新疆小麥 TaLox-B1、 TaLox-B2、 TaLox-B3位點的等位變異及其分布規律
2016-12-29 01:44:23禹飛雄茹仙古麗牙生呂文娟耿洪偉
麥類作物學報 2016年11期
關鍵詞:新疆
白 璐,禹飛雄,茹仙古麗·牙生,呂文娟,任 毅,謝 磊,耿洪偉
(1.新疆農業大學科學技術學院,新疆烏魯木齊 830052; 2.新疆農業大學農學院,新疆農業大學生物技術重點實驗室,新疆烏魯木齊 830052; 3.新疆巴州農業局,新疆庫爾勒 841000)
新疆小麥 TaLox-B1、 TaLox-B2、 TaLox-B3位點的等位變異及其分布規律
白 璐1,禹飛雄2,茹仙古麗·牙生3,呂文娟2,任 毅2,謝 磊2,耿洪偉2
(1.新疆農業大學科學技術學院,新疆烏魯木齊 830052; 2.新疆農業大學農學院,新疆農業大學生物技術重點實驗室,新疆烏魯木齊 830052; 3.新疆巴州農業局,新疆庫爾勒 841000)
面粉顏色是小麥品質分級的重要指標。為了給新疆小麥面粉白度改良提供依據,本研究利用位于4BS染色體上的功能標記LOX16、LOX18和Lox-B23,對123份新疆小麥品種(系)的 TaLox-B1、 TaLox-B2和 TaLox-B3 等位變異進行分子標記檢測。結果表明,在123份新疆小麥品種(系)中,具有高LOX活性等位基因 TaLox-B1a和低LOX活性等位基因 TaLox-B1b的材料分別有33份(26.8%)和90份(73.2%);具有高LOX活性等位基因 TaLox-B2a和低LOX活性等位基因 TaLox-B2b的材料分別有122份(99.2%)和1份(0.8%);具有高LOX活性等位基因 TaLox-B3a和低LOX活性等位基因 TaLox-B3b的材料分別有95份(77.2%)和28份(22.8%)。在不同類型冬小麥品種(系)中, TaLox-B1a的分布頻率表現為引進品種(系)>自育品種(系)>地方品種外, TaLox-B3a的分布頻率表現為地方品種>引進品種(系)>自育品種(系);在不同類型春小麥中 TaLox-B1a只存在于晚期品種, TaLox-B2a和 TaLox-B3a的分布頻率均表現為早期品種(100.0%)>晚期品種。所檢測的新疆小麥品種(系)共有5種基因型組合,具有最高LOX活性的基因型組合 TaLox-B1a/ TaLox-B2a/ TaLox-B3a和 TaLox-B1b/ TaLox-B2a/ TaLox-B3a,具有中等LOX活性的基因型組合 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b以及具有低LOX活性的基因型組合 TaLox-B1b/ TaLox-B2b/ TaLox-B3b的分布頻率分別為23.6%、53.7%、19.5%、2.4%和0.8%。在新疆小麥資源中具有高LOX活性基因型組合的材料的頻率高于全國平均水平。本研究將3個位點功能標記組合起來使用,能更有效地評價檢測材料的LOX活性水平。
新疆小麥;脂肪氧化酶;功能標記;變異組合
小麥面粉顏色是小麥品質分級的重要指標。小麥籽粒中脂肪氧化酶(lipoxygenase,LOX)活性是影響面粉色澤的重要因素之一,高LOX活性可以提高面粉及其相關產品的白度從而提高其商品價值[1-3]。LOX能夠與小麥面粉中的類胡蘿卜素發生偶聯氧化作用降低黃色素含量,進而使小麥面粉變白[4]。還有研究認為,低 LOX 活性更有助于小麥的貯藏[1],高LOX活性則有助于增強面筋強度[5]。
LOX活性受基因型和環境的影響,但主要受遺傳因素的影響[6]。前人研究表明,小麥籽粒LOX活性主要受第4和第5同源群上的基因調控[7-10]。Geng等[11]研究表明,在1AL上的 QLpx.caas-1AL位點和4BS上的 QLpx.caas-4B位點上存在決定LOX活性的2個主效基因,分別與SSR標記 Xwmc312和 Xgwm251緊密連鎖。基于4BS上的 TaLox-B1位點基因不同等位變異的序列差異,Geng等[12]開發了互補顯性標記LOX16和LOX18,其中,LOX16可在具有等位基因 TaLox-B1a(a指與高LOX活性相關,下同)的材料中擴增出489 bp的片段,而在具有等位基因 TaLox-B1b(b指與低LOX活性相關,下同)的材料中無擴增產物;LOX18在具有等位基因 TaLox-B1b的材料中可擴增出791 bp的片段,而在具有等位基因 TaLox-B1a的材料中無擴增產物。利用這一對互補顯性標記,Geng等[12]檢測198份中國冬小麥材料,結果表明這一對功能標記能有效應用于分子標記輔助選擇。