13個鐵線蓮品種的核型比較分析
2016-12-09 02:15:34任佳倫季夢成劉志高
浙江農林大學學報 2016年6期
任佳倫,季夢成,趙 爽,劉志高
(浙江農林大學 風景園林與建筑學院,浙江 臨安 311300)
13個鐵線蓮品種的核型比較分析
任佳倫,季夢成,趙 爽,劉志高
(浙江農林大學 風景園林與建筑學院,浙江 臨安 311300)
鐵線蓮屬Clematis為觀賞價值極高的藤本植物。鐵線蓮品種數量繁多,形態變異豐富,遺傳背景也相對復雜,品種間存在雜交不親和性的障礙,對雜交親本的染色體核型分析等細胞學特征的研究顯得十分重要。采用酶解法-去壁低滲火焰干燥法與Giemsa-C分帶染色技術,以13個鐵線蓮品種自然生長的幼嫩根尖為試材,進行染色體參數和核型分析。結果表明:13個品種的染色體均為2n=2x=16,均為二倍體;染色體核型分為2種類型:2n= 2x=16=10m+4st+2t,2n=2x=16=10m+6st;隨體均在第6條,第7條或第8條染色體上,且位置均在短臂上;13個品種的核型類型均為 “2A”,具有很大的相似性,說明雜交不親和現象由其他因素導致。圖3表3參18
植物細胞遺傳學;鐵線蓮屬;染色體;核型
1 材料與方法
1.1 試驗材料
13個鐵線蓮品種均取自浙江農林大學鐵線蓮種質資源圃(表1),所取試材均為自然生長狀態下幼嫩根尖。
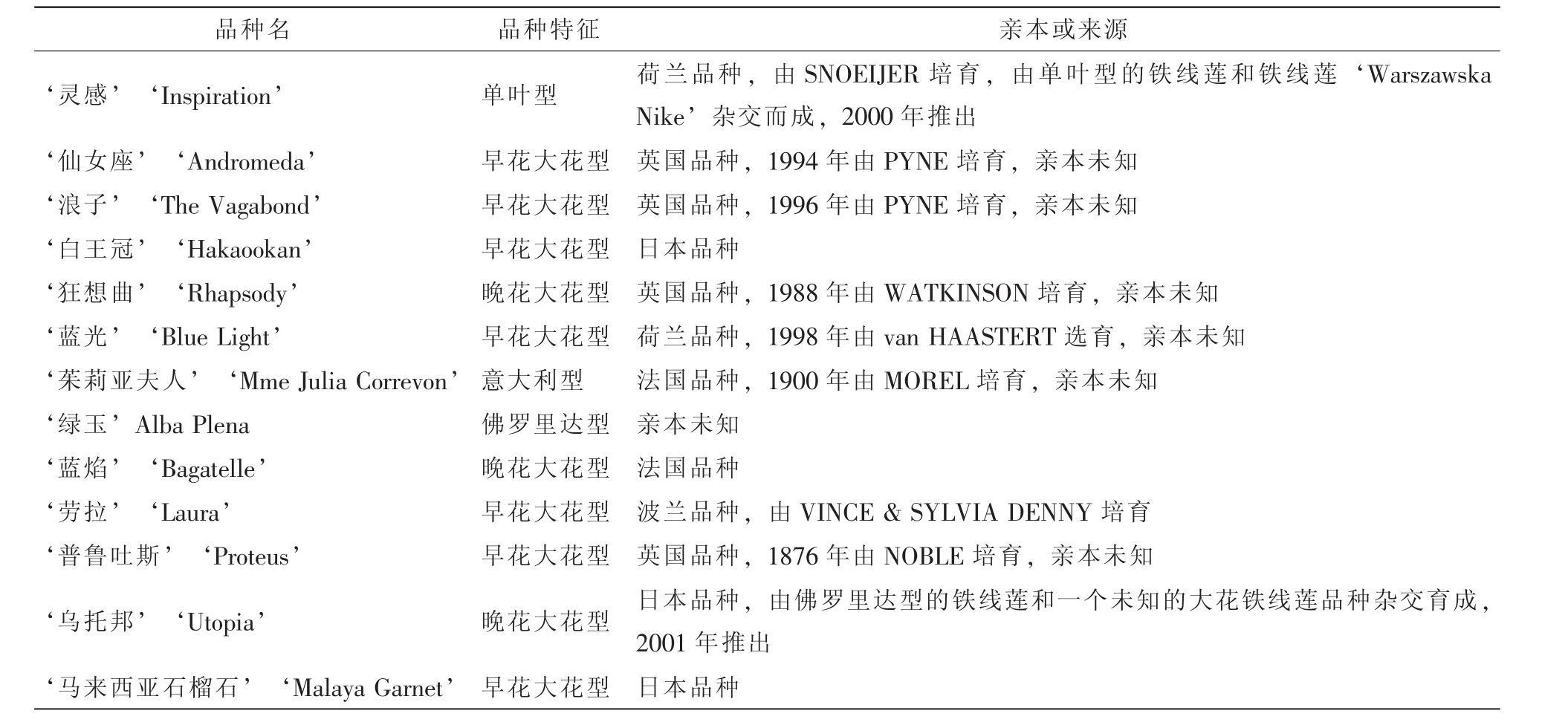
表1 13個鐵線蓮品種的基本信息Table 1 Information of thirteen Clematis cultivars
