命科學(xué)學(xué)院,廣州 510631 2 韶關(guān)市水產(chǎn)研究所, 韶關(guān) 512006 3 深圳華大水產(chǎn)科技有限公司, 深圳 518083 4 中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所, 廣州 510300
?
基于線(xiàn)粒體Cytb基因的中國(guó)南方唇魚(yú)骨(Hermibarbuslabeo)和間魚(yú)骨(Hermibarbusmedius)種群分化及親緣地理研究
藍(lán)昭軍1,2,范明君1,3,黃小林1,4,趙 俊1,*
1 廣東省水產(chǎn)健康安全養(yǎng)殖重點(diǎn)實(shí)驗(yàn)室,廣東省高等學(xué)校生態(tài)與環(huán)境科學(xué)重點(diǎn)實(shí)驗(yàn)室,廣州市亞熱帶生物多樣性與環(huán)境生物監(jiān)測(cè)重點(diǎn)實(shí)驗(yàn)室,華南師范大學(xué)生命科學(xué)學(xué)院,廣州 510631 2 韶關(guān)市水產(chǎn)研究所, 韶關(guān) 512006 3 深圳華大水產(chǎn)科技有限公司, 深圳 518083 4 中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所, 廣州 510300
為了解中國(guó)南方唇魚(yú)骨(Hermibarbuslabeo)和間魚(yú)骨(Hermibarbusmedius)的種群分化、親緣地理格局及物種有效性,作者對(duì)唇魚(yú)骨 8個(gè)水系及間魚(yú)骨 9個(gè)水系共148尾樣本的Cytb基因全序列進(jìn)行了測(cè)定。在所有序列中,共有128個(gè)變異位點(diǎn),共檢測(cè)出了41個(gè)單倍型,單倍型多樣性為0.954,核苷酸多樣性為0.02153。基于Cytb基因全序列構(gòu)建的NJ樹(shù)顯示,中國(guó)南方的唇魚(yú)骨與間魚(yú)骨合聚為兩大支,其中韓江和九龍江的全部唇魚(yú)骨樣本組成了Ⅰ支,而其他水系的唇魚(yú)骨和間魚(yú)骨樣本則組成了Ⅱ支。兩支系間的遺傳距離為5.1%,而唇魚(yú)骨和間魚(yú)骨之間的遺傳距離為3.2%。現(xiàn)有證據(jù)不支持間魚(yú)骨與唇魚(yú)骨達(dá)到種一級(jí)的分化。單倍型網(wǎng)絡(luò)圖顯示,韓江、九龍江種群和其他水系的種群分化較大;海南島三大水系種群和漠陽(yáng)江種群的單倍型分支與代表珠江水系單倍型的分支之間的親緣關(guān)系較近,與其他地理區(qū)間種群的親緣關(guān)系則相對(duì)較遠(yuǎn);西江可能為珠江水系、漠陽(yáng)江水系和海南島三大水系間魚(yú)骨種群的擴(kuò)散中心,其中一支往東向北江和東江擴(kuò)散;另一支往南向海南島擴(kuò)散,海南島三大水系種群爾后在冰期海退時(shí)向北擴(kuò)散至漠陽(yáng)江水系及珠江水系。AMOVA分析表明,唇魚(yú)骨和間魚(yú)骨地理區(qū)之間變異約占54.50%,地理區(qū)內(nèi)種群間變異約占18.64%,種群內(nèi)的變異占26.86%,這說(shuō)明,唇魚(yú)骨和間魚(yú)骨種群Cytb的遺傳分化主要是來(lái)自地理區(qū)之間。錯(cuò)配分析及中性檢驗(yàn)顯示,全部種群、唇魚(yú)骨種群、間魚(yú)骨種群在歷史上均沒(méi)有發(fā)生過(guò)明顯的擴(kuò)張,兩個(gè)mtDNA支系亦未發(fā)生過(guò)擴(kuò)張,而海南島的昌化江種群曾發(fā)生過(guò)種群擴(kuò)張。
唇魚(yú)骨;間魚(yú)骨;種群分化;親緣地理;細(xì)胞色素b基因;中國(guó)南方
親緣地理學(xué),又稱(chēng)譜系生物地理學(xué),是研究有密切親緣關(guān)系的種間和種內(nèi)支系現(xiàn)有的地理分布格局的形成過(guò)程和形成機(jī)制的學(xué)科。生物在漫長(zhǎng)的演化過(guò)程中,受各種因素(地質(zhì)因素、氣候因素等)的影響,都會(huì)在其演化的過(guò)程中留下印跡,如影響生物的地理隔離、遷移、區(qū)域擴(kuò)張和滅絕等,而生物的這些活動(dòng)都會(huì)具有影響一定區(qū)域內(nèi)物種基因流的作用,改變遺傳變異的分布,從而產(chǎn)生新的物種進(jìn)化模式[1]。尤其是,地質(zhì)地貌的格局對(duì)于種群基因流的改變和分化往往起著重要的作用[2- 6]。而分子親緣地理學(xué)是采用分子生物技術(shù),在分子水平上探討物種的系統(tǒng)地理格局的形成機(jī)制,進(jìn)一步闡述其起源進(jìn)化歷史,分析區(qū)域類(lèi)群在時(shí)間上和空間上的發(fā)展變化,從而重建生物區(qū)系的進(jìn)化歷史過(guò)程[7]。
唇魚(yú)骨(Hemibarbuslabeo)和間魚(yú)骨(Hemibarbusmedius)均隸屬于鯉形目(Cypriniformes),鯉科(Cyprinidae),鮈亞科(Gobioninae),魚(yú)骨屬(Hemibarbus)[8- 9],兩者為近緣種。間魚(yú)骨是在原唇魚(yú)骨種群中劃分出的一個(gè)種[8],在此之前,一直被歸為唇魚(yú)骨[10- 14]。1995年,樂(lè)佩琦對(duì)我國(guó)魚(yú)骨屬魚(yú)類(lèi)進(jìn)行分類(lèi)整理后認(rèn)為,唇魚(yú)骨華南地區(qū)及西南部分地區(qū)的種群與唇魚(yú)骨其他地方種群的差別已經(jīng)達(dá)到了物種級(jí)的水平,并以海南島的12尾標(biāo)本作為模式標(biāo)本,將華南及西南部分地區(qū)的唇魚(yú)骨種群描述為一新種——間魚(yú)骨(H.mediusYue),并指出其與唇魚(yú)骨的鑒別特征主要為吻長(zhǎng)較短,唇稍薄,不甚發(fā)達(dá),下唇側(cè)葉稍狹窄,無(wú)皺褶,背鰭刺較細(xì)弱及鰓耙數(shù)較少等[8]。1998年,樂(lè)佩琦等較為詳細(xì)地記載唇魚(yú)骨和間魚(yú)骨的地理分布,其中唇魚(yú)骨分布于我國(guó)臺(tái)灣各水系和閩江、錢(qián)塘江、長(zhǎng)江、黃河等直至黑龍江水系;間魚(yú)骨分布于華南、西南部分地區(qū)各水系[9]。