張鈺玉等[13]、相吉山等[14]和楊淑萍等[15]利用互補顯性標記LOX16和LOX18分別對陜西、新疆和黑龍江小麥材料進行了等位變異檢測,結果表明該標記能有效應用于小麥分子育種實踐。隨后,Zhang等[16]在中國小麥4BS上成功克隆了 TaLox-B2、 TaLox-B3基因,并發現 TaLox-B2基因存在 TaLox-B2a和 TaLox-B2b 兩個等位變異,其將 TaLox-B3 基因類型定為 TaLox-B3a變異類型,不含有 TaLox-B3基因的暫定為 TaLox-B3b類型。基于這兩個LOX基因,Zhang等[16]開發了共顯性標記Lox-B23,能準確區分 TaLox-B2和 TaLox-B3位點的不同等位基因。 Lox-B23在具有 TaLox-B2a/TaLox-B3a基因型的材料中能同時擴增出788 bp和677 bp兩種帶型,在具有 TaLox-B2a/TaLox-B3b的材料中只能擴增出788 bp的單一帶型,而在具有 TaLox-B2b/TaLox-B3b的材料中只能擴增出660 bp的單一帶型。對136份中國小麥材料的 TaLox-B1、 TaLox-B2、 TaLox-B3位點進行等位變異檢測及LOX活性分析表明,具有最高LOX活性的基因型組合是 TaLox-B1a/TaLox-B2a/TaLox-B3a或 TaLox-B1b/TaLox-B2a/TaLox-B3a,具有中高LOX活性的基因型組合是 TaLox-B1b/TaLox-B2a/TaLox-B3b或 TaLox-B1a/TaLox-B2a/TaLox-B3b,具有最低LOX活性的基因型組合是 TaLox-B1a/TaLox-B2b/TaLox-B3b[16]。
雖然相吉山等[14]利用互補顯性標記LOX16和LOX18對新疆小麥種質資源 TaLox-B1位點分布特征做了研究,但目前尚未見有關新疆小麥 TaLox-B2、 TaLox-B3等位變異檢測以及三個位點組合基因型的檢測及其分布規律的研究報道。因此,本研究利用與LOX 活性相關的LOX16、LOX18和Lox-B23功能標記,對123份新疆麥區具有代表性的品種(系)的三個LOX活性基因 TaLox-B1、 TaLox-B2和 TaLox-B3的等位變異及其分布規律進行研究,以期為改良新疆面粉和面制品白度以及選育高LOX活性品種提供依據。
1 材料與方法
1.1 材料及其來源
參試材料為123份新疆小麥品種(系),其中,冬小麥品種(系)99份(17份為新疆地方品種,39份為引進品種,43份為自育品種),春小麥品種(系)24份(2000年前選育的早期品種4份,2000年后選育的晚期品種20份)。這些材料涵蓋新疆主要種植區各個時期種植和使用的冬、春小麥品種,具有較好的代表性。上述小麥材料由新疆農業大學農學院小麥課題組收集保存,其中部分小麥材料由新疆農科院糧食作物研究所冬小麥課題組、新疆農墾科學院作物研究所、新疆農科院核能與生物技術研究所小麥課題組提供。
1.2 基因組DNA的提取
為避免由于單粒種子偶然性的結果錯誤,每份小麥品種(系)均選取3粒種子,按照Lagudah等[17]的方法提取基因組DNA。DNA提取液包含:200 mmol·L-1Tris-HCl(pH=8.0),25 mmol·L-1EDTA,288 mmol·L-1NaCl 和0.5% SDS。
1.3 PCR擴增與檢測
1.3.1 引物
利用Geng等[12]開發的顯性標記LOX16和LOX18檢測小麥4B染色體上 TaLox-B1基因的等位變異;利用Zhang等[16]開發的共顯性標記 Lox-B23檢測小麥4B染色體上 TaLox-B2和 TaLox-B3基因的等位變異。引物(表1)均由北京六合華大基因科技股份有限公司合成。
表1 檢測目標基因等位變異的引物信息
Table 1 Primers for detecting alleles of target genes

基因Gene標記Marker引物序列(5'-3')Primersequence(5'-3')產物大小Productsize/bp等位變異Allele參考文獻ReferenceTaLox-B1LOX16F:CCATGACCTGATCCTTCCCTTR:GCGCGGATAGGGGTGGT489TaLox-B1a[12]LOX18F:GCGCGGATAGGGGTGGTR:GCGCGGATAGGGGTGC791TaLox-B1b[12]TaLox-B23Lox-B23F:TTCCCGCTCCAGCTCATCAAR:AGGGCCTGCATGCAACAAGT788/677788660TaLox-B2a/TaLox-B3aTaLox-B2a/TaLox-B3bTaLox-B2b/TaLox-B3b[16]