1.2 試驗方法
1.2.1 染色體制片 采用酶解法與去壁低滲火焰干燥法制片[8]。將材料放入0.002 mol·L-18-羥基喹啉溶液中預處理4.0~4.5 h,預處理后以0.075 mol·L-1氯化鉀溶液進行前低滲處理,0.5 h后在V(甲酸)∶V(冰乙酸)=3∶1的根尖固定液下固定數小時。再以體積分數為2.5%纖維素酶和2.5%果膠酶混合液對根尖進行4.0~5.0 h的酶解去壁,酶解完全后去除上清液和雜質,再固定0.5 h后用滴管將此黏稠溶液滴于預先在乙醇溶液中冷凍-20℃的載玻片上,用乙醇燈火焰微熱烘干,即得到含有根尖細胞的玻片。
1.2.2 染色 采用染色體Giemsa-C分帶技術[9]進行染色。將0.2 mol·L-1氯化氫和2×檸檬酸鈉(SSC)緩沖液分別放入60℃恒溫水浴鍋中加熱,將制好的載玻片放入0.2 mol·L-1氯化氫中處理160 s。之后將玻片放入水中洗去殘留的氯化氫,再放入氫氧化鋇溶液中處理420~430 s,后轉入水龍頭下將氫氧化鋇溶液沖洗掉。最后將玻片放入2×SSC水浴鍋中加熱,1.0 h后放入20.0 mL磷酸氫二鈉+10.0 mL磷酸二氫鉀+50~60滴Giemsa染液的混合溶液中染色數小時。
1.2.3 鏡檢 染色后用蒸餾水輕輕沖洗玻片,空氣中干燥后即可進行染色體觀測和拍照。先在40倍物鏡下找到分裂較好的染色體,向載玻片滴2滴二甲苯,蓋玻片滴2滴鏡油,再轉至100倍物鏡下拍照。
