吉林西部鹽堿地水田水稻不同生長期土壤酶活性與有機碳含量的關系
2016-12-19 03:42:03湯潔趙晴王思寧王靜靜孫珊珊
生態環境學報 2016年9期
關鍵詞:水稻
湯潔,趙晴,王思寧,王靜靜,孫珊珊,
1. 吉林大學環境與資源學院,吉林 長春 130012;2. 吉林省環境監察總隊,吉林 長春 130033
吉林西部鹽堿地水田水稻不同生長期土壤酶活性與有機碳含量的關系
湯潔1,趙晴1,王思寧1,王靜靜1,孫珊珊1,2
1. 吉林大學環境與資源學院,吉林 長春 130012;2. 吉林省環境監察總隊,吉林 長春 130033
土壤酶在土壤有機碳的轉化過程具有關鍵作用,研究鹽堿地水田土壤酶與SOC的關系對深入了解此類土壤碳循環機制有著重要意義。本文選取吉林省西部前郭縣典型鹽堿水田作為研究區,分別于水稻未插秧期、幼苗期、分蘗期、抽穗期和結實期采集0~30 cm和30~60 cm的土壤樣品。樣品采集后帶回實驗室進行測試,分別采用高錳酸鉀容量法測定過氧化氫酶活性,3,5-二硝基水楊酸比色法測定淀粉酶活性,總有機碳分析儀測定SOC含量,分析土壤過氧化氫酶、淀粉酶活性和SOC含量的剖面分異特征及變化規律,并探討酶與SOC的相關性。結果表明:鹽堿水田土壤過氧化氫酶、淀粉酶活性以及SOC含量均隨土壤剖面深度的加深而降低,且隨水稻的生長而波動。插秧前,表層土壤過氧化氫酶、淀粉酶分別與SOC呈極顯著正相關(n=18,P<0.01)和顯著正相關(n=18,P<0.05),SOC含量不僅與兩種酶活性密切相關,并受過氧化氫酶活性影響更大。水稻移栽后,兩種酶與SOC的相關性均有不同程度降低,過氧化氫酶活性在幼苗期、抽穗期和結實期與SOC均呈顯著正相關(n=18,P<0.05);淀粉酶活性在幼苗期、分蘗期和結實期與SOC呈顯著正相關(n=18,P<0.05)。土壤過氧化氫酶、淀粉酶在水稻生長發育過程中受到外界干擾以及水稻根系發育等影響,活性發生明顯變化,進而影響 SOC。由于抽穗期水稻生長發育緩慢,對淀粉酶分解作用需求降低,使得其活性減弱,故對SOC的影響最小。
鹽堿水田;生長期;土壤酶;土壤有機碳
土壤擁有最大的陸地有機碳庫(Reichstein et al.,2008;Van Groenigen et al.,20141-2),據估計(Falkowski et al.,2000;解憲麗等,2004),全球土壤有機碳(soil organic carbon,SOC)庫介于1395~2200 Pg(1 Pg=1015g),是陸地植被碳庫(500~600 Pg)的2~3倍,是全球大氣碳庫(750 Pg)的2倍多。由于土壤有機碳貯量庫容巨大,其微小的變化就會影響到大氣中的碳量,對調制氣候變化具有關鍵性的作用(Van Groenigen et al.,20141-2;陳朝等,2011)。SOC含量受到多種因素的影響,其中一個重要的因子就是土壤酶活性。土壤酶參與了土壤中的生物化學過程,與生態系統中營養物質循環等密切相關(關松蔭,1986)61-64。