然而,目前不同學(xué)者對(duì)于兩者的分布區(qū)域與樂(lè)佩琦等的觀(guān)點(diǎn)不盡相同[15- 18]。唇魚(yú)骨和間魚(yú)骨之間的鑒別特征較為模糊,且某些性狀可能存在連續(xù)變化,不易區(qū)分。因此,造成對(duì)兩者分布區(qū)描述差異的原因可能為物種鑒定存在偏差。此外,兩者的分布均較為廣泛,是研究親緣地理的理想素材。有關(guān)唇魚(yú)骨和間魚(yú)骨的親緣地理的研究,目前已有林宗鍵[17]及Lin等[18]基于線(xiàn)粒體控制區(qū)序列對(duì)唇魚(yú)骨的報(bào)道,但該研究并未涉及珠江等水系的種群。因此,作者在中國(guó)南方唇魚(yú)骨和間魚(yú)骨的分布區(qū),采集不同地理種群的樣本,對(duì)其線(xiàn)粒體細(xì)胞色素b(Cytb)基因序列進(jìn)行了測(cè)定與分析,旨在澄清二者物種有效性的前提下,探討其種群分化和分布特征,推測(cè)其種群演化歷史及親緣地理模式。
1 材料與方法
1.1 樣品采集
本研究所用樣品均為2009年9月—2010年8月采自貴州省、廣西壯族自治區(qū)、廣東省、海南省、福建省、江西省和浙江省等地,所有樣本根據(jù)樂(lè)佩琦等提出的分類(lèi)鑒定特征[8- 9]進(jìn)行鑒定,其中間魚(yú)骨樣本共92尾;唇魚(yú)骨樣本56尾。將所有個(gè)體按所采樣的水系進(jìn)行種群劃分,即用水系名稱(chēng)對(duì)種群進(jìn)行命名。具體的樣品信息及采樣點(diǎn)見(jiàn)表1。
1.2 DNA的提取及測(cè)序
基因組 DNA的提取參考慶寧等[19]的方法,即采用從上海生工生物工程技術(shù)服務(wù)有限公司購(gòu)買(mǎi)的“基因組 DNA抽提試劑盒”所示方法,略有改動(dòng)。PCR反應(yīng)體系:反應(yīng)體系約25μL,按順序分別將PCR Master Mix 12.5μL、Primer1 3μL、Primer2 3μL、模板DNA 4μL及滅菌雙蒸水2.5μL混合,每一次反應(yīng)都用ddH2O代替模板DNA做陰性對(duì)照。反應(yīng)在PTC100或PTC200型PCR儀上進(jìn)行,具體擴(kuò)增程序如下:A: 95℃預(yù)變性4min,B: 94℃變性40sec,C: 58℃退火45sec,D: 72℃延伸1min,E: 重復(fù)步驟B—D45次;F:最后72℃延伸10min。PCR及測(cè)序所用引物均為通用引物L(fēng)14724和H15915,由上海英駿生物技術(shù)有限公司合成,其序列分別為:L14724:5′-GACTTG AAA AACCAC CGTTG- 3′;H15915:5′-CTCCGATCTCCGGATTACAAGAC- 3′。PCR產(chǎn)物用1%瓊脂糖電泳檢驗(yàn)?zāi)康钠魏?將PCR產(chǎn)物送上海生工生物工程技術(shù)服務(wù)有限公司純化并測(cè)序。
1.3 數(shù)據(jù)分析
對(duì)測(cè)得的序列使用Clustal X 1.81 軟件進(jìn)行排序比對(duì)并輔以人工校對(duì)。用軟件MEGA 5.0[20]計(jì)算所分析序列的堿基組成。采用Kimura′s two-parameter模型計(jì)算遺傳距離,并采用該軟件中的鄰接法(Neighbor-joining method,NJ),以似鮈(Pseudogobiovaillanti)作為外類(lèi)群,構(gòu)建分支系統(tǒng)樹(shù)。用軟件DNAsp 5.0[21]對(duì)各種群序列的多樣性進(jìn)行分析,統(tǒng)計(jì)核苷酸多樣性(Nucleotide diversity,π)與單倍型多樣性(Haplotype diversity,h),并以此估計(jì)不同水系種群的遺傳多樣性;并利用該軟件進(jìn)行錯(cuò)配分布(Mismatch-distribution)分析及Tajima′s D和Fu and Li′s檢驗(yàn),以檢驗(yàn)種群是否發(fā)生過(guò)擴(kuò)張。使用Network 4.6 軟件,以Median-joining法構(gòu)建各單倍型之間網(wǎng)絡(luò)關(guān)系圖,并對(duì)圖中各個(gè)單倍型進(jìn)行群體對(duì)應(yīng)關(guān)系分析。利用Arlequin Ver 2000[22]軟件進(jìn)行分子變異分析(AMOVA),測(cè)試種群間與地理區(qū)間遺傳變異程度。
2 結(jié)果與分析
2.1 Cyt b 基因的序列變異
本研究共得到148尾唇魚(yú)骨與間魚(yú)骨的Cytb基因全序列,序列長(zhǎng)1140bp,無(wú)堿基的插入或缺失,Cytb基因所有序列中,共128個(gè)變異位點(diǎn),占全序列的11.23%,其中14個(gè)為單個(gè)多態(tài)位點(diǎn),114個(gè)為簡(jiǎn)約信息位點(diǎn)。大多數(shù)的變異發(fā)生在密碼子第3位(82.81%),第1位(14.02%)和第2位(3.12%)的變異較低。唇魚(yú)骨與間魚(yú)骨在Cytb序列中A、C、T、G平均含量分別為27.5%、28.4%、28.4%和15.6%,G的含量明顯低于其他3種堿基含量,表現(xiàn)出很強(qiáng)的堿基組成偏向性。其中A+T的含量55.9%,G+C的含量為44.0%,A+T的含量明顯高于G+C的含量。
2.2 Cyt b 基因的單倍型多樣性和遺傳多樣性