1.3.2 PCR的反應體系和擴增程序
PCR反應以基因組DNA為模板,在Eppendorf AG 22331 Hamburg PCR儀上進行。25 μL PCR反應體系:50 ng DNA,TaqDNA Polymerase (2.5 U·μL-1)0.25 μL (北京天根生化科技公司),上、下游引物(10 μmol·L-1)各1 μL,dNTP(25 μmol·L-1) 1 μL(北京天根生化科技公司),10×PCR Buffer 2.5 μL(北京天根生化科技公司),最后用ddH2O補充至25 μL。標記LOX16、LOX18的PCR擴增程序:94 ℃預變性5 min;94 ℃變性45 s,62 ℃退火45 s,72 ℃延伸1 min,共35個循環;最后72 ℃延伸10 min,4 ℃保存。標記Lox-B23的擴增程序:94 ℃預變性4 min;94 ℃變性45 s,退火溫度從65 ℃開始每循環降0.3 ℃,退火時間30 s,72 ℃延伸2 min,共35個循環;最后72 ℃延伸10 min,4 ℃保存。
1.3.3 瓊脂糖凝膠電泳檢測
標記LOX16、LOX18的PCR擴增產物用1.5%的瓊脂糖凝膠電泳檢測,標記Lox-B23的PCR擴增產物用3.0%的瓊脂糖凝膠電泳檢測,緩沖體系1×TAE溶液,150 V電壓電泳35 min,0.5%溴化乙錠(EB)染色10 min,蒸餾水漂洗后用VILBERLOURMAT凝膠成像系統掃描成像,并存入計算機。
1.4 統計方法
根據每個品種(系)三粒種子DNA檢測結果判斷該品種的LOX基因 TaLox-B1、 TaLox-B2、 TaLox-B3位點的等位變異類型,對PCR產物條帶清晰、單一并符合目標條帶大小的結果進行統計,若出現結果不一致的,需重新提取DNA并檢測,以3次檢測一致的結果為準。
2 結果與分析
2.1 新疆小麥LOX基因 TaLox-B1、 TaLox-B2和 TaLox-B3等位變異類型及分布
用互補顯性功能標記LOX16 、LOX18和共顯性功能標記 Lox-B23對123份新疆小麥品種(系)進行等位變異檢測(圖1和圖2),結果表明,除 TaLox-B1位點的 TaLox-B1b變異類型的材料有90份(73.2%),為優勢等位變異外, TaLox-B2和 TaLox-B3位點分別以 TaLox-B2a和 TaLox-B3a為優勢等位變異類型,分別為122份(99.2%)和95份(77.2%)(表2)。99份冬小麥材料在三個位點的變異類型的分布頻率也具有上述特點,具有各基因優勢等位變異 TaLox-B1b、 TaLox-B2a和 TaLox-B3a的材料分別有69份(69.7%)、99份(100%)和89份(89.9%)(表2)。24份春小麥材料中,除 TaLox-B2位點具有 TaLox-B2a的材料有23份(95.8%),為優勢等位變異外, TaLox-B1和 TaLox-B3位點均是以 TaLox-B1b和 TaLox-B3b為優勢等位變異,分別為21份(87.5%)和18份(75.0%)(表2)。

M:DL2000;1:冀麥24;2:碧螞6號;3:濟南4號;4:新春27;5:新春29;6:奎花2號;7:新冬17;8:新春17;9:新春35;10:新春37。
M:DL2000;1:Jimai 24;2:Bima 6;3:Jinan 4;4:Xinchun 27;5:Xinchun 29;6:Kuihua 2;7:Xindong 17;8:Xinchun 17;9:Xinchun 35;10:Xinchun 37.
圖1 LOX16(a)和 LOX18(b)標記檢測部分小麥品種 TaLox-B1位點的等位變異結果
Fig.1 Identification of TaLox-B1 alleles by functional markers LOX16 (a) and LOX18 (b)in part of wheat varieties
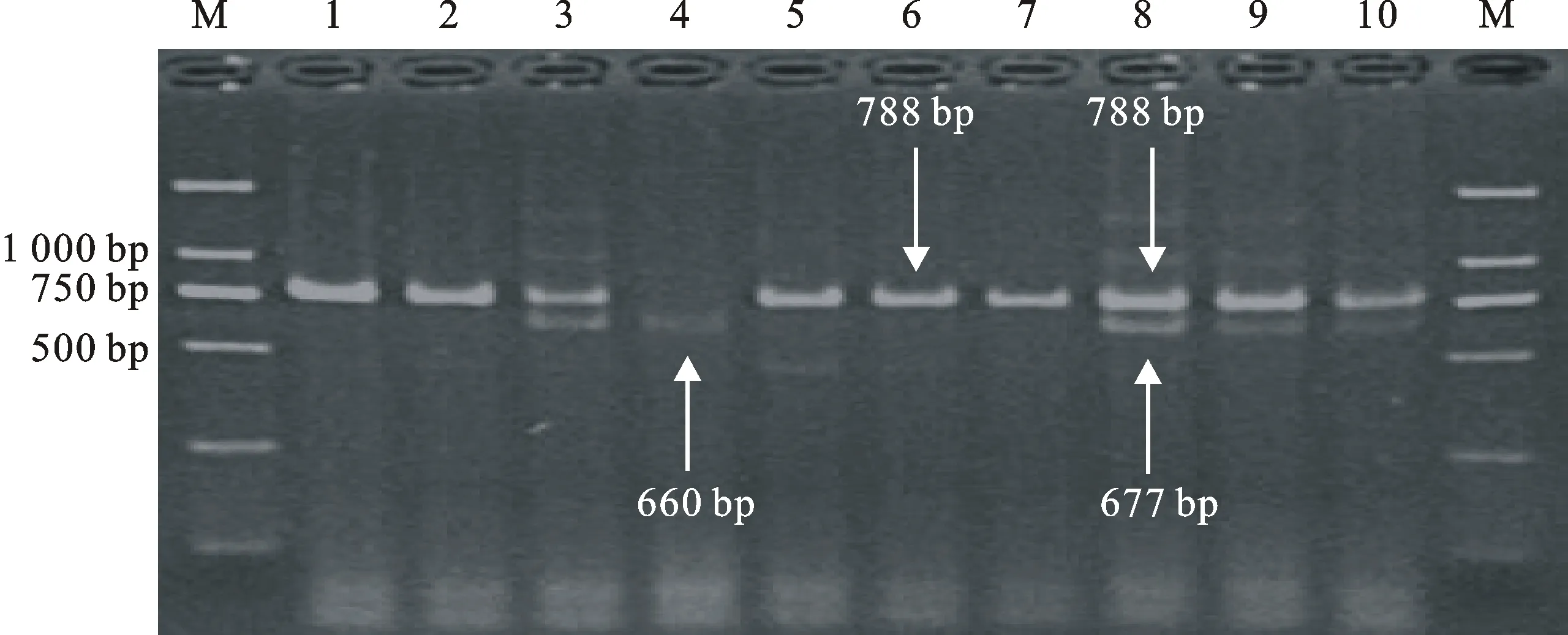
M:DL2000;1:新春14;2:紅直頭10;3:新春20;4:新春21;5:洛夫林18號;6:新春23;7:新春26;8:小白冬麥;9:新春28;10:新冬2號。
M:DL2000;1:Xinchun 14;2:Hongzhitou 10;3:Xinchun 20;4:Xinchun 21;5:Luofulin 18;6:Xinchun 23;7:Xinchun 26;8:Xiaobaidongmai;9:Xinchun 28;10:Xindong 2.