SOC的分解轉化幾乎與所有的土壤酶活性有關,因此,土壤酶在土壤碳循環中有著舉足輕重的作用,研究其活性與土壤有機碳含量的相關關系是十分必要的。目前國內外學者對 SOC和土壤酶的研究,主要集中在兩者的變化規律以及影響因素方面,而對兩者相關性的研究較少,且研究多見于森林、草原、濕地、農田等生態系統(Jiang et al.,2014;Ye et al.,2014;Zhang et al.,2015;萬忠梅等,2008a;孫文娟等,2008;吳旭東等,2013),對鹽堿水田生態系統的研究則少見。此外,以往的研究一般只集中研究水稻的某一生長期,而本研究將水稻各個不同生長期(未插秧期、幼苗期、分蘗期、抽穗期、結實期)均納入研究范圍。
吉林西部位于松嫩平原西南側,科爾沁草原東部,屬于全球變化的中國東北樣帶,是生態脆弱帶(李忠和,2014),也是世界三大鹽堿地集中分布區之一(楊福等,2007)。為治理蘇打鹽堿地,該區開展了多項研究,得出種植水稻是改良利用蘇打鹽堿地的有效方法之一的結論(趙國臣等,2012)。自20世紀50年代開始,水田改造工程初步實施,從而形成具有吉林西部特色的鹽堿水田生態系統。本研究以吉林西部前郭爾羅斯蒙古族自治縣(以下簡稱前郭縣)鹽堿水田為研究對象,選取與土壤碳循環密切相關的過氧化氫酶和淀粉酶進行研究。其中,過氧化氫酶分解土壤中過氧化氫,有利于防止過氧化氫對生物體的毒害作用,其活性表征土壤腐殖化強度大小和有機質積累程度(萬忠梅等,2008b);土壤淀粉酶可將淀粉水解為麥芽糖,后在麥芽糖酶的作用下水解成葡萄糖,從而為植物提供營養,其與土壤有機質的礦化分解息息相關(萬忠梅等,2009)。同步監測 SOC,探究水稻生長過程中土壤酶活性和土壤有機碳含量的變化特征以及兩者的關系,細化并完善鹽堿水田區水稻生長期土壤酶活性與 SOC關系的研究,以期為進一步研究吉林西部鹽堿水田生態系統碳循環機制提供基礎數據。
1 材料與方法
1.1 土壤樣品的采集與前處理
研究區位于吉林省前郭縣的鹽堿水田區,分別于2014年4月18—22日、6月8—11日、7月5—9日、8月12—16日、9月25—28日采集土壤樣品,分別對應插秧前(Ⅰ)、幼苗期(Ⅱ)、分蘗期(Ⅲ)、抽穗期(Ⅳ)和結實期(Ⅴ)5個生長期。選取 3個水田樣地,分別設為 S1(124.717°E,45.005°N)、S2(124.686°E,45.006°N)和 S3(124.714°E,45.000°N),按照對角線布點法均勻采集0~30 cm表層土、30~60 cm底層土樣品(理化性質見表1),每層采集3個平行樣混合裝入密封塑料袋中帶回實驗室,去除土壤中的動植物殘體和石塊,自然風干后碾碎壓細,過0.20 mm篩備用。3個樣點的施肥和管理措施一致,均為4月初進行犁地,5月初灌水,5月8日和6月20日各施1次氮肥(尿素)。
1.2 土壤樣品的測試與數據分析插秧
過氧化氫酶活性的測定采用高錳酸鉀容量法(楊蘭芳等,2011),單位以mgg-1表示;淀粉酶活性采用3, 5-二硝基水楊酸比色法測定(關松蔭,1986278-280;楊蘭芳等,2011),單位以 mgg-1表示;SOC采用島津TOC-V系列總有機碳分析儀的SSM-5000A固體樣品模塊進行測定,單位以%表示。