在唇魚(yú)骨56尾樣本中,檢測(cè)出19個(gè)單倍型,其中17個(gè)為特有單倍型,2個(gè)為與間魚(yú)骨共享的單倍型;間魚(yú)骨92尾樣本,檢測(cè)出24個(gè)單倍型,其中22個(gè)為獨(dú)有單倍型,2個(gè)為與唇魚(yú)骨共享的單倍型(表1)。在全部148尾樣本中,共檢測(cè)出了41個(gè)單倍型,單倍型多樣性為0.954。
在一些不同的種群中,存在共享單倍型,表明這些種群的親緣關(guān)系十分密切。其中海南島三大水系之一南渡江(NAN)、廣東西部獨(dú)立入海水系漠陽(yáng)江(MOY)及珠江水系第二大支流北江(BEI)的種群共享單倍型NAN1-MOY1-BEI5;西江支流桂江(GUI)和柳江(LIU)的間魚(yú)骨種群與長(zhǎng)江水系支流湘江(XIA)的唇魚(yú)骨種群共享單倍型GUI1-LIU1-XIA1。
單倍型多樣性(h)和核苷酸多樣性(π)分析表明(表1),148尾唇魚(yú)骨和間魚(yú)骨樣本的單倍型多樣性為0.954,核苷酸多樣性為0.02153,遺傳多樣性較高。核苷酸多樣性(π)分析表明(表1),湘江、桂江、北江、東江、九龍江、閩江種群的核苷酸遺傳多樣性較高,均在0.002以上。
2.3 種群間遺傳距離及遺傳分化
統(tǒng)計(jì)表明,唇魚(yú)骨和間魚(yú)骨之間的遺傳距離為3.2%,兩者遺傳差異不大。表2分別顯示了不同水系的唇魚(yú)骨與間魚(yú)骨基于Cytb基因的遺傳距離和及遺傳分化指數(shù)。由表2可知,在各地理區(qū)(如海南島地區(qū)、東南地區(qū)等)和水系內(nèi)部(如長(zhǎng)江水系和珠江水系)的遺傳距離、遺傳分化指數(shù)較低;珠江水系和長(zhǎng)江水系的遺傳距離、遺傳分化指數(shù)亦較低。在唇魚(yú)骨和間魚(yú)骨各個(gè)種群之間,韓江、九龍江和閩江種群與其他各種群的遺傳距離較大,遺傳分化指數(shù)較高,而余下各種群之間的遺傳距離和遺傳分化指數(shù)則較低。
2.4 系統(tǒng)發(fā)育分析
為更好地了解中國(guó)南方地區(qū)唇魚(yú)骨與間魚(yú)骨各種群之間的親緣關(guān)系,以似鮈為外類(lèi)群,對(duì)148尾樣品的Cytb基因中的41個(gè)單倍型進(jìn)行系統(tǒng)發(fā)育分析,利用最大鄰接法(NJ)構(gòu)建分子系統(tǒng)發(fā)育樹(shù)(圖1)。
在我國(guó)南方的唇魚(yú)骨與間魚(yú)骨種群間,聚成Ⅰ和Ⅱ兩支。韓江和九龍江的全部樣本組成了Ⅰ支,而其他水系的樣本則組成了Ⅱ支,支系Ⅰ和Ⅱ之間的遺傳距離為5.1%,這說(shuō)明韓江和九龍江種群已和其他種群發(fā)生了一定程度的遺傳分化。在Ⅱ支中,又可分為A和B個(gè)兩姐妹群。A群包含了海南島三大水系、漠陽(yáng)江、柳江、賀江、北江、錢(qián)塘江和甌江的全部樣本及桂江、東江、湘江和閩江的部分樣本;B群則包含了沅江的全部樣本及桂江、湘江和閩江的部分樣本。在支系Ⅱ的A群中,又可分為A1和A2兩個(gè)姐妹群。A1有兩個(gè)分支,分別為海南島三大水系、漠陽(yáng)江的全部樣本和北江水系一尾樣本組成一個(gè)分支,說(shuō)明海南島三大水系種群與漠陽(yáng)江水系種群的親緣關(guān)系較近;而賀江、柳江的全部標(biāo)本及湘江、北江和東江的部分樣本組成另一支,這則說(shuō)明珠江水系三大支流之間的遺傳差異不大,而珠江水系與湘江水系之間的遺傳差異亦較低。A2則包含了錢(qián)塘江、甌江的全部樣本及閩江部分樣本和東江的一尾樣本。
就唇魚(yú)骨與間魚(yú)骨這兩個(gè)物種而言,除了支系Ⅰ全部為唇魚(yú)骨、A1其中的一個(gè)分支全部為間魚(yú)骨之外,其余均含有唇魚(yú)骨與間魚(yú)骨兩個(gè)物種,這也說(shuō)明兩者間遺傳差異不大。
2.5 單倍型網(wǎng)絡(luò)分析
圖2顯示了中國(guó)南方唇魚(yú)骨和間魚(yú)骨基于Cytb基因的單倍型網(wǎng)絡(luò)親緣關(guān)系,大多數(shù)單倍型之間的變異為1—7步,遺傳差異不大。但是桂江和湘江種群的共享單倍型GUI2-XIA2與韓江和九龍江種群之間的變異近40步;桂江與錢(qián)塘江、甌江與閩江種群的變異達(dá)約20步,表明這些種群的單倍型間存在一定的遺傳分化。

物種Species水系Riversystems采集地Samplelocations樣品數(shù)Samplesize單倍型(個(gè)體數(shù))Haplotypes(numberofindividuals)變異位點(diǎn)數(shù)Numberofpolymorphicsites(S)單倍型數(shù)Numebrofhaplotypes單倍型多樣性Haplotypediversity(h)核苷酸多樣性Nucleotidediversity(π)間魚(yú)骨H.medius昌化江(CHJ)五指山10CHJ1(3)、CHJ2(2)、CHJ3(1)、CHJ4(1)、CHJ5(2)450.8670.00121南渡江(NAN)屯昌9NAN1-MOY1-BEI5*(9)010.0000.00000萬(wàn)泉河(WAN)瓊海9WAN1(7)、WAN2(2)120.3890.00034漠陽(yáng)江(MOY)陽(yáng)春11NAN1-MOY1-BEI5(10)、MOY2(1)120.1820.00016東江(DON)河源10DON1(1)、DON2(1)、DON3(5)、DON4-HEJ1(3)*、2940.7110.00532北江(BEI)連州9BEI1(1)、BEI2(10)、BEI3(1)、BEI4(1)、1750.5060.00224始興5NAN1-MOY1-BEI5(1)賀江(HEJ)賀州11DON4-HEJ1(4)、HEJ2(7)120.5090.00045桂江(GUI)桂林6GUI1-LIU1-XIA1(4)*、GUI2-XIA2(1)*、GUI3(2)、3050.8060.00614荔浦3GUI4-LIU2(1)*、GUI5(1)柳江(LIU)從江3GUI1-LIU1-XIA1(3)、GUI4-LIU2(2)、LIU3(4)430.6670.