圖2 Lox-B23標記檢測部分小麥品種 TaLox-B2和 TaLox-B3位點的等位變異結果
Fig.2 Identification of TaLox-B2 and TaLox-B3 alleles by functional markers Lox-B23 in part of wheat varieties
表2 供試材料的 TaLox-B1、 TaLox-B2和 TaLox-B3 基因分布頻率
Table 2 Frequency of TaLox-B1, TaLox-B2 and TaLox-B3 in tested materials
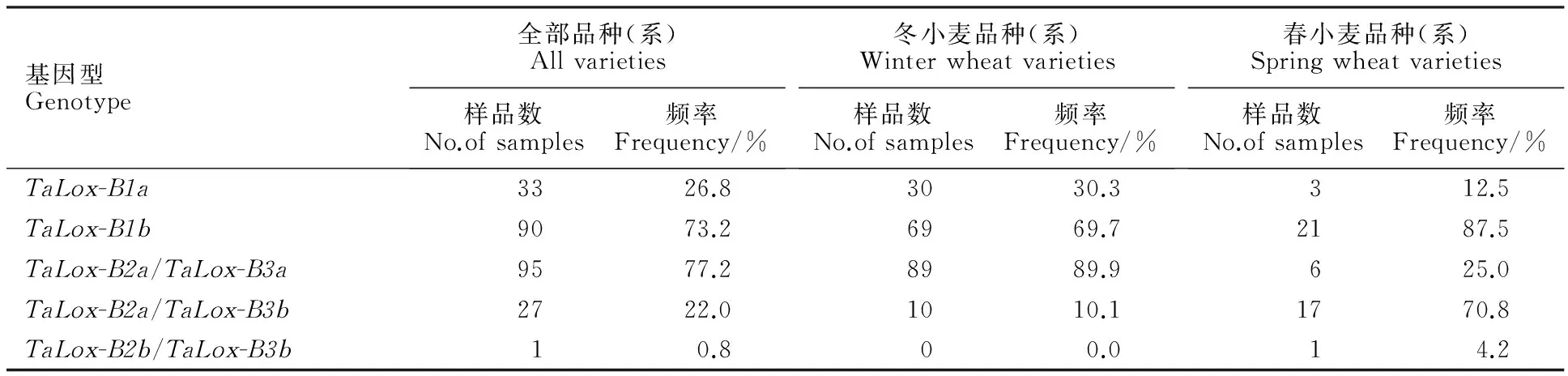
基因型Genotype全部品種(系)Allvarieties樣品數No.ofsamples頻率Frequency/%冬小麥品種(系)Winterwheatvarieties樣品數No.ofsamples頻率Frequency/%春小麥品種(系)Springwheatvarieties樣品數No.ofsamples頻率Frequency/%TaLox-B1a3326.83030.3312.5TaLox-B1b9073.26969.72187.5TaLox-B2a/TaLox-B3a9577.28989.9625.0TaLox-B2a/TaLox-B3b2722.01010.11770.8TaLox-B2b/TaLox-B3b10.800.014.2
2.2 不同類型新疆冬小麥材料的 TaLox-B1、 TaLox-B2和 TaLox-B3 等位變異類型及分布
對99份新疆冬小麥材料 TaLox-B1位點的檢測結果表明,17份地方品種中具有 TaLox-B1a的材料有1份(5.9%),具有 TaLox-B1b的材料有16份(94.1%);39份引進品種中具有 TaLox-B1a的材料有19份(48.7%),具有 TaLox-B1b的材料有20份(51.3%);43份自育品種中具有 TaLox-B1a的材料有10份(23.3%),具有 TaLox-B1b的材料有33份(76.7%)。說明新疆冬小麥地方品種中優異等位變異 TaLox-B1a的分布頻率遠低于 TaLox-B1b,但在地方品種、自育品種和引進品種中逐次升高。 對于 TaLox-B3位點而言,17份地方品種均具有 TaLox-B3a;39份引進品種具有 TaLox-B3a的材料有37份(94.9%),2份(5.1%)具有 TaLox-B3b;43份自育品種中具有 TaLox-B3a的材料有35份(81.4%),具有 TaLox-B3b的材料有8份(18.6%)。總的來看,新疆冬小麥地方品種中優異等位變異 TaLox-B3a的分布頻率遠高于 TaLox-B3b,并在地方品種、引進品種和自育品種中逐次降低(表3)。
表3 不同類型冬小麥品種(系)的 TaLox-B1、 TaLox-B2和 TaLox-B3 基因分布頻率
Table 3 Frequency of TaLox-B1, TaLox-B1 and TaLox-B1 in different types of winter wheat varieties

基因型Genotype全部品種(系)Allvarieties樣品數No.ofsamples頻率Frequency/%地方品種(系)Localvarietieies樣品數No.ofsamples頻率Frequency/%引進品種(系)Introducedvarieties樣品數No.ofsamples頻率Frequency/%自育品種(系)Bredvarieties樣品數No.ofsamples頻率Frequency/%TaLox-B1a3030.315.91948.71023.3TaLox-B1b6969.71694.12051.33376.7TaLox-B2a/TaLox-B3a8989.917100.03794.93581.4TaLox-B2a/TaLox-B3b1010.1000.025.1818.6
2.3 不同時期新疆春小麥材料的 TaLox-B1、 TaLox-B2和 TaLox-B3等位變異類型及分布
24份新疆春小麥材料 TaLox-B1位點的檢測結果表明,4份早期品種都具有 TaLox-B1b,20份晚期品種中有3份(15.0%)具有 TaLox-B1a,17份(85.0%)具有 TaLox-B1b;新疆春小麥早期品種未發現優異等位變異 TaLox-B1a,晚期品種雖然檢測到 TaLox-B1a,但其分布頻率也不高。對于 TaLox-B2位點而言,4份早期品種均具有 TaLox-B2a,20份晚期品種中19份(95.0%)材料具有 TaLox-B2a變異類型,只有1份(5.0%)材料具有 TaLox-B2b,表明新疆春小麥早期品種優異等位變異 TaLox-B2a的分布頻率高于晚期品種。對于 TaLox-B3位點而言,4份早期品種均具有 TaLox-B3a,20份晚期品種中具有 TaLox-B3a和 TaLox-B3b的材料分別有6份(30.0%)和14份(70.0%);新疆春小麥材料早期品種均為優異等位變異 TaLox-B3a,且其分布頻率遠高于晚期品種(表4)。