每個樣品選取3個平行樣進行測定,數據采用平均值,所有數據采用Microsoft Excel 2003軟件進行統計整理和繪圖,利用SPSS 19.0進行方差分析。
2 結果與分析
2.1 土壤酶活性變化特征
鹽堿水田區水稻土過氧化氫酶和淀粉酶空間剖面變化規律見圖1。在水稻不同生長期,土壤過氧化氫酶活性和淀粉酶活性均隨土壤深度的增加而減弱,造成這種現象的主要原因是二者均來源于植物根系的分泌以及真菌和細菌的分解作用,90%的水稻根系分布在表層土壤中(劉建,2013),且大量研究(盛浩等,2013;Sotomayor-Ramírez et al.,2009;Cui et al.,2015;肖燁等,2015)表明微生物生物量在表層土壤中含量最多。另外,本區鹽堿水田土壤的特殊性,即底層pH、粘粒含量、土壤容重、含鹽量明顯高于表層土壤,抑制了底層土壤酶的活性,因此,表層酶活性高于底層酶活性。
從圖1可知,表層土壤過氧化氫酶、淀粉酶的活性均隨水稻生長期呈“減-增-減-增”的變化規律,但淀粉酶的變化幅度略大。水稻移栽后,其根系受到損傷,生長滯緩,釋放的土壤酶減少,同時土壤中微生物受到干擾,使得幼苗期酶活性降低(楊安中,2014;馬偉等,2015);分蘗期是水稻根系生長的主要時期(王伯倫,2010),此時,水稻根系將分泌出更多的有機酸和碳水化合物,刺激過氧化氫酶和淀粉酶活性的提高(鄭丕堯,1992)。另外,分蘗期采樣時間為7月5—9日,與第二次施肥時間(6月20日)較近,這一時期酶活性增高可能也受到了氮肥影響。抽穗期,水稻各種營養器官逐漸停止生長,整個水稻系統處于不活躍狀態,且根系生長活動所需的各種營養基本都由分蘗節上的近葉提供(張洪程,2011),不再過多依賴土壤,故兩種酶活性呈現減弱趨勢;結實期是水稻米粒生長和充實期,是水稻生長的又一活躍期,因此兩種酶的活性又呈增強的趨勢。
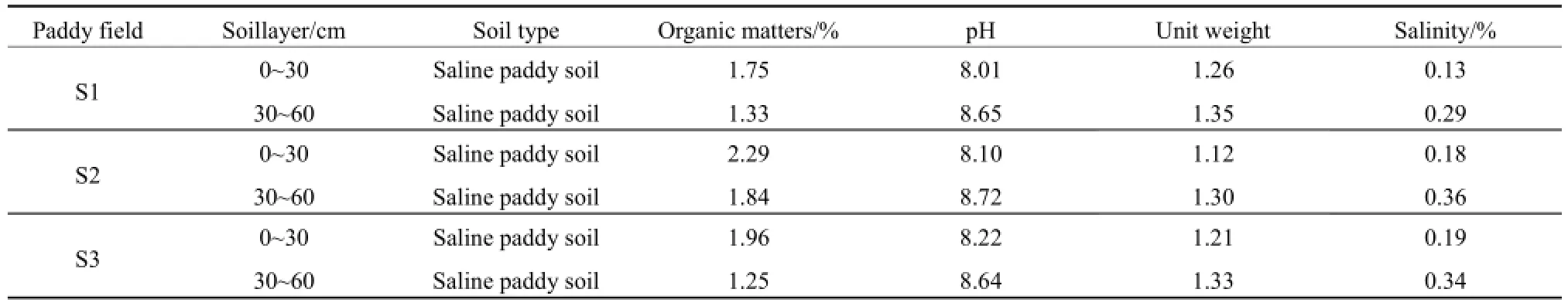
表1 吉林西部鹽堿地水田土壤基本理化性質Table 1 Basic physico-chemical properties of the saline paddy field in west Jilin
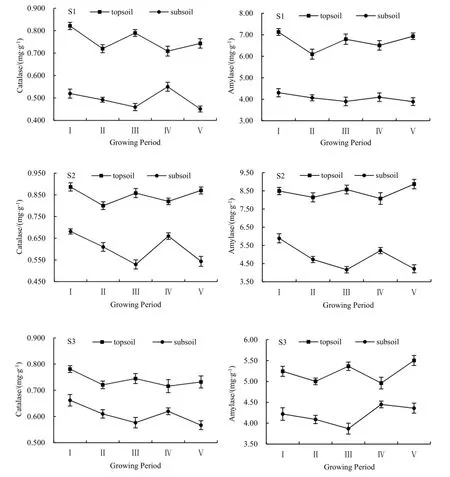
圖1 吉林西部鹽堿地水田水稻不同生長期土壤酶變化特征Fig. 1 Soil enzymes activities of the saline paddy field in west Jilin during rice growing
對于底層土壤,過氧化氫酶和淀粉酶活性隨水稻生長均呈“減-減-增-減”的變化規律。從幼苗期到分蘗期,底層土壤酶活性均處于較低的狀態。可能是因為這兩個時期水稻根系主要分布在表層土壤,致使底層土壤有機質含量和微生物活躍度均較低,所以,土壤酶活性不高。抽穗期水稻進入穩定狀態,伴隨著根系不斷向下層土壤發育(王伯倫,2010),土壤中微生物開始向底層活躍,底層土壤酶活性升高。進入結實期后,水稻開始結籽,微生物又逐漸集中在表層土壤,底層酶活性降低。