1.2.4 參數計算及制作核型模式圖 統計細胞分裂相30個·品種-1,其中85%以上的分裂相中染色體一致的數目定為該品種的染色體數[10]。分析時,各個品種取1個染色體形態好且分散好的分裂相,染色體相對長度的計算按照LEVAN等[11]的方法。核型類型以染色體長度比和臂比值來衡量,方法參照STEBINNS[12]的分類標準。著絲點位置的命名按照LEVAN等[11]的方法。核不對稱系數(As.K.,%)的計算按照ARANO[13]的方法。染色體相對長度系數(I.R.L)的分類參照KUO等[14]的方法。以Adobe Photoshop CS測量染色體的長短臂與隨體的相對長度[15],再以Excel計算染色體的相對長度[16],臂比值,臂比均值(AAR),臂比值大于2的染色體比例(PCA),臂指數(N.F值),著絲點指數等,并制作核型模式圖。
2 結果與分析
經觀察發現:13個鐵線蓮品種均為二倍體,染色體數為2n=2x=16,染色體基數為8,且品種間的核型公式極為相似,核型類型均為2A。13個鐵線蓮品種的核型參數見表2。由表2可知:核型公式‘靈感’為2n=2x=16=10m+4st+2t(2SAT), ‘仙女座’為2n=2x=16=10m+6st(4SAT), ‘浪子’為2n=2x=16=10m+4st(2SAT)+2t, ‘白王冠’為2n=2x=16=10m+4st+2t(2SAT), ‘狂想曲’為2n=2x=16=10m+6st(2SAT), ‘藍光’為2n=2x=16=10m+4st+2t(2SAT), ‘茱莉亞夫人’為