2.4 新疆小麥材料的 TaLox-B1、 TaLox-B2和 TaLox-B3等位變異基因組合及分布頻率
對123份新疆小麥材料 TaLox-B1、 TaLox-B2、 TaLox-B3等位變異類型的統計表明,具有最高LOX活性的基因型 TaLox-B1a/ TaLox-B2a/ TaLox-B3a或 TaLox-B1b/ TaLox-B2a/ TaLox-B3a以及具有中高LOX活性的基因型 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b的材料分別有95份(77.2%)和27份(22.0%);材料中未發現最低LOX活性的基因組合 TaLox-B1a/ TaLox-B2b/ TaLox-B3b,其中 TaLox-B1b/ TaLox-B2b/ TaLox-B3b是新發現的基因組合,在新疆材料中僅有1份(0.8%),其LOX活性尚未檢測。而99份新疆冬小麥材料中,具有 TaLox-B1a/ TaLox-B2a/ TaLox-B3a和 TaLox-B1b/ TaLox-B2a/ TaLox-B3a以及 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b基因組合的材料分別有89份(89.9%)和10份(10.1%)。24份新疆春小麥材料中,具有 TaLox-B1a/ TaLox-B2a/ TaLox-B3a和 TaLox-B1b/ TaLox-B2a/ TaLox-B3a以及 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b和 TaLox-B1b/ TaLox-B2b/ TaLox-B3b基因組合的材料分別有6份(25.0%)、17份(70.8%)和1份(4.2%)(表5)。總的來看,新疆冬小麥品種具有最高LOX活性的基因型頻率遠高于春小麥。8個冬小麥(石冬8號、新冬18號、新冬20號、新冬21號、新冬29號、新冬30號、新冬33號、新冬36號、伊農19、新冬7號)和2個春小麥自育品種(新春27號和新春29號)以及1個地方品種(庫車白冬麥)具有最高LOX活性基因組合 TaLox-B1a/ TaLox-B2a/ TaLox-B3a,上述11個材料可作為選育攜帶高LOX活性基因的優異種質材料。
表4 不同類型春小麥品種(系)的 TaLox-B1、 TaLox-B2和 TaLox-B3基因的分布頻率
Table 4 Frequency of TaLox-B1, TaLox-B2 and TaLox-B3 in different types of spring wheat varieties

基因型Genotype全部品種(系)Allvarieties樣品數No.ofsamples頻率Frequency/%早期品種(系)Earlyvarieties樣品數No.ofsamples頻率Frequency/%晚期品種(系)Latevarieties樣品數No.ofsamples頻率Frequency/%TaLox-B1a312.500315.0TaLox-B1b2187.54100.01785.0TaLox-B2a/TaLox-B3a625.04100.0630.0TaLox-B2a/TaLox-B3b1770.8001365.0TaLox-B2b/TaLox-B3b14.20015.0
表5 新疆小麥品種不同 TaLox-B等位基因的組合頻率
Table 5 Frequency of Xinjiang wheat cultivars with different TaLox-B allelic combinations

等位變異組合Alleliccombination全部品種(系)Allvarieties樣品數No.ofsamples頻率Frequency/%冬小麥品種(系)Winterwheatvarieties樣品數No.ofsamples頻率Frequency/%春小麥品種(系)Springwheatvarieties樣品數No.ofsamples頻率Frequency/%TaLox-B1a/TaLox-B2a/TaLox-B3a2923.62727.328.3TaLox-B1b/TaLox-B2a/TaLox-B3a6653.76262.6416.7TaLox-B1b/TaLox-B2a/TaLox-B3b2419.588.11666.7TaLox-B1a/TaLox-B2a/TaLox-B3b32.422.014.2TaLox-B1b/TaLox-B2b/TaLox-B3b10.80014.2
3 討 論