2.2 土壤有機碳含量變化特征
由圖2可得,在水稻生長的各個時期表層SOC含量均高于底層土壤。表層土壤因有植物凋落物或殘體對其補充有機質,且水熱條件適宜,故表層有機殘體分解情況較好(趙仁竹等,2015)。同時,表層土壤根系在發育的過程中會分泌有機質,故表層 SOC含量較高。底層土壤受外界干擾較小,不僅來自外界的有機碳補充量偏少,而且不發育的根系分泌的有機質也較少,因此,其 SOC含量比表層低。
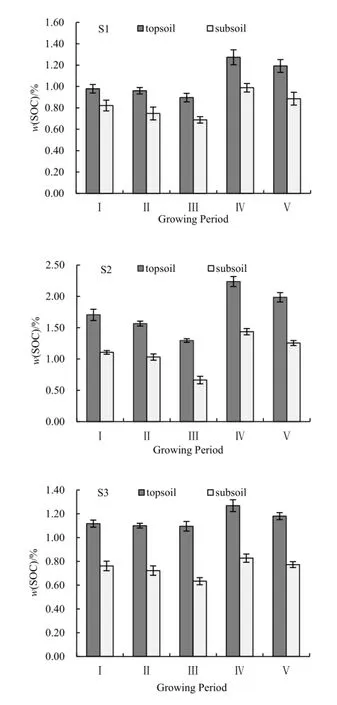
圖2 吉林西部鹽堿地水田水稻不同生長期土壤有機碳變化特征Fig. 2 Changes of SOC of the saline paddy field in west Jilin
從圖2可看出,表層和底層SOC基本都呈“減-減-增-減”的變化規律。SOC含量在幼苗期和分蘗期下降的主要原因是,水稻在插秧后進入生長發育期,需要大量的營養物質,SOC在酶的作用下不斷礦化分解,從而為水稻提供養分(張豪等,2013)。抽穗期水稻生長發育緩慢,需要的營養物質減少,且所需的營養物質可由葉片的光合作用提供,故土壤 SOC含量增加。結實期水稻籽粒的生長發育需要礦質元素,SOC在礦化分解過程中自身含量下降。分蘗期是 SOC含量下降幅度最大的時期,此時水稻處于最活躍的生長階段,需要的營養物質最多,故SOC含量亦最低。
2.3 水稻不同生長期土壤酶活性與土壤有機碳含量的關系
為了研究水稻不同生長期土壤酶活性與 SOC的關系,對水稻每個生長期的土壤酶活性與有機碳含量逐一進行了相關分析(表2)。插秧前,過氧化氫酶與SOC的相關系數為0.895(n=18,P<0.01),淀粉酶與 SOC的相關系數為 0.812(n=18,P<0.05)。可見,在插秧前SOC的含量對這兩種酶活性的影響顯著,并且過氧化氫酶的變化對 SOC的響應更大。土壤過氧化氫酶在催化過氧化氫分解的過程中,會釋放出氧氣,有利于土壤中好氧微生物的生命活動以及酶促反應中產物水的生成,從而增大土壤腐殖化強度,提高SOC含量(關松蔭,1986)。而淀粉酶只是催化有機物質使土壤中SOC的含量增加,并不像過氧化氫酶那樣對土壤環境產生較大影響。
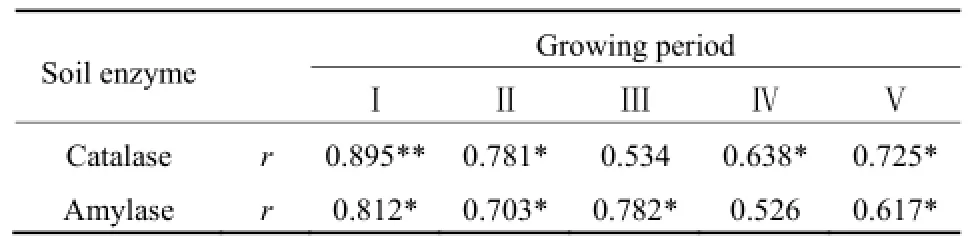
表2 吉林西部鹽堿地水田水稻不同生長期土壤酶與有機碳的相關關系Table 2 Relationship between soil enzymes activities and TOC of the saline paddy field in west Jilin
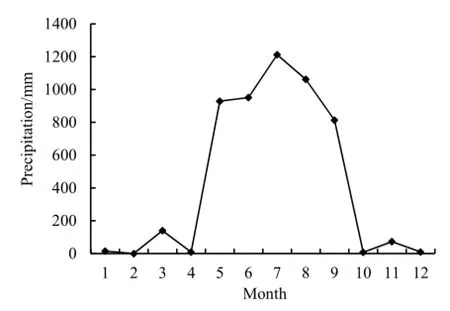
圖3 前郭縣2014年月平均降水量Fig. 3 Average monthly precipitation of Qianguo in 2014