00137永福6唇魚(yú)骨H.labeo韓江(HAN)上杭10HAN1(8)、HAN2(1)、HAN3(1)330.3780.00053九龍江(JIU)漳平5JIU1(2)、JIU2(3)120.6000.03072閩江(MIN)邵武9MIN1(2)、MIN2(1)、MIN3(1)、MIN4(2)、MIN5(3)4250.8610.01652湘江(XIA)全州9GUI1-LIU1-XIA1(6)、GUI2-XIA2(3)2720.5000.01184沅江(YUA)錦屏7YUA1(7)010.0000.00000信江(XIN)上饒3XIN1(2)、XIN2(1)120.6670.00058甌江(OUJ)麗水9OUJI(4)、OUJ2(5)220.5000.00088錢(qián)塘江(QIA)開(kāi)化4QIA1(3)、QIA2(1)120.5000.00044合計(jì)Total148128410.9540.02153
﹡表示此單倍型為不同河流共享單倍型
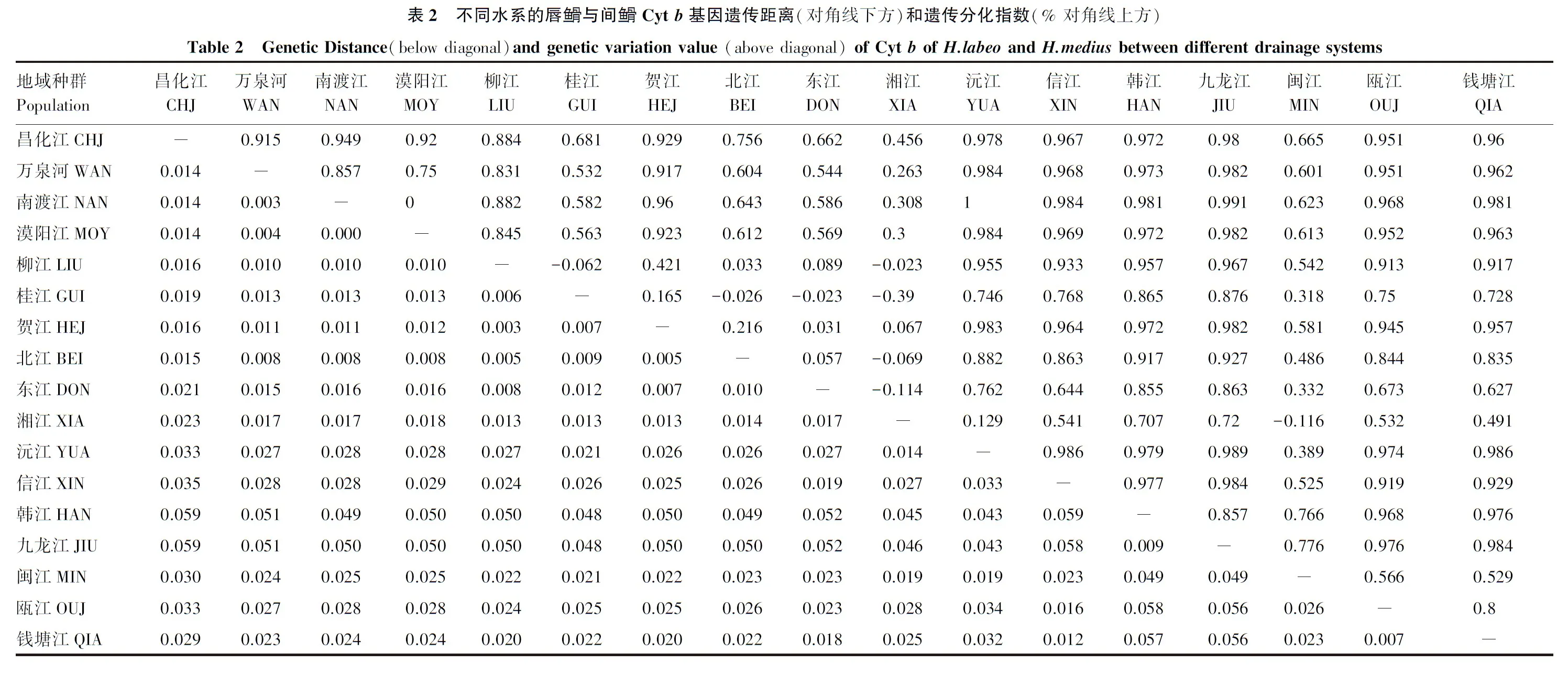
表2 不同水系的唇魚(yú)骨與間魚(yú)骨Cyt b基因遺傳距離(對(duì)角線(xiàn)下方)和遺傳分化指數(shù)(% 對(duì)角線(xiàn)上方)
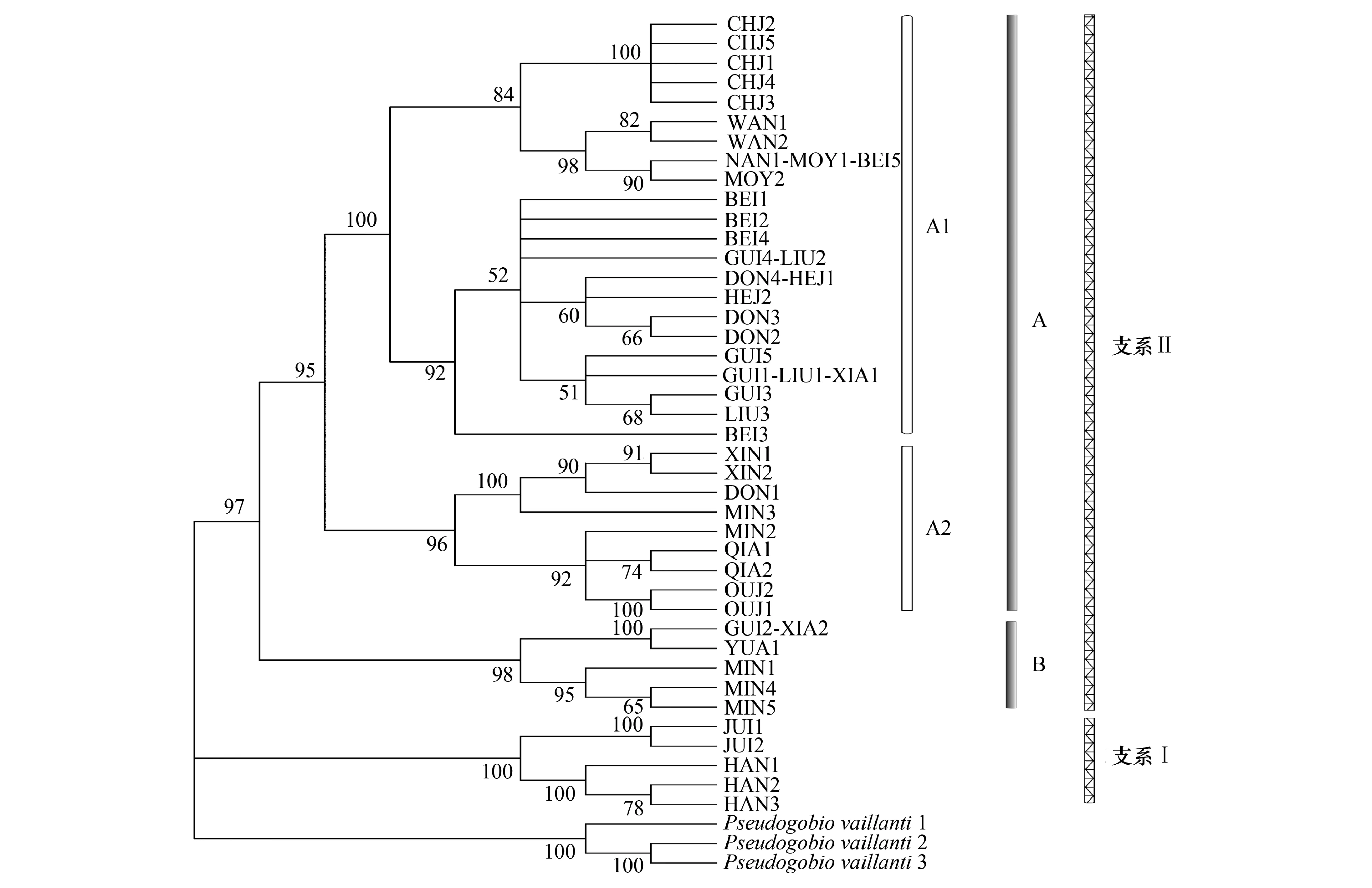
圖1 唇魚(yú)骨與間魚(yú)骨中國(guó)南方種群基于Cyt b基因的最大鄰接NJ樹(shù)Fig.1 Neighbor-joining tree of H.labeo and H.medius populations of south China based on Cyt b gene

圖2 唇魚(yú)骨與間魚(yú)骨中國(guó)南方種群基于Cyt b基因的單倍型網(wǎng)絡(luò)圖Fig.2 Minimum spanning network of Cyt b sequences of H.labeo and H.medius populations of south China
單倍型網(wǎng)絡(luò)圖還揭示,桂江及柳江水系種群與長(zhǎng)江水系支流湘江水系種群的親緣關(guān)系十分密切,兩者具有共享單倍型且各個(gè)單倍型之間的變異不大;海南島三大水系的種群與漠陽(yáng)江種群的親緣關(guān)系比較密切;錢(qián)塘江、甌江及閩江種群的親緣關(guān)系亦較為密切;韓江與九龍江種群的親緣關(guān)系較近;而我國(guó)東南部水系閩江的種群不僅與長(zhǎng)江水系支流湘江、沅江的關(guān)系較近,且與珠江水系支流東江、長(zhǎng)江水系支流信江種群的關(guān)系亦較密切。這與NJ樹(shù)的結(jié)果相似。從單倍型網(wǎng)絡(luò)圖推測(cè),西江可能為珠江水系、漠陽(yáng)江水系和海南島水系間魚(yú)骨種群的擴(kuò)散中心,其中一支往東向北江和東江擴(kuò)散;另一支往南向海南島擴(kuò)散,海南島種群爾后在第四紀(jì)冰期海退時(shí)向北擴(kuò)散至漠陽(yáng)江水系及珠江水系。
2.6 分子變異分析(AMOVA)
為了解唇魚(yú)骨和間魚(yú)骨種群分子變異的分布模式,根據(jù)系統(tǒng)發(fā)育分析及單倍型網(wǎng)絡(luò)分析的結(jié)果,將唇魚(yú)骨和間魚(yú)骨17個(gè)水系的種群分成5個(gè)地理區(qū),進(jìn)行分子變異分析(AMOVA)。地理區(qū)的劃分主要根據(jù)各種群親緣關(guān)系的親疏及地理位置,其中海南島的南渡江、萬(wàn)泉河、昌化江和漠陽(yáng)江為一地理區(qū);珠江水系為一地理區(qū),包括西江支流柳江、桂江、賀江、珠江另外兩條支流北江及東江;長(zhǎng)江水系為一地理區(qū),包括湘江、沅江及信江;韓江和九龍江為一地理區(qū);閩江、甌江和錢(qián)塘江為一個(gè)地理區(qū)。AMOVA分析分析表明,唇魚(yú)骨和間魚(yú)骨地理區(qū)之間變異約占54.50%,地理區(qū)內(nèi)種群間約占18.64%,種群內(nèi)的變異占26.86%(表3)。這說(shuō)明,唇魚(yú)骨和間魚(yú)骨種群Cytb的遺傳分化主要是來(lái)自地理區(qū)之間。
2.7 種群動(dòng)態(tài)分析
除了昌化江、桂江、北江、東江、閩江等種群外,其他種群的單倍型數(shù)目未達(dá)到4,故無(wú)法計(jì)算Fu and Li′s D和Tajima′s D值。對(duì)唇魚(yú)骨和間魚(yú)骨的全部種群、唇魚(yú)骨種群和間魚(yú)骨種群進(jìn)行錯(cuò)配分析發(fā)現(xiàn),全部種群、唇魚(yú)骨及間魚(yú)骨種群均呈現(xiàn)多峰分布,說(shuō)明其均沒(méi)有發(fā)生過(guò)明顯的擴(kuò)張。而中性檢驗(yàn)也顯示,全部種群、唇魚(yú)骨及間魚(yú)骨種群的Fu and Li′s D和Tajima′s D值均沒(méi)有呈現(xiàn)顯著的負(fù)值,表明其在歷史上均沒(méi)有發(fā)生過(guò)明顯的擴(kuò)張(表4)。表4還表明,兩個(gè)支系的Fu and Li′s D和Tajima′s D值均沒(méi)有呈現(xiàn)顯著的負(fù)值,這說(shuō)明兩個(gè)支系亦未發(fā)生過(guò)擴(kuò)張。

表3 唇魚(yú)骨和間魚(yú)骨分子變異分析

表4 唇魚(yú)骨與間魚(yú)骨種群基于Cyt b基因的Tajima′s D 和 Fu and Li′s D檢驗(yàn)
就各個(gè)水系的種群而言,桂江、北江、東江、閩江等種群的Fu and Li′s D和Tajima′s D均不為顯著的負(fù)值,說(shuō)明這幾個(gè)種群亦相對(duì)穩(wěn)定,沒(méi)有發(fā)生過(guò)擴(kuò)張;而昌化江(CHJ)的Fu and Li′s D為顯著的負(fù)值,而Tajima′s D則不顯著,但其錯(cuò)配分析圖為單峰分布(圖3),所以可以推測(cè)昌化江種群曾發(fā)生過(guò)種群擴(kuò)張。