面粉及面制品顏色是小麥品質分級的重要指標[18-19]。長期以來我國消費者有喜歡高白度面食的習慣[20 ]。LOX通過與小麥面粉中的類胡蘿卜素發生偶聯氧化作用降低黃色素含量進而使小麥面粉變白[4],有望從根本上解決使用面粉增白劑對人體產生的潛在危害[21]。選育高LOX活性的小麥品種對提高面粉白度具有重要的作用。分子標記技術具有便捷、準確、穩定的特點,在小麥抗病性檢測、品質分析以及與小麥各種農藝性狀相關因素的檢測方面發揮了巨大作用[22-23]。本研究所用的LOX活性相關功能標記LOX16、LOX18在Geng等[12]和相吉山等[14]研究中發現能有效應用于大群體小麥品種資源的檢測以及LOX活性的判斷,在育種實踐中具有較高的應用價值;而Zhang等[16]在研究中也證明了Lox-B23的可靠性。利用LOX基因3個位點的功能標記能更有效地對高LOX活性材料進行選育。
相吉山等[14]利用 TaLox-B1位點的LOX16和LOX18功能標記檢測195份新疆小麥資源,結果表明 TaLox-B1a基因的頻率24.1%。許多研究利用LOX16、LOX18功能標記對不同區域小麥資源的檢測結果均表明 TaLox-B1a基因型頻率所占的比例較低[13,16,24-26]。本研究利用3對功能標記LOX16、LOX18和Lox-B23對123份新疆小麥資源進行檢測,結果表明各位點等位變異類型的分布與前人研究結果基本一致。新疆冬小麥資源中的地方品種、自育品種和引進品種 TaLox-B1a基因型分布頻率依次升高,其中地方品種(5.9%)和自育品種(23.3%)低于全國的分布頻率(26.5%),但引進品種(48.7%)高于全國的分布頻率,這可能與國內對LOX活性重視和研究起步較晚有關。自育品種 TaLox-B1a基因型分布頻率高于地方品種但低于引進品種,這表明新疆小麥受地方品種的影響,使 TaLox-B1a基因型分布頻率比較低,但隨著引進品種不斷引入新疆以及育種工作的進行,使高LOX活性 TaLox-B1a基因型頻率升高,但新疆冬小麥 TaLox-B1a基因型分布頻率總體水平仍然較低(30.3%),與國外品種存在差距。同時也表明,引種是小麥品質遺傳改良的有效途徑之一,應加大對國外優良品種的引進和自育品種的選育,進而縮小與國外優良品種的差距。
Zhang等[16]利用Lox-B23標記檢測136份中國小麥資源,結果表明, TaLox-B1a/ TaLox-B2a/ TaLox-B3a或 TaLox-B1b/ TaLox-B2a/ TaLox- B3a基因組合的頻率為62.5%。本研究利用Lox-B23功能標記對123份新疆小麥資源進行檢測,結果最高LOX活性的基因組合頻率為77.2%,高于Zhang等[16]的研究結果。在新疆冬小麥品種中最高LOX活性的基因組合頻率高達89.9%,而在新疆春小麥品種中只有25.0%。雖然新疆小麥品種最高LOX活性的基因組合頻率受春小麥品種的影響有所降低,但仍然高于全國水平(62.5%);新疆冬小麥品種最高LOX活性的基因組合頻率高于全國水平,同時高于新疆總體水平,說明新疆冬小麥品種資源是培育高LOX活性品種的優異基因源。本研究還檢測出新春21號品種的基因型是 TaLox-B1b/ TaLox-B2b/ TaLox-B3b,雖其LOX活性尚未檢測,但其 TaLox-B1、 TaLox-B2、 TaLox-B3位點基因都與高LOX活性負相關,且低LOX活性材料具有更耐儲藏的特性[1,16],因此新春21號小麥品種可能具有較長的儲藏期。
本研究將3個LOX主效基因位點結合起來進行選擇,很大程度上彌補了單一位點選擇的片面和不足,將能更有效的選出高LOX活性材料。目前,對新疆小麥LOX活性的研究都集中在分子標記檢測上,并用基因型來選擇高LOX活性材料,還沒有對新疆小麥材料LOX活性進行測定研究。同時,本研究所用春小麥樣本數有限,均為自育品種,分類時未能與冬小麥材料采用同一標準,致使春小麥是按不同時期進行分類,雖然也能有效反映新疆小麥材料中不同等位變異類型分布的演化規律及其成因,但未能反映冬春麥之間演化規律的異同。下一步我們的工作是進一步擴大新疆春小麥材料數量,開展新疆小麥材料LOX活性的表型測定,并通過表型結合基因型對新疆小麥材料的LOX活性進行綜合評價,這將提升高LOX活性選育效率,為小麥品質改良提供優異基因源和基礎材料。
[1] 耿洪偉.小麥脂肪氧化酶(LOX)活性QTL定位與功能標記開發[D].烏魯木齊:新疆農業大學,2010:9.
GENG H W.Mapping QTLs for lipoxygenase activity and development of functional markers for LOX gene in common wheat [D].Urumqi:Xinjiang Agricultural University,2010:9.
[2] 胡瑞波,田紀春.小麥主要品質性狀與面粉色澤的關系[J].麥類作物學報,2006,26(3):96.
HU R B,TIAN J C.Relationship between main quality characteristics and wheat flour color [J].JournalofTriticeaeCrops,2006,26(3):96.
[3] 汪 帆,鄭文寅,黃建華.20個小麥品種(系)籽粒LOX活性和類胡蘿卜素含量及全麥粉色澤的研究[J].麥類作物學報,2012,32(1):68.