2n=2x=16=10m+6st(2SAT),‘綠玉’為2n=2x=16=10m+4st+2t(2SAT), ‘藍焰’為2n=2x=16=10m+6st(2SAT), ‘勞拉’為2n=2x=16=10m+6st(2SAT), ‘普魯吐斯’為2n=2x=16=10m+4st+2t(2SAT), ‘烏托邦’為2n=2x=16=10m+6st(2SAT), ‘馬來西亞石榴石’為2n=2x=10m+2st+4t(4SAT)。
13個品種的臂指數均為26,臂比值大于2的染色體比例均為37.50%。由圖1~圖3可知, ‘靈感’‘白王冠’ ‘狂想曲’ ‘綠玉’ ‘藍焰’在第8對染色體上具有隨體, ‘仙女座’ ‘馬來西亞石榴石’于第6對與第8對染色體上具有隨體, ‘浪子’ ‘茱莉亞夫人’ ‘普魯吐斯’ ‘烏托邦’于第7對染色
體上具有隨體, ‘藍光’ ‘勞拉’在第6對染色體上具有隨體。其他染色體組的核型參數比較見表3。
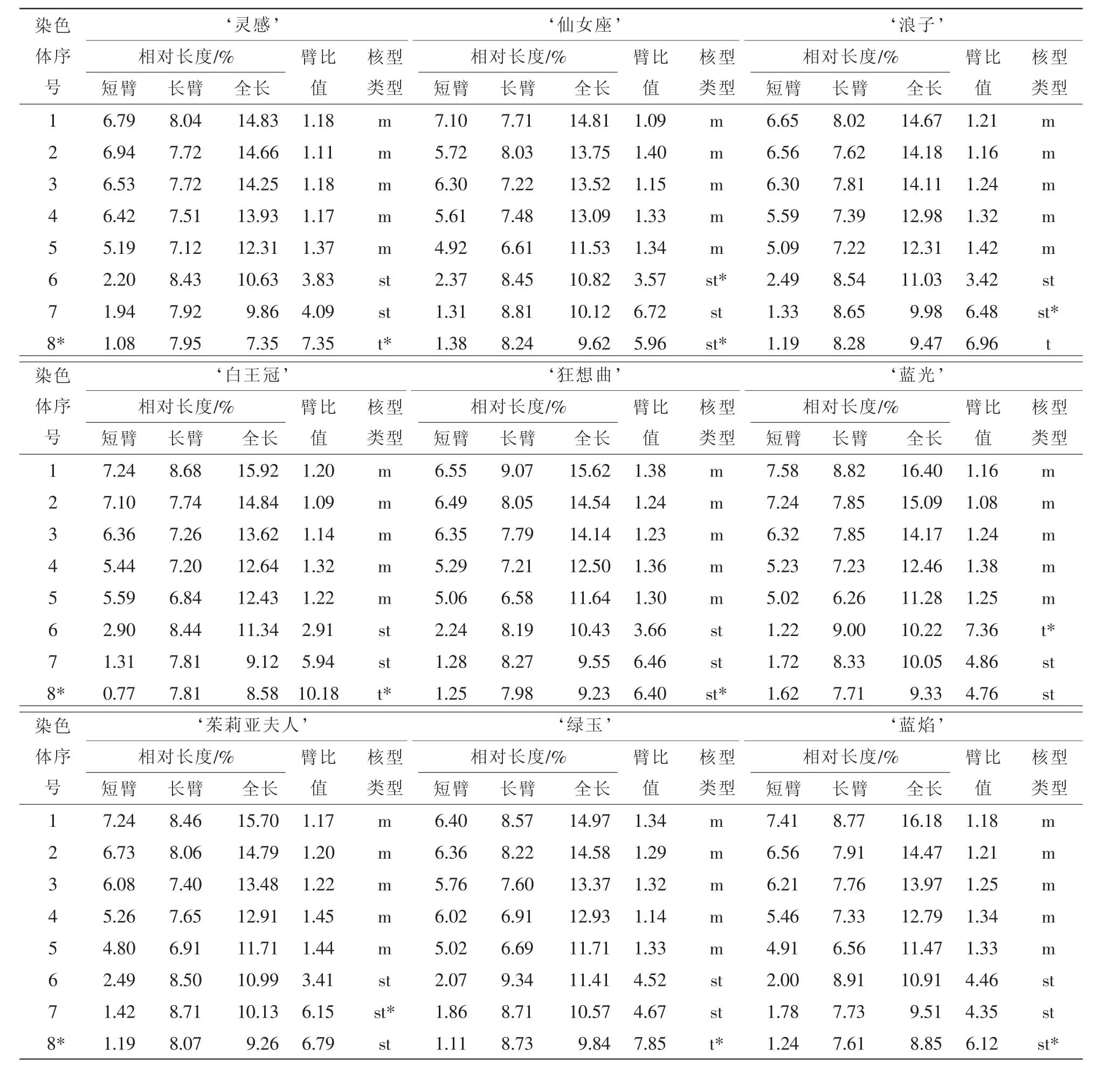
表2 13個鐵線蓮品種的染色體參數Table 2 Chromosomes parameters of thirteen Clematis cultivars
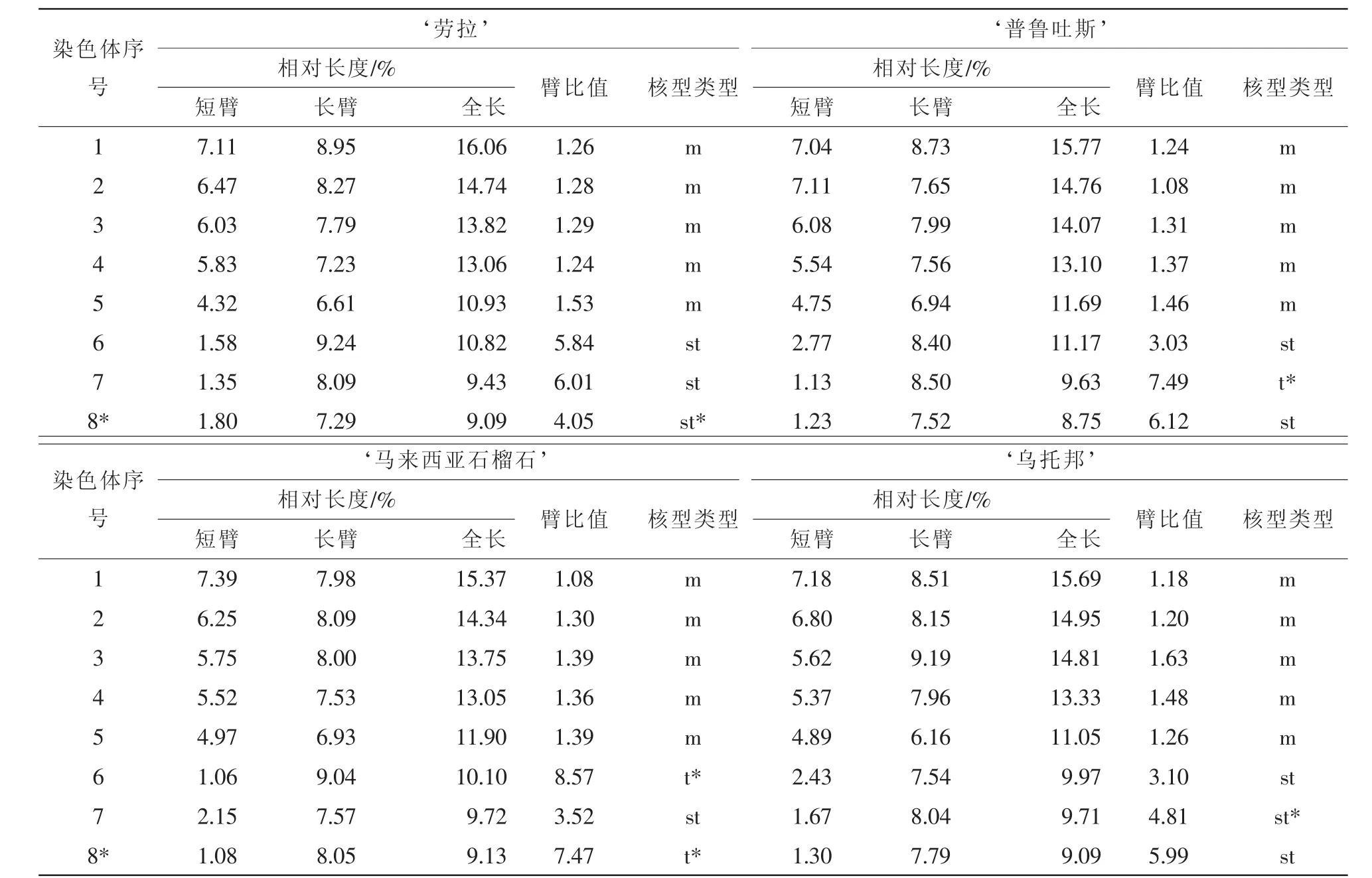
表2 (續)Table 2 Continued
3 討論