圖3 昌化江種群的歧點(diǎn)分布分析圖(其中實(shí)線(xiàn)代表期望值,虛線(xiàn)代表觀(guān)察值)Fig.3 Mismatch-distribution analysis of H.medius of Changhuajiang population of mtDNA haplotype sequences(solid lines represent the distribution expeted under constant population size, and dotted lines represent the observe distribution)
3 討論
3.1 間魚(yú)骨物種有效性
在本研究中,遺傳距離分析表明,唇魚(yú)骨和間魚(yú)骨之間的遺傳距離為3.2%,低于唇魚(yú)骨種內(nèi)各種群間的最大遺傳距離(韓江與信江,5.9%)。系統(tǒng)分析顯示,間魚(yú)骨并未形成單系群,而是與部分唇魚(yú)骨種群組成一支,而韓江和九龍江的唇魚(yú)骨組成了另一支。單倍型分布分析顯示,湘江唇魚(yú)骨種群的所有單倍型均與桂江或柳江的間魚(yú)骨種群共享,湘江、桂江和柳江種群的遺傳差異極小。系統(tǒng)發(fā)育分析及單倍型網(wǎng)絡(luò)圖還顯示,除了唇魚(yú)骨的湘江、沅江種群與間魚(yú)骨的桂江、柳江種群的親緣關(guān)系較近外,唇魚(yú)骨的信江和閩江種群與間魚(yú)骨的東江種群亦有著較近的親緣關(guān)系。此外,藍(lán)昭軍基于多變量形態(tài)度量學(xué)的研究亦表明,唇魚(yú)骨和間魚(yú)骨存在一定的形態(tài)差異,但各個(gè)種群間存在梯度變異,差異并不顯著,無(wú)法截然分開(kāi)[23];而林龍峰基于線(xiàn)粒體CO I和ND5基因分別對(duì)中國(guó)南方唇魚(yú)骨與間魚(yú)骨的種群遺傳分化進(jìn)行了研究,結(jié)果均表明間魚(yú)骨與唇魚(yú)骨之間的遺傳差異較小,兩者存在共享單倍型,且間魚(yú)骨不能單獨(dú)構(gòu)成單系群,而是與唇魚(yú)骨部分樣本構(gòu)成一個(gè)分支[24]。因此,綜合考慮基于線(xiàn)粒體Cytb、CO I和ND5基因和多變量形態(tài)度量學(xué)的分析結(jié)果,我們認(rèn)為唇魚(yú)骨和間魚(yú)骨之間的遺傳差異未達(dá)到種級(jí)水平,間魚(yú)骨應(yīng)為唇魚(yú)骨的次定同物異名。
3.2 韓江、九龍江種群與其他種群間的遺傳分化
系統(tǒng)發(fā)育分析顯示,在我國(guó)南方的唇魚(yú)骨與間魚(yú)骨種群間,可以分成Ⅰ和Ⅱ兩支。韓江和九龍江種群遺傳差異不大,兩條江的全部樣本組成了Ⅰ支,而其他種群的樣本則組成了Ⅱ支,兩支系之間的遺傳距離為5.1%,這說(shuō)明韓江和九龍江種群已和其他種群發(fā)生了一定程度的遺傳分化。林弘都[25]和梁曉旭等[26]對(duì)部分淡水魚(yú)類(lèi)的研究亦表明,華南大陸東部的蓮花山脈是粵東地區(qū)的一個(gè)重要地理屏障,而慶寧等[27]對(duì)華南地區(qū)黑眶蟾蜍(BufoMelanostictus)的研究亦得出了相似的結(jié)果。
韓江是廣東省東部地區(qū)的最大河流,其有東西兩源,東源稱(chēng)梅江,發(fā)源于蓮花山山脈北麓;西源稱(chēng)汀江,發(fā)源于武夷山南麓。蓮花山脈位于廣東省東北部,其山勢(shì)由粵北部的大埔向西南一直延伸到香港附近,最高峰為銅鼓障,高1559 m; 整個(gè)山脈高度基本在980—1 170 m之間[28]。九龍江為福建省第二大水系,發(fā)源于福建玳瑁山南麓。由此可知,韓江和九龍江被蓮花山、武夷山、玳瑁山等較高的山脈圍繞,將其與珠江水系、長(zhǎng)江水系及閩江水系隔離。據(jù)此,我們推測(cè):一方面,由于蓮花山、武夷山、玳瑁山等山脈的隔離,韓江和九龍江的種群無(wú)法與其他種群進(jìn)行基因交流,因而產(chǎn)生了相對(duì)較大的遺傳分化;另一方面,韓江和九龍江在冰期海退時(shí),可能曾發(fā)生過(guò)連接,使得兩種群之間得以基因交流,因而遺傳差異不顯著。
3.3 珠江及長(zhǎng)江種群間的遺傳分化
分子系統(tǒng)發(fā)育分析顯示,除湘江與桂江和柳江種群的親緣關(guān)系較近以及東江少數(shù)樣本與信江、閩江種群的親緣關(guān)系較近外,唇魚(yú)骨和間魚(yú)骨剩下的絕大部分單倍型則分別歸入了不同的分支之內(nèi)。這表明,在唇魚(yú)骨和間魚(yú)骨分布區(qū)之間,可能存在地理障壁,造成其不同的分布范圍,而這一地理障壁可能為長(zhǎng)江水系和珠江水系的分水嶺之一——南嶺山脈。南嶺山脈由越城嶺、都龐嶺、萌渚嶺、騎田嶺和大庾嶺5條主要山嶺所組成。其橫亙?cè)谙婀稹⑾婊洝②M粵之間,向東延伸至閩南,東西長(zhǎng)約600km,南北寬約200km。廣義的南嶺還包括苗兒山、海洋山、九嶷山、香花嶺、瑤山、九連山等。有研究表明,南嶺山脈在11.06—8.04百萬(wàn)年前已經(jīng)形成[29],但在隆起初期,其高度相對(duì)較低,可能不足以阻隔兩側(cè)的淡水魚(yú)類(lèi)的基因交流。而在青藏高原隆起的時(shí)期,南嶺山脈也呈加速隆起之勢(shì)[25]。南嶺的加速隆起,可能使其成為了南嶺以北水系與南嶺以南水系間的地理障壁,因而限制了南北兩側(cè)水系種群的基因交流,使得其兩側(cè)的種群產(chǎn)生了遺傳分化,進(jìn)而形態(tài)上也發(fā)生了一定程度的分化。
在本研究中,多種分析結(jié)果亦顯示,長(zhǎng)江水系支流湘江、沅江及信江的唇魚(yú)骨種群與珠江水系間魚(yú)骨種群的親緣關(guān)系較近,其中湘江與沅江的種群在系統(tǒng)發(fā)育樹(shù)中與桂江和柳江的親緣關(guān)系均較近;而東江的部分樣本則與信江種群的親緣關(guān)系較近。在Li等對(duì)馬口魚(yú)(Opsariichthysbidens)的研究[30]及Yang等對(duì)大鰭鳠(Hemibagrusmacropterus)的研究[31]中,其結(jié)果均顯示珠江水系的種群與長(zhǎng)江水系的種群親緣關(guān)系較近。