WANG F,ZHENG W Y,HUANG J H,etal.Lipoxygenase activity,carotenoids content and wholemill colors in grains of twenty wheat varieties [J].JournalofTriticeaeCrops,2012,32 (1):68.
[4] 鄭文寅,汪 帆,司紅起.普通小麥籽粒LOX、PPO活性和類胡蘿卜素含量變異及對全麥粉色澤的影響[J].中國農業科學,2013,46(6):1087.
ZHENG W Y,WANG F,SI H Q,etal.Variations of LOX and PPO activities and carotenoid conten as well as their influence on whole flour color in common wheat [J].ScientiaAgricultureSinica,2003,46(4):1087.
[5] SHEWRY P R,TATHAM A S,BARRO F,etal.Biotechnology of breadmaking-unravelling and manipulating the multi-protein gluten complex [J].Biotechnology,1995,13(11):1185.
[6] BORRELLI G M,TROCCOLI A,FONZO N,etal.Durum wheat lipoxygenase activity and other quality parameters that affect pasta color [J].CerealChemistry,1999,76(3):335.
[7] 王 慧,鄭文寅,樊 宏,等.不同小麥品種籽粒中LOX活性及基因型和環境互作分析[J].中國糧油學報,2011,26(1):11.
WANG H,ZHENG W Y,FAN H,etal.Lipoxygenase activity and its genotype and environment interactions for different wheat varieties [J].JournaloftheChineseCerealsandOilsAssociation,2011,26(1):11.
[8] PSHENICHNIKOVA T A,OSIPOVA S V,PERMYAKOVA M D,etal.Mapping of quantitative trait loci (QTL) associated with activity of disulfide reductase and lipoxygenase in grain of bread wheatTriticumaestivumL. [J].RussianJournalofGenetics,2008,44(5):567.
[9] FENG B,DONG Z Y,XU Z B,etal.Molecular analysis of lipoxygenase (LOX) genes in common wheat and phylogenetic investigation of LOX proteins from model and crop plants [J].JournalofCerealScience,2010,52(3):387.
[10] HART G E,LANGSTON P J.Chromosomal location and evolution of isozyme structural genes in hexaploid wheat [J].Heredity,1977,39(2):263.
[11] GENG H W,ZHANG Y,HE Z H,etal.Molecular markers for tracking variation in lipoxygenase activity in wheat breeding [J].MolecularBreeding,2011,28(1):117.
[12] GENG H W,XIA X C,ZHANG L P,etal.Development of functional markers for a lipoxygenase gene TaLox-B1 on chromosome 4BS in common wheat [J].CropScience,2012,52(2):568.
[13] 張鈺玉,王曉龍,張曉科,等.陜西小麥品種(系)脂肪氧化酶活性基因的遺傳多態性分析[J].麥類作物學報,2012,32(4):616.
ZHANG Y Y,WANG X L,ZHANG X K,etal.Genetic polymorphism of lipoxygenase genes in Shaanxi wheat cultivars and lines [J].JournalofTriticeaeCrops,2012,32(4):616.
[14] 相吉山,穆培源,桑 偉,等.新疆小麥品種資源脂肪氧化酶活性基因 TaLox-B1的分布特征研究[J].麥類作物學報,2013,33(2):279.
XIANG J S,MU P Y,SANG W,etal.Distribution characteristics of lipoxygenase gene TaLox-B1 in wheat varieties from Xinjiang [J].JournalofTriticeaeCrops,2013,33(2):279.
[15] 楊淑萍,張宏紀,劉文林,等.黑龍江春小麥脂肪氧化酶活性基因多態性分析[J].核農學報,2014,28(11):1972.
YANG S P,ZHANG H J,LIU W L,etal.Polymorphism of lipoxygenase genes in Heilongjiang spring wheat cultivars[J].JournalofNuclearAgriculturalSciences,2014,28(11):1972.
[16] ZHANG F Y,CHEN F,WU P,etal.Molecular characterizeation of lipoxygenase genes on chromosome 4BS in Chinese bread wheat (TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2015,128(8):1467.
[17] LAGUDAH E S,APPLES R,MCNEIL D.The Nor-D3 locus ofTriticumtauschii:natural variation and genetic linkage to markers in chromosome 5 [J].Genome,1991,34(3):387.
[18] 何中虎,晏月明,莊巧生,等.中國小麥品種品質評價體系建立與分子改良技術研究[J].中國農業科學,2006,39(6):1091.
HE Z H,YAN Y M,ZHUANG Q S,etal.Establishment of quality evaluation system and utilization of molecular methods for the improvement of Chinese wheat quality [J].ScientiaAgricultureSinica,2006,39(6):1091.
[19] 胡瑞波,田紀春,鄧志英,等.中國白鹽面條色澤影響因素的研究[J].作物學報,2006,32(9):1338.
HU R B,TIAN J C,DENG Z Y,etal.Factors related to Chinese white salted noodle color [J].ActaAgronomicaSinica,2006,32(9):1338.
[20] 張 曉,田紀春.若干高白度小麥的色澤優勢及形成因素分析[J].中國農業科學,2008,41(2):347.
ZHANG X,TIAN J C.The color advantage of Chinese wheat with high whiteness and analysis of factors affecting color formation [J].ScientiaAgriculturaSinica,2008,41(2):347.
[21] 田紀春.禁用面粉增白劑后保證中國傳統面食品色澤品質的對策[J].麥類作物學報,2007,27(2):364.
TIAN J C.Strategy of keeping quality of wheat flour and food color when the flour bleacher inhibited [J].JournalofTriticeaeCrops,2007,27(2):364.