水稻移栽后,土壤酶活性與 SOC的相關性發生了明顯變化,但兩種酶的變化規律有所差異。從水稻的幼苗期到結實期,土壤過氧化氫酶與 SOC的相關系數分別為0.781、0.534、0.638和0.725。其中,土壤過氧化氫酶在分蘗期與 SOC無顯著相關性,這是由于該區6—7月為豐水期(見圖3),在降雨增加稻田積水的同時,土壤逐漸積累了過氧化氫,過氧化氫酶活性也就隨之增大;雖然過氧化氫酶會間接促進 SOC的積累,但是,分蘗期是水稻生長最旺盛的時期,需要大量的 SOC礦化分解提供養分,故 SOC含量下降。在水稻生長的其他時期,兩者關系均呈顯著正相關(n=18,P<0.05),但相關系數明顯比未插秧時期小,其原因為水稻生長發育,特別是根部的活動會促進過氧化氫酶活性的增強,同時水稻生長需要大量營養物質,又會消耗SOC,因此,兩者正相關性下降。
土壤淀粉酶和 SOC的相關性隨著水稻的生長也發生了明顯的變化,除了分蘗期,兩者的相關系數與插秧前相比,均呈明顯下降趨勢。其中,幼苗期兩者相關系數為0.703,抽穗期為0.526,結實期為0.617。在幼苗期和結實期,土壤淀粉酶與SOC均呈顯著正相關(n=18,P<0.05)。但在不同生長期土壤過氧化氫酶活性和 SOC的變化并不完全一致,抽穗期時兩者相關系數最小,無明顯相關性,因為抽穗期水稻生長發育變慢,對淀粉酶分解作用需求降低,使得淀粉酶活性明顯減小,而 SOC在這一時期累積量卻最大。分蘗期兩者的相關系數高于水稻生長過程中的其他時期,與未插秧時期基本相同。
3 結論
(1)鹽堿水田區表層土的 SOC含量、過氧化氫酶和淀粉酶活性最高,受到水稻根系生長、微生物活性以及周圍環境的影響,不同生長期,其變化規律有所不同。SOC含量在抽穗和結實期明顯增高,說明水稻種植是土壤碳匯過程,可有效減少大氣中二氧化碳的含量。
(2)水稻種植前后,土壤過氧化氫酶、淀粉酶活性與 SOC的相關性均有不同程度降低。除分蘗期和抽穗期外,兩種酶活性均與 SOC呈現顯著正相關。分蘗期降水增多、抽穗期水稻生長發育減緩分別是影響過氧化氫酶、淀粉酶與 SOC相關性的主要因素。
(3)隨著水稻的種植,鹽堿水田區的表土層不僅有機質不斷積累,而且土壤 pH、粘粒含量、土壤容重和含鹽量都降低了。土壤根系發育、酶活性高、含鹽量低是表土有機碳富集的重要因素。
CUI J, HOLDEN N. 2015. The relationship between soil microbial activity and microbial biomass, soil structure and grassland management [J]. Soil and Tillage Research, 146(Part A): 32-38. FALKOWSKI P, SCHOLES R J, BOYLE E, et al. 2000. The global carbon cycle: a test of our knowledge of earth as a system [J]. Science, 290(5490): 291-296.