我們推測(cè),造成這種情況的原因可能有二,一是由于地殼運(yùn)動(dòng)、洪水泛濫等原因,使長(zhǎng)江水系和珠江水系在歷史上曾發(fā)生過(guò)襲奪事件或連接事件,使長(zhǎng)江南部支流上游的部分支流被珠江襲奪或與珠江水系支流相連接,從而使兩水系種群得以基因交流;二是水利樞紐工程的影響,在湘江水系與桂江水系上游,有一條建成已有2200多年人工運(yùn)河——靈渠將兩江聯(lián)通[32],這也可能促進(jìn)長(zhǎng)江水系與珠江水系種群的基因交流。
3.4 海南島種群的起源、擴(kuò)散及隔離
從唇魚(yú)骨和間魚(yú)骨單倍型的分布可知,海南島的南渡江水系、廣東西部水系漠陽(yáng)江及北江水系共享一個(gè)單倍型,表明三種群間親緣關(guān)系較近。在單倍型網(wǎng)絡(luò)圖中,單倍型WAN1與桂江、柳江和湘江的共享單倍型GUI1-LIU1-XIA1及桂江的單倍型GUI3之間的變異均較小,約為10步,表明這些單倍型間有著較近的親緣關(guān)系;萬(wàn)泉河的單倍型WAN1與南渡江、漠陽(yáng)江和北江的共享單倍型NAN1-MOY1-BEI5只有3步,兩者之間亦有著很近的親緣關(guān)系。此外,單倍型網(wǎng)絡(luò)圖還顯示,海南島中部山脈以東的南渡江和萬(wàn)泉河種群與西邊的昌化江種群之間關(guān)系相對(duì)較遠(yuǎn)(16步)。
有關(guān)華南沿海地質(zhì)構(gòu)造的研究顯示,在第四紀(jì)時(shí),瓊州海峽至少經(jīng)歷過(guò)4次海侵與海退[33];而在第四紀(jì)冰期,由于海平面下降,華南沿海的海岸線(xiàn)在海南島以南,海南島北部均為遼闊的濱海平原[34];在更新世,在海南島和越南之間是一片陸地,其間有一條聯(lián)系西江、廣西、越南沿海和海南島諸水系的古河道[35]。有鑒于此,結(jié)合本研究系統(tǒng)發(fā)育分析及單倍型網(wǎng)絡(luò)圖等研究結(jié)果,海南島的間魚(yú)骨種群有可能為西江水系的種群沿著上述這一古河道擴(kuò)散而來(lái),然后進(jìn)一步在海南島內(nèi)擴(kuò)散。隨后,由于海平面的上升,海南島與大陸隔離,因而使海南島種群與大陸種群的基因交流受到了限制,發(fā)生了遺傳分化。而到了晚更新世冰期時(shí),由于海退等原因,雷州半島與海南島連成一片山地丘陵[36],這使得海南島的種群得以向北擴(kuò)散,到達(dá)雷州半島,進(jìn)而擴(kuò)散至漠陽(yáng)江水系,并重新回到珠江水系。間魚(yú)骨的這一擴(kuò)散途徑,與海南紋胸鮡(Glyptothoraxhainanensis)[37]及中間黃顙魚(yú)(Pelteobagrusintermedius)[38]有相似之處。
在林宗鍵和Lin等對(duì)唇魚(yú)骨的親緣地理研究中,基于線(xiàn)粒體控制區(qū)序列的分析結(jié)果顯示,海南島昌化江種群(實(shí)為間魚(yú)骨)與長(zhǎng)江中游種群的親緣關(guān)系較其與廣東東部水系榕江種群的關(guān)系近,而昌化江與長(zhǎng)江的距離遠(yuǎn)大于其與榕江的距離,他們認(rèn)為其原因是長(zhǎng)江和海南島的水系曾通過(guò)古紅河水系發(fā)生過(guò)連接[17- 18]。值得指出的是,在Lin等的研究中,并沒(méi)有采集珠江水系的樣本,而海南島也只有昌化江水系的樣本。在本研究中,也發(fā)現(xiàn)海南島三大水系種群與長(zhǎng)江水系種群的親緣關(guān)系近于其與韓江和九龍江水系種群的親緣關(guān)系。由于韓江與榕江的地理位置很近,故此該結(jié)果與Lin等的相似。可以認(rèn)為,造成本研究及Lin等所述之結(jié)果的另一重要原因是海南島的種群由西江水系種群沿著古河道擴(kuò)散而來(lái),而西江曾與長(zhǎng)江水系發(fā)生過(guò)襲奪或連接事件,其種群與長(zhǎng)江水系種群親緣關(guān)系較近。因此,這一現(xiàn)象進(jìn)一步印證了我們對(duì)海南島種群來(lái)源及擴(kuò)散之推測(cè)。
另一方面,在海南島內(nèi)部,單倍型網(wǎng)絡(luò)圖顯示昌化江種群與萬(wàn)泉河及南渡江種群的變異達(dá)到16步,說(shuō)明昌化江種群與其他兩種群之間,發(fā)生了一定的遺傳分化。地質(zhì)研究表明,在第三紀(jì)末期,海南島中部山地已經(jīng)形成,在多雨山區(qū)發(fā)源的河流也四散分流,獨(dú)自入海,不相聯(lián)系匯合[39]。在間魚(yú)骨擴(kuò)散至海南島各主要水系后,可能由于海南島中部山地,如五指山等的隆起,將海南島西部的昌化江種群與東部的萬(wàn)泉河及南渡江種群隔離,進(jìn)而使昌化江種群產(chǎn)生了分化。而左艷玲等基于線(xiàn)粒體DNA控制區(qū)序列對(duì)擬平鰍(Liniparhomalopteradisparis)的研究也表明:擬平鰍的昌化江種群與南渡江和萬(wàn)泉河的種群亦存在較大的遺傳分化[40]。因此,本研究的結(jié)果進(jìn)一步說(shuō)明了海南島中部山脈可能對(duì)海南島的淡水魚(yú)類(lèi)存在著較為廣泛的隔離。
致謝:華南師范大學(xué)陳湘粦教授和慶寧教授幫助實(shí)驗(yàn)方案設(shè)計(jì)及修改,林弘都博士和張雄同學(xué)參與了部分樣品的采集,Radhakrishnan博士幫助寫(xiě)作,特此致謝。
[1] Hewitt G. The genetic legacy of the quaternary ice ages. Nature, 2000, 405(6789): 907- 913.