[22] 武玉華.分子標記輔助選擇在小麥抗病和品質遺傳育種中的應用[J].麥類作物學報,2007,27(5):928.
WU Y H.Application of marker-assisted selection in disease-resistance and high-quality wheat breeding [J].JournalofTriticeaeCrops,2007,27(5):928.
[23] 王長有,吉萬全,薛秀莊.分子標記技術在小麥遺傳育種中的應用現狀[J].麥類作物學報,2000,20(4):75.
WANG C Y,JI W Q,XUE X Z.The application of molecular marker techniques in the wheat genetic and breeding research [J].JournalofTriticeaeCrops,2000,20(4):75.
[24] 吳培培,宋 雙,張福彥,等.黃淮麥區部分小麥種質脂肪氧化酶活性分析及等位基因檢測[J].中國農業科學,2015,48(2):207.
WU P P,SONG S,ZHANG F Y,etal.The allelic variation of lipoxygenase genes in bread wheat cultivars from the Yellow and Huai wheat areas of China [J].ScientiaAgricultureSinica,2015,48(2):207.
[25] 楊淑萍,張宏紀,劉文林,等.黑龍江春小麥脂肪氧化酶活性基因多態性分析[J].核農學報,2014(11):1972.
YANG S P,ZHANG H J,LIU W L,etal.Polymorphism of lipoxygenase genes in Heilongjiang spring wheat cultivars [J].JournalofNuclearAgriculturalScience,2014,28(11):1972.
[26] 楊 杰,亢 玲,王憲國,等.寧夏小麥脂肪氧化酶基因的檢測與分布[J].麥類作物學報,2015,35(7):896.
YANG J,KANG L,WANG X G,etal.Detections and analysis of lipoxygenase genes in Ningxia wheat varieties [J].JournalofTriticeaeCrops,2015,35(7):896.
Molecular Identification and Distribution of TaLox-B1, TaLox-B2 and TaLox-B3 Loci in Xinjiang Wheat Cultivars
BAI Lu1,YU Feixiong2,YASENG Ruxianguli3,Lü Wenjuan2,REN Yi2,XIE Lei2,GENG Hongwei2
(1.College of Science and Technology,Xinjiang Agricultural University,Urumqi,Xinjiang 830052,China; 2.Key Laboratory of Agricultural Biological Technology/College of Agronomy,Xinjiang Agricultural University,Urumqi,Xinjiang 830052,China;3.Bayingol Mongolian Autonomous Prefecture of Agriculture,Korla,Xinjiang 841000,China)
The color of the wheat flour is an important index of wheat quality traits. This study used the functional markers LOX16,LOX18 and Lox-B23,based on the development of wheat 4BS chromosome,and molecular markers were detected by TaLox-B1, TaLox-B2 and TaLox-B3 in 123 wheat varieties in Xinjiang. The results showed that,there were 33(26.8%)varieties with TaLox-B1a allele exhibited high Lox activity,90(73.2%)varieties with TaLox-B1b allele showed low Lox activity; 122(99.2%)varieties with TaLox-B2a allele exhibited high Lox activity,only 1(0.8%)varieties with TaLox-B2b allele showed low Lox activity; 95(77.2%)varieties with TaLox-B2a allele exhibited high LOX activity,28(22.8%)varieties with TaLox-B2b allele showed low Lox activity. In different types of winter wheat varieties,the frequency distribution of TaLox-B1a was in order of introduced varieties>self-cultivated varieties >local varieties. In different types of spring wheat varieties,the TaLox-B1a exists only in the late varieties,and the distribution frequency of both TaLox-B2a and TaLox-B3a was in order of early varieties (100%) >late varieties.The analysis of 3 loci genotype combinations indicated that there are 5 kinds of genetic combinations in Xinjiang wheat varieties,which were both TaLox-B1a/ TaLox-B2a/ TaLox-B3a and TaLox-B1b/ TaLox-B2a/ TaLox-B3a combinations exhibited the highest LOX activity,and were both TaLox-B1b/ TaLox-B2a/ TaLox-B3b and TaLox-B1a/ TaLox-B2a/ TaLox-B3b combinations exhibited high LOX activity,and TaLox-B1b/ TaLox-B2b/ TaLox-B3b showed the lowest LOX activity,and the distribution frequency was 23.6%,53.7%,19.5%,2.4% and 0.8%,respectively. The genotype frequency of high LOX activity in wheat cultivars in Xinjiang was higher than the national average. In this study,3 loci functional markers were combined more effectively applied to wheat quality molecular marker assisted selection.
Xinjiang wheat; Lipoxygenase; Function marker; Variation combination
時間:2016-11-04
2016-05-16
2016-06-05
國家自然科學基金項目(31260327);新疆自治區級大學生創新項目(20150016);2014年度留學人員科技活動擇優資助項目(耿洪偉)
E-mail:bailu32@163.com
耿洪偉(E-mail:hw-geng@163.com)
S512.1;S330
A
1009-1041(2016)11-1456-08
網絡出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20161104.0924.014.html
猜你喜歡
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
新作文·高中版(2021年4期)2021-07-14 11:55:06
環球人物(2021年8期)2021-05-06 08:00:43
音樂天地(音樂創作版)(2020年12期)2021-01-11 01:13:40
音樂天地(音樂創作版)(2020年7期)2020-09-11 06:17:18
華人時刊(2018年15期)2018-11-10 03:25:32
人民中國(日文版)(2015年10期)2015-04-16 03:53:48
絲綢之路(2014年9期)2015-01-22 04:24:46
新疆人文地理(2009年7期)2009-09-29 09:56:14