JIANG G, XU M, HE X, et al. 2014. Soil organic carbon sequestration in upland soils of northern China under variable fertilizer management and climate change scenarios [J]. Global Biogeochemical Cycles, 28(3): 319-333.
REICHSTEIN M, HEIM ANN M. 2008. Terrestrial ecosystem carbon dynamics and climate feedbacks [J]. Nature, 451(7176): 289-292.
SOTOMAYOR-RAMíREZ D, ESPINOZA Y, ACOSTA-MARTíNEZ V. 2009. Land use effects on microbial biomass C, β-glucosidase and β-glucosaminidase activities, and availability, storage, and age of organic C in soil [J]. Biology and Fertility of Soils, 45(5):487-497.
VAN GROENIGEN K J, QI X, OSENBERG C W, et al. 2014. Faster decomposition under increased atmospheric CO2limits soil carbon storage [J]. Science, 344(6183): 508-509.
YE X, LIU H, LI Z, et al. 2014. Effects of green manure continuous application on soil microbial biomass and enzyme activity [J]. Journal of Plant Nutrition, 37(4): 498-508.
ZHANG X, DONG W, DAI X, et al. 2015. Responses of absolute and specific soil enzyme activities to long term additions of organic and mineral fertilizer [J]. Science of the Total Environment, 536: 59-67.
陳朝, 呂昌河, 范蘭, 等. 2011. 土地利用變化對土壤有機碳的影響研究進展[J]. 生態學報, 31(18): 5358-5371.
關松蔭. 1986. 土壤酶及其研究法[M]. 北京: 農業出版社.
韓永偉, 郭彥君, 張蘊薇, 等. 2005. 農牧交錯帶退耕還草對土壤淀粉酶和脲酶活性的影響[J]. 草地學報,13(1): 59-62, 74.
解憲麗, 孫波, 周慧珍, 等. 2004. 不同植被下中國土壤有機碳的儲量與影響因子[J]. 土壤學報, 41(5): 687-699.
李忠和. 2014. 吉林西部稻田土壤微生物及酶活性對碳變化響應機制研究[D]. 長春: 吉林大學: 2.
劉建. 2013. 優質水稻高產高效栽培技術[M]. 北京: 中國農業科學技術出版社: 10.
馬偉, 霍志國, 姬靜華, 等. 2015. 殼聚糖和水楊酸混配液對移栽返青期水稻幼苗抗寒性影響及機理[J]. 中國稻米, 21(4): 150-154.
盛浩, 周萍, 袁紅, 等. 2013. 亞熱帶不同稻田土壤微生物生物量碳的剖面分布特征[J]. 環境科學, 34(4): 1576-1582.
孫文娟, 黃耀, 張穩, 等. 2008. 農田土壤固碳潛力研究的關鍵科學問題[J]. 地球科學進展, 23(9): 997-1002.
萬忠梅, 宋長春. 2008a. 三江平原不同類型濕地土壤酶活性及其與營養環境的關系[J]. 水土保持學報, 22(5): 158-161.
萬忠梅, 宋長春. 2008b. 小葉章濕地土壤酶活性分布特征及其與活性有機碳表征指標的關系[J]. 濕地科學, 6(2): 249-257.
萬忠酶, 宋長春. 2009. 土壤酶活性對生態環境的響應研究進展[J]. 土壤通報, 40(4): 952-956.
王伯倫. 2010. 水稻栽培技術[M]. 沈陽: 東北大學出版社: 17.
吳旭東, 張曉娟, 謝應忠, 等. 2013. 不同種植年限紫花苜蓿人工草地土壤有機碳及土壤酶活性垂直分布特征[J]. 草業學報, 22(1): 245-251.
肖燁, 黃志剛, 吳海濤, 等. 2015. 三江平原典型濕地類型土壤微生物特征與土壤養分的研究[J]. 環境科學, 36(5): 1842-1848.
楊安中. 2014. 水稻高產實用技術[M]. 合肥: 安徽大學出版社: 10.