[2] Angers B, Magnan P, Plante M, Bernatchez L. Canonical correspondence analysis for estimating spatial and environmental effects on microsatellite gene diversity in brook charr (Salvelinusfontinalis). Molecular Ecology, 1999, 8(6): 1043- 1053.
[3] Keyghobadi N, Roland J, Strobeck C. Influence of landscape on the population genetic structure of the alpine butterflyParnassiussmintheus(Papilionidae). Molecular Ecology, 1999, 8(9): 1481- 1495.
[4] Castric V, Bonney F, Bernatchez L. Landscape structure and hierarchical genetic diversity in the brook charr,Salvelinusfontinalis. Evolution, 2001, 55(5): 1016 - 1028.
[5] Cassel A, Tammaru T. Allozyme variability in central, peripheral and isolated populations of the scarce heath (Coenonymphahero: Lepidoptera, Nymphalidae); implications for conservation. Conservation Genetics, 2003, 4(1): 83- 93.
[6] Costello A B, Down T E, Pollard S M, Pacas C J, Taylor E B. The influence of history and contemporary stream hydrology on the evolution of genetic diversity within species: an examination of microsatellite DNA variation in bull trout,Salvelinusconfluentus(Pisces: Salmonidae). Evolution, 2003, 57(2): 328- 344.
[7] Hewitt G M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biological Journal of the Linnean Society, 1996, 58: 247- 276.
[8] 樂(lè)佩琦. 魚(yú)骨和間魚(yú)骨屬魚(yú)類(lèi)的分類(lèi)整理(鯉形目: 鯉科). 動(dòng)物分類(lèi)學(xué)報(bào), 1995, 20(1): 116- 123.
[9] 陳宜瑜. 中國(guó)動(dòng)物志硬骨魚(yú)綱鯉形目(中卷). 北京: 科學(xué)出版社, 1998: 239- 241.
[10] 伍獻(xiàn)文. 中國(guó)鯉科魚(yú)類(lèi)志(下卷). 上海: 上海人民出版社, 1977: 439- 537.
[11] 廣西壯族自治區(qū)水產(chǎn)研究所, 中國(guó)科學(xué)院動(dòng)物研究所. 廣西淡水魚(yú)類(lèi)志. 南寧: 廣西人民出版社, 1981: 112- 113.
[12] 中國(guó)水產(chǎn)科學(xué)研究院珠江水產(chǎn)研究所等. 海南島淡水及河口魚(yú)類(lèi)志. 廣州: 廣東科學(xué)技術(shù)出版社, 1986: 93- 94.
[13] 鄭慈英. 珠江魚(yú)類(lèi)志. 北京: 科學(xué)出版社, 1989: 119- 121.
[14] 中國(guó)水產(chǎn)科學(xué)研究院珠江水產(chǎn)研究所等. 廣東淡水魚(yú)類(lèi)志. 廣州: 廣東科技出版社, 1991.
[15] Yang J Q, He S P, Freyhof J, Witte K, Liu H Z. The Phylogenetic Relationships of the Gobioninae (Teleostei: Cyprinidae) Inferred from Mitochondrial CytochromebGene Sequences. Hydrobiologia, 2006, 553(1): 255- 266.
[16] 廣西壯族自治區(qū)水產(chǎn)研究所, 中國(guó)科學(xué)院動(dòng)物研究所. 廣西淡水魚(yú)類(lèi)志(第二版). 南寧: 廣西人民出版社, 2006, 193- 195.
[17] 林宗鍵. 唇魚(yú)骨和間魚(yú)骨的親緣地理研究. 臺(tái)灣:國(guó)立成功大學(xué), 2009.
[18] Lin C J, Lin H D, Wang J P, Chao S C, Chiang T Y. Phylogeography ofHemibarbuslabeo(Cyprinidae): secondary contact of ancient lineages of mtDNA. Zoologica Scripta, 2010, 39(1): 23- 35.
[19] 慶寧, 丘城鋒, 廖偉群, 馬天峰, 梁曉旭, 列金妮. 華南沿海西部美麗小條鰍基于線(xiàn)粒體控制區(qū)的種群遺傳變異及親緣地理格局. 生態(tài)學(xué)報(bào), 2010, 30(1): 258- 264.
[20] Kumar S, Nei M, Dudley J, Tamura K. MEGA: a biologist-centric software for evolutionary analysis of DNA and protein sequences. Briefings in Bioinformatics, 2008, 9(4): 299- 306.
[21] Librado P, Rozas J. DNASP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 2009, 25(11): 1451- 1452.
[22] Excoffier L, Smouse P E, Quattor J M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics, 1992, 131(2): 479- 491.
[23] 藍(lán)昭軍. 中國(guó)南方唇魚(yú)骨和間魚(yú)骨的種群分化、親緣地理及間魚(yú)骨的生物學(xué)特征[D]. 廣州: 華南師范大學(xué), 2011.
[24] 林龍峰. 基于COI和ND5序列的中國(guó)南方唇魚(yú)骨和間魚(yú)骨種群遺傳變異與親緣地理分析[D]. 廣州: 華南師范大學(xué), 2014.
[25] 林弘都. 臺(tái)灣與中國(guó)大陸地區(qū)鯉科魚(yú)類(lèi)之親緣地理研究[D]. 臺(tái)灣: 國(guó)立成功大學(xué), 2008.
[26] 梁曉旭, 慶寧, 楊柯林, 萬(wàn)彩霞, 趙俊, 陳湘粦. 廣東地區(qū)寬鰭鱲種群遺傳變異和親緣地理. 水生生物學(xué)報(bào), 2010, 34(4): 806- 814.
[27] 慶寧, 馬天峰, 梁曉旭, 林弘都, 盧文華, 鹽司櫓. 華南地區(qū)黒眶蟾蜍的遺傳變異和地理分化. 動(dòng)物分類(lèi)學(xué)報(bào), 2011, 36(2): 356- 367.
[28] 曾昭璇, 黃偉峰. 廣東自然地理. 廣州: 廣東人民出版社, 2001.
[29] 地質(zhì)礦產(chǎn)部《南嶺項(xiàng)目》構(gòu)造專(zhuān)題組. 南嶺區(qū)域構(gòu)造特征及控巖控礦構(gòu)造研究. 北京: 地質(zhì)出版社, 1988.
[30] Li G Y, Wang X Z, Zhao Y H, Zhang J, Zhang C G, He S P. Speciation and phylogeography ofOpsariichthysbidens(Pisces: Cypriniformes: Cyprinidae) in China: analysis of the cytochrome b gene of mtDNA from diverse populations. Zoological Studies, 2009, 48(4): 569- 583.
[31] Yang L, Mayden R L, He S P. Population genetic structure and geographical differentiation of the Chinese catfishHemibagrusmacropterus(Siluriformes, Bagridae): evidence for altered drainage patterns. Molecular Phylogenetics and Evolution, 2009, 51(2): 405- 411.