核型分析是細胞分類學的一個重要指標,能夠識別染色體的形態特征,對研究系統的演化、種間親緣關系及分類有重要的意義[17]。本研究報道了13個鐵線蓮栽培品種的染色體數目與核型,均為首次詳細報道。13個品種均為二倍體(2n=2x=16),染色體基數為8,說明鐵線蓮品種的染色體在數目上是比較恒定的,這與彭綠春等[7]的研究結果一致,這證明鐵線蓮屬的染色體基數比較恒定的表現為x=8。本試驗中13個鐵線蓮品種的核型類型均為 “2A”,這與張鐿鋰等[2,4]的研究結果有所不同。
13個品種的臂指數值均為26,核型結構為:第1~5對均為具中部著絲點(m)染色體,第6對為均為具近端(st)著絲點染色體,第7至8對為具近端(st)著絲點染色體或端部(t)著絲點染色體,這說明鐵線蓮品種的核型基本模式極為一致,這與張鐿鋰等[4]研究一致。但這13個品種所包含的m,st,t這4類染色體的數量卻不盡相同,從核型模式圖中可明顯看出這種差異。核型分析結果表明,13個鐵線蓮品種都能觀察到隨體染色體,除 ‘仙女座’和 ‘馬來西亞石榴石’具有2對隨體外,其他品種均具有1對隨體,且位置都在短臂上,這與前人的研究結果也一致[2-4]。在被子植物中,核型進化的基本趨勢是由對稱向不對稱方向發展的,系統演化上處于比較古老或原始的植物往往具有較對稱的核型,不對稱的核型通常出現在較進化或特化的植物中[12]。本研究中13個鐵線蓮品種的核不對稱系數均相對較低(61.78%~64.78%),變異很小,其按進化程度由高到低的順序為 ‘綠玉’ ‘茱莉亞夫人’ ‘浪子’ ‘勞拉’ ‘烏托邦’ ‘普魯吐斯’ ‘馬來西亞石榴石’ ‘狂想曲’ ‘藍光’ ‘藍焰’ ‘仙女座’ ‘靈感’‘白王冠’。此結果與楊光穗等人的研究結果基本一致[18]。