楊福, 梁正偉. 2007. 關于吉林省西部鹽堿地水稻發展的戰略思考[J].北方水稻, (6): 7-12.
楊蘭芳, 曾巧, 李海波, 等. 2011. 紫外分光光度法測定土壤過氧化氫酶活性[J]. 土壤通報, 42(1): 207-210.
張豪, 湯潔, 梁爽. 2013. 吉林西部不同開發年份鹽堿水田土壤有機碳和碳酸鹽的季節動態[J]. 生態環境學報, 22(12): 1899-1903.
張洪程. 2011. 水稻新型栽培技術[M]. 北京: 金盾出版社: 20-25.
趙國臣, 齊春艷, 侯立剛, 等. 2012. 吉林省蘇打鹽堿地水稻生產歷史進程與展望[J]. 沈陽農業大學學報, 43(6): 673-680.
趙仁竹, 湯潔, 梁爽, 等. 2015. 吉林西部鹽堿田土壤蔗糖酶活性和有機碳分布特征及其相關關系[J]. 生態環境學報, 24(2): 244-249.
鄭丕堯. 1992. 作物生理學導論[M]. 北京: 北京農業大學出版社.
Correlation of Soil Enzyme Activities with Soil Organic Carbon Content in Saline-alkali Paddy Field
TANG Jie1, ZHAO Qing1, WANG Sining1, WANG Jingjing1, SUN Shanshan1,2
1. College of Environmental and Resources, Jilin University, Changchun 130021, China; 2. Environmental Inspection Corps of Jilin Province, Changchun 130033, China
Soil enzymes play a critical role in soil organic carbon transformation, so study on correlation between enzymes and SOC of Saline-alkali paddy field is important for us to research carbon cycle mechanism more systematically. We selected Saline-alkali paddy field in Qianguo county of western Jilin province as a research area, collecting samples of topsoil (0~30 cm) and subsoil (30~60 cm) in not planting rice, seedling stage, tillering stage, tassel formation and fructicative stage respectively. The activities of catalase and amylase were measured by KMnO4capacity method and 3, 5-dinitrosalicylic acid colorimetry respectively, and SOC were measured with a total organic carbon analyzer, to analyze the distribution of SOC, soil catalase activity and soil amylase activity in different rice growth stages, furthermore to explore the correlation between soil enzyme and SOC. The results showed that soil catalase activity, soil amylase activity and SOC content all reduced significantly with increasing soil depth, and also fluctuated with rice growing Before transplanting of rice, SOC extremely significantly correlated with soil catalase and significantly with soil amylase, respectively (P<0.01, P<0.05); SOC content was not only closely related to the two soil enzymes but also influenced more by catalase. After transplanting, the positive correlation between them all decreased in varying degrees. SOC had a significant positive correlation with catalase in seedling stage, tassel formation and fructicative stage (P<0.05), with amylase activity in seedling stage, tillering stage and fructicative stage (P<0.05). Soil catalase and amylase interfered by surroundings and development of rice root system during the rice growth caused the significant change of activity, which further affected SOC. As rice growed slowly during tassel formation, its demand of amylase to decompose organic matter diminished, leading to the decline of its activity, so it had the least impact on SOC.
saline-alkali paddy field; growth stages; soil enzyme; SOC
10.16258/j.cnki.1674-5906.2016.09.001
S15; X144
A
1674-5906(2016)09-1425-06
湯潔, 趙晴, 王思寧, 王靜靜, 孫珊珊. 2016. 吉林西部鹽堿地水田水稻不同生長期土壤酶活性與有機碳含量的關系[J]. 生態環境學報, 25(9): 1425-1430.
TANG Jie, ZHAO Qing, WANG Sining, WANG Jingjing, SUN Shanshan. 2016. Correlation of soil enzyme activities with soil organic carbon content in saline-alkali paddy field [J]. Ecology and Environmental Sciences, 25(9): 1425-1430.
國家自然科學基金項目(41471152);博士點基金項目(20130061110065)
湯潔(1957年生),女,教授,博士生導師,主要從事生態環境系統工程理論與技術研究。E-mail: tangjie0724@163.com
2016-07-04
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