[32] 宗鳳書(shū). 靈渠地質(zhì)地貌初步研究. 中國(guó)巖溶, 1990, 9(1): 88- 93.
[33] 趙煥庭, 王麗榮, 袁家義. 瓊州海峽成因與時(shí)代. 海洋地質(zhì)與第四紀(jì)地質(zhì), 2007, 27(2): 33- 40.
[34] 張虎男, 陳偉光. 華南沿海新構(gòu)造運(yùn)動(dòng)與地質(zhì)環(huán)境. 北京: 地震出版社, 1990.
[35] Harold K V. Maps of Pleistocene sea levels in Southeast Asia: shorelines, river systems and time durations. Journal of Biogeography, 2000, 27(5): 1153- 1167.
[36] 中國(guó)科學(xué)院南海海洋研究所海洋地質(zhì)研究室. 華南沿海第四紀(jì)地質(zhì). 北京: 科學(xué)出版社, 1978.
[37] Chen X L, Chiang T Y, Lin H D, Zheng H S, Shao K T, Zhang Q, Hsu K C. Mitochondrial DNA phylogeography ofGlyptothoraxfokiensisandGlyptothoraxhainanensisin Asia. Journal of Fish Biology, 2007, 70(S): 75- 93.
[38] 慶寧, 呂鳳義, 廖偉群, 趙俊, 丘城鋒. 中間黃顙魚(yú)群體遺傳變異與親緣生物地理. 動(dòng)物學(xué)報(bào), 2007, 53(5): 845- 852.
[39] 曾昭璇, 曾憲中. 海南島自然地理. 北京: 科學(xué)出版社, 1989.
[40] 左艷玲, 林岳光, 梁曉旭, 馬天峰, 慶寧. 基于mtDNA控制區(qū)序列的擬平鰍遺傳變異和種群分化. 水產(chǎn)學(xué)報(bào), 2009, 33(6): 925- 931.
Population diversity and phylogeography ofHemibarbuslabeoandHemibarbusmediusin South China
LAN Zhaojun1,2, FAN Mingjun1,3, HUANG Xiaolin1,4, ZHAO Jun1,*
1GuangdongProvincialKeyLaboratoryforHealthyandSafeAquaculture,KeyLaboratoryofEcologyandEnvironmentScienceinGuangdongHigherEducation,GuangzhouKeyLaboratoryofSubtropicalBiodiversityandBiomonitor,SchoolofLifeScience,SouthChinaNormalUniversity,Guangzhou510631,China2ShaoguanFisheriesResearchInstitute,Shaoguan512006,China3ShenzhenBGIFisheriesScience&Technologycorporation,Ltd,Shenzhen518083,China4SouthChinaSeaFisheriesResearchInstitute,ChineseAcademyofFisherySciences,Guangzhou510300,China
HemibarbuslabeoandHemibarbusmedius(Cypriniformes: Cyprinidae: Gobioninae:Hemibarbus) are primary freshwater fish species, and their wide distribution makes them ideal models for phylogeographical studies. In this study, variations in the population genetics and phylogeographical patterns of 148 specimens belonging to 8 populations ofH.labeoand 9 populations ofH.mediuscollected from 17 drainage systems in south China were investigated using the nucleotide sequences of the mtDNA Cytochromebgene (1140 bp). The results reveal 128 variable sites (11.23%) within the gene sequences ofH.labeoandH.medius. A total of 41 haplotypes were identified, with the haplotype diversity (h) and nucleotide diversity (π) being 0.954 and 0.02153, respectively. This indicates a high level of genetic diversity and evolutionary potential in both species. The results of the neighbor-joining tree demonstrate thatH.labeoandH.mediusindividuals fall into two major clades (clade I and clade II): clade I is composed of all specimens collected from the Hanjiang and Jiulongjiang Rivers, all of which were identified asH.labeo. Meanwhile, all remaining populations fell into clade II, which includes specimens of bothH.labeoandH.medius. The genetic distance between clade I and clade II was 5.1%, while that betweenH.labeoandH.mediuswas 3.2%. Our results indicate that the specimens collected from these sampling localities may not be differentiated into two or more subspecies. The haplotype network indicates that the populations of the Hanjiang and Jiulongjiang Rivers exhibit a relatively high level of genetic variation compared to that of the rest of the rivers in the region, and that the populations from Hainan Island and Moyangjiang River were genetically close to those from the Pearl River System, but not to those from the Yangtze River System,Hanjiang River, Jiulongjiang River, Minjiang River, Oujiang River and Qiantangjiang River. TheH.mediuspopulations of south China seem to have originated from the Xijiang River and reached south China by one of two routes: one route may be through the Xijiang River to the Beijiang and Dongjiang rivers; and the other may be though the southern Guangxi drainages to Hainan Island, and then to the Moyangjiang and Beijiang rivers through the Qiongzhou Strait. According to the neighbor-joining tree and the haplotype network, five regions of genetic distribution were defined: Hainan Island and the Moyangjiang River region; the Pearl River region; the Yangtze River region; the Hanjiang and Jiulongjiang river region; and the Minjiang, Oujiang, and Qiantangjiang river region. An analysis of molecular variance (AMOVA) showed that the genetic variation of populations among regions was 54.50%; conversely, the genetic variation among populations within regions was 18.64%, and that within the populations of different regions was 26.86%. This indicates that the greatest genetic variation is found among the populations of the different regions. Mismatch distribution and tests of neutrality taking populations ofH.labeoandH.mediusinto account both together and separately were all multimodal types, and the value of Fu, Li′s D, and Tajima′s D for all comparisons were all negative but non-significant or positive. The results reveal relatively stable populations ofH.labeoandH.medius. Mismatch distribution and tests of neutrality also showed that both clade I and clade II were relatively stable; however, the population of the Changhuajiang River has undergone an obvious population expansion.
Hemibarbuslabeo;Hemibarbusmedius; population diversity; phylogeography;Cytochromebgene; South China
國(guó)家自然科學(xué)基金項(xiàng)目(31372178);公益性行業(yè)(農(nóng)業(yè))科研專(zhuān)項(xiàng)經(jīng)費(fèi)資助(201303048);廣東省科技計(jì)劃項(xiàng)目(2015A070706009)
2014- 11- 04;
日期:2016- 01- 15
10.5846/stxb201411042169
*通訊作者Corresponding author.E-mail: zhaojun@scnu.edu.cn
藍(lán)昭軍,范明君,黃小林,趙俊.基于線(xiàn)粒體Cytb基因的中國(guó)南方唇魚(yú)骨(Hermibarbuslabeo)和間魚(yú)骨(Hermibarbusmedius)種群分化及親緣地理研究.生態(tài)學(xué)報(bào),2016,36(19):6091- 6102.
Lan Z J, Fan M J, Huang X L, Zhao J.Population diversity and phylogeography ofHemibarbuslabeoandHemibarbusmediusin South China.Acta Ecologica Sinica,2016,36(19):6091- 6102.

- 生態(tài)學(xué)報(bào)的其它文章
- 2010年中國(guó)農(nóng)作物凈初級(jí)生產(chǎn)力及其空間分布格局
- 立地類(lèi)型對(duì)張廣才嶺天然白樺林生態(tài)系統(tǒng)碳儲(chǔ)量的影響
- 大興安嶺北部興安落葉松(Larixgmelinii)林下穿透雨空間分布特征
- 模擬酸雨對(duì)紅樹(shù)林底泥中營(yíng)養(yǎng)元素及Cu、Zn分布的影響
- 不同林齡油松(Pinustabulaeformis)人工林植物、凋落物與土壤C、N、P化學(xué)計(jì)量特征
- 酸棗葉片結(jié)構(gòu)可塑性對(duì)自然梯度干旱生境的適應(yīng)特征