本研究13個鐵線蓮品種的核型特征極為相似,說明雜交不親和可能是溫度、濕度或人為等其他因素造成,有待于進一步探究。13個品種間的核型參數間存在的差異,可為鐵線蓮雜交育種提供細胞學資料。本研究中各染色體顯帶都不明顯,因此,未將染色體Giemsa-C分帶技術應用到鐵線蓮品種的核
型分析中。

圖1 13個鐵線蓮品種的染色體中期分裂相(×1 000)Figure 1 Metaphase cleavage phase of chromosomes of thirteen Clematis cultivars(×1 000)
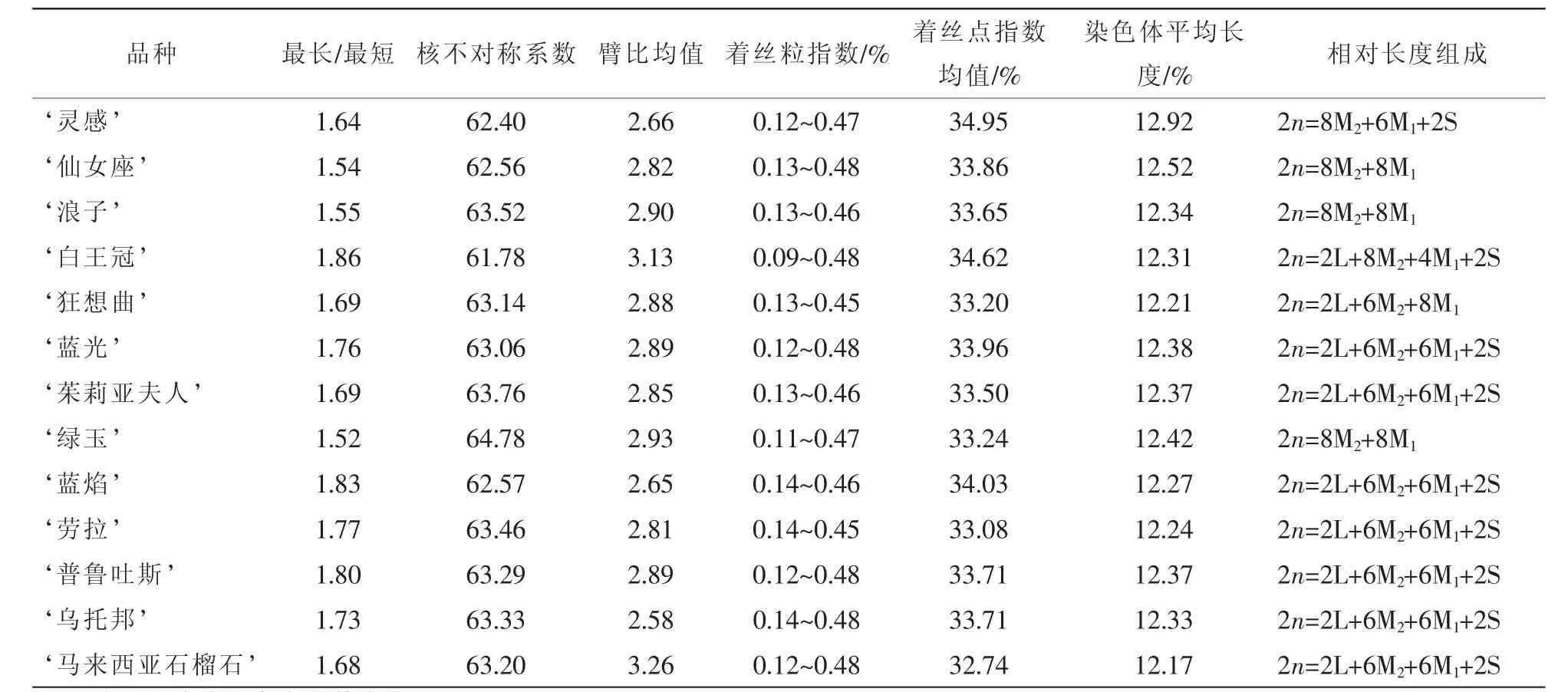
表3 13個鐵線蓮品種染色體參數比較Table 3 Chromosomes parameters of thirteen Clematis cultivars

圖2 13個鐵線蓮品種的染色體核型圖Figure 2 Karyotype of thirteen Clematis cultivars
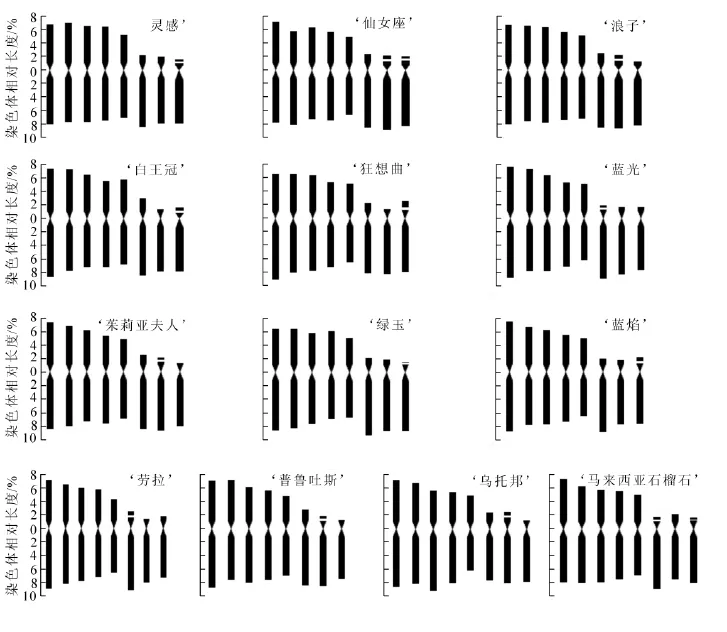
圖3 13個鐵線蓮品種的核型模式圖Figure 3 Chromosomes karyotype mode of thirteen Clematis cultivars
[1] 蔡艷飛,李世峰,李涵,等.中國鐵線蓮屬植物研究進展[J].中國農學通報,2009,25(4):195-198.
CAI Yanfei,LI Shifeng,LI Han,et al.Advance in the research of Climatis L.[J].Chin Agric Sci Bull,2009,25(4):
195-198.
[2] 張鐿鋰,賀士元.國產6種鐵線蓮的染色體研究[J].武漢植物學研究,1990,8(2):115-121.
ZHANG Yili,HE Shiyuan.Chromosome studies on 6 species of Clematis in China[J].J Wuhan Bot Res,1990,8(2): 115-121.
[3] 龔維忠,龍雅宜,李懋學.北京地區鐵線蓮屬植物的研究[J].武漢植物學研究,1985,3(4):371-379.
GONG Weizhong,LONG Yayi,LI Maoxue.Karyotype studies on Clematis from Beijing,China[J].J Wuhan Bot Res, 1985,3(4):371-379.
[4] 張鐿鋰.7種鐵線蓮的染色體研究[J].武漢植物學研究,1991,9(2):107-111.
ZHANG Yili.Chromosome studies on 7 species of Clematis in China[J].J Wuhan Bot Res,1991,9(2):107-111.
[5] 楊親二,羅毅波,洪德元.湖南7種毛茛科植物的核型研究[J].廣西植物,1994,14(1):27-36.
YANG Qinger,LUO Yibo,HONG Deyuan.A karyotype study of six species in the Ranunculaceae from Hunan,China[J].Guihaia,1994,14(1):27-36.
[6] 楊親二.國產毛茛科銀蓮花族17種植物的細胞學研究[J].植物分類學報,2002,40(5):396-405.
YANG Qinger.Cytology of ten species in Anemone,one in Anemoclema and six in Clematis(Trib.Anemoneae,Ranunculaceae)from China[J].Acta Phytotaxon Sin,2002,40(5):396-405.
[7] 彭綠春,于恒雋,余娜,等.10種鐵線蓮的核型特征及核型似近系數聚類分析[J].湖南農業大學學報:自然科學版,2012,38(6):617-622.
PENG Lüchun,YU Hengjuan,YU Na,et al.Karyotype features of 10 species of Clematis and cluster analysis of karyotypies resemblance-near coefficient[J].J Hunan Agric Univ Nat Sci,2012,38(6):617-622.
[8] 寧華.植物細胞核不同制備方法的比較研究[J].華中師范大學學報:自然科學版,2009,43(2):308-311.
NING Hua.Compared study on methods of plant nucleus preparation with different ways[J].J Huazhong Norm Univ Nat Sci,2009,43(2):308-311.
[9] 高猛,安玉麟,孫瑞芬.向日葵染色體Giemsa C-分帶研究[J].植物遺傳資源學報,2012,13(6):1078-1081.
GAO Meng,AN Yulin,SUN Ruifen.Giemsa C-banding patterns analysis of sunflower chromosomes[J].J Plant Genet Resour,2012,13(6):1078-1081.
[10] 李懋學,陳瑞陽.關于植物核型分析的標準化問題[J].武漢植物學研究,1985,3(4):297-302.
LI Maoxue,CHEN Ruiyang.A suggestion on the standardization of karyotype analysis in plants[J].J Wuhan Bot Res,1985,3(4):297-302.
[11] LEVAN A,FREDGA K,SANDBERG A A.Nomenclature for centromeric position on chromosomes[J].Hereditas, 1964,52(2):201-220.
[12] STEBBINS G L.Chromosome Evolution in Higher Plants[M].London:Edward Amold,1971.
[13] ARANO H.Cytological studies in subfamily Carduoidae(Comositae)of Japan(Ⅸ)the karyotype analysis and phylogenic consideration on Pertya and Ainsliaea[J].Bot Mag Tokyo,1963,76(895):32-39.
[14] KUO S R,WANG T T,HUANG T C.Karyotype analysis of some formosan gymnosperms[J].Taiwania,1972,17(1): 66-80.
[15] 盧玉飛,蔣建雄,易自力.利用Photoshop軟件處理芒屬植物染色體圖像的方法探討[J].草業科學,2013,30(6):922-926.
LU Yufei,JIANG Jianxiong,YI Zili.The protocol for processing of the chromosomal images of Misanthus for karyotype analysis using Adobe Photoshop software[J].Pratacultural Sci,2013,30(6):922-926.
[16] 劉泉,傅祖蕓.一種植物染色體圖像核型分析系統[J].計算機工程與應用,2000,36(3):71-74.
LIU Quan,FU Zuyun.A system of analyzing plant chromosome image’s karyotype[J].Comput Eng Appl,2000,36(3):71-74.
[17] 楊漢民.細胞生物學實驗[M].北京:高等教育出版社,1997.
[18] 楊光穗,尹俊梅,冷青云,等.鶴蕉屬植物8個品種間的核型比較分析[J].熱帶作物學報,2010,31(12):2118-2123.
YANG Guangsui,YIN Junmei,LENG Qingyun,et al.Karyotypes comparison of eight varieties of Heliconia spp.[J]. Chin J Trop Crops,2010,31(12):2118-2123.
Karyotypes in 13 Clematis cultivars
REN Jialun,JI Mengcheng,ZHAO Shuang,LIU Zhigao
(School of Landscape Architecture,Zhejiang A&F University,Lin’an 311300,Zhejiang,China)
Clematis has extremely high ornamental value,complicated genetic background and crossincompatibility phenomena.Studies on chromosome karyotype analysis of hybrid parent is very important. Chromosome types and karyotypes with natural growth of tender root tip materials in 13 Clematis cultivars were studied by the method of cell wall degradation and hypotonic treatment for chromosome preparations.Results indicated that all chromosomes for the 13 provenances were diploids and the chromosome number was 2n=2x =16.There were 2 types of chromosome karyotypes classified from the morphological observation:2n=2x= 16=10m+4st+2t and 2n=2x=16=10m+6st.Satellites were found in chromosomes of each cultivar for numbers 6,7,and 8.Both karyotypes were “2A”.Thus,both chromosome karyotypes had a great deal of similarity.So cross incompatibility phenomena may caused by other factors.[Ch,3 fig.3 tab.18 ref.]
plant cytogenetics;Clematis;chromosome;karyotype
S682.39;Q943
A
2095-0756(2016)06-1033-07
2015-12-22;
2016-02-12
浙江省科學技術重大資助項目(2012C12909-16)
任佳倫,從事野生園林植物資源分類與應用。E-mail:1416001262@qq.com。通信作者:劉志高,副教授,從事觀賞植物栽培與育種技術研究。E-mail:1692362952@qq.com
10.11833/j.issn.2095-0756.2016.06.015
毛茛科Ranunculaceae鐵線蓮屬Clematis為觀賞價值極高的藤本植物,全世界大約有300種,中國約110種,分布于華中西南地區。鐵線蓮屬植物花色美麗,花型特別,有 ‘攀援植物皇后’的美稱,是非常優秀的垂直綠化材料,因具多種抗逆性,也是雜交育種的優良資源[1]。早在1885年,就有科學家GURIGNARD對鐵線蓮屬植物進行了細胞學研究,證實C.recta的單倍染色體n=8,MEURMAN等在1939年研究發現了四倍體、六倍體與雜合染色體。據張鐿鋰等[2]的統計,該屬約有106種與變種的染色體數目及部分種的核型有過記載,其他并未見報道。對鐵線蓮屬的細胞學研究多集中在野生種上[2-6],而有關栽培品種的的研究則鮮有報道[7]。通過自然選擇及人工雜交選育,鐵線蓮品種數量繁多,形態變異豐富,遺傳背景也相對復雜,為鐵線蓮種質資源調查、品種分類與鑒定等工作帶來了一定的難處。針對雜交育種工作中存在子一代生長周期較長,且本研究的13個品種間存在雜交不親和性的障礙,因此,
對雜交親本的染色體核型分析等細胞學特征的研究顯得十分重要。本研究采用酶解法、去壁低滲火焰干燥法和Giemsa-C分帶技術對13個鐵線蓮品種進行核型分析,研究核型差異是否是導致其雜交不親和原因。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
