能源橡膠草GGPPS基因啟動子的克隆及瞬時表達研究
2017-01-03 10:50:06李永梅馮玉杰曹新文趙李靖祝建波閆潔
草業學報 2016年12期
李永梅,馮玉杰,曹新文,趙李靖,祝建波,閆潔
(石河子大學生命科學學院,新疆 石河子 832000)
?
能源橡膠草GGPPS基因啟動子的克隆及瞬時表達研究
李永梅,馮玉杰,曹新文,趙李靖,祝建波,閆潔*
(石河子大學生命科學學院,新疆 石河子 832000)
以多年生宿根型草本植物橡膠草為材料,根據已獲得的橡膠草牻牛兒基牻牛兒基焦磷酸合酶(geranylgeranyl pyrophosphate synthase,GGPPS)基因序列設計引物,采用TAIL-PCR擴增到GGPPS基因5′上游大小為1131 bp的序列。采用PlantCare和PLACE軟件分析,表明該序列不僅具有啟動子的基本元件,同時還有與多個器官特異表達元件以及與脅迫相關的順式作用元件,命名為pTkGGPPS(GenBank:KT901796)。將該序列代替pCAMBIA1304質粒上的CaMV 35S啟動子序列,以GFP基因作為報告基因,構建pCAMBIA1304-pTkGGPPS-GFP植物表達載體,利用農桿菌介導法轉化洋蔥表皮細胞,結果表明,該啟動子能夠驅動GFP基因表達,具有一定的活性。TkGGPPS基因啟動子克隆及瞬時表達為橡膠草中橡膠合成及組織特異性研究奠定了基礎。
橡膠草;GGPPS基因;TAIL-PCR;啟動子;序列分析;瞬時表達
啟動子是位于結構基因5′端上游的DNA序列,能夠活化RNA聚合酶,使之與模板DNA準確結合并具有轉錄起始特異性。啟動子(promoters)就像“開關”,決定轉錄的方向和效率,控制基因表達的起始時間、空間和表達程度。目前,大多數轉基因植物所用的啟動子是組成型啟動子(如煙草花葉病毒35S啟動子)。組成型啟動子是基因工程中應用最早,且應用最為廣泛的一類啟動子。它能夠在組織中非特異性和高效表達,外源基因的高效表達,能夠滿足人類的需求;但是,外源基因的非特異性表達和異源基因的高效表達,打破植物原有的代謝平衡,影響植物的正常生長[1]。為了減少外源基因在組織中非特異性和高效表達所帶來的不利影響,尋求組織特異性啟動子就顯得尤為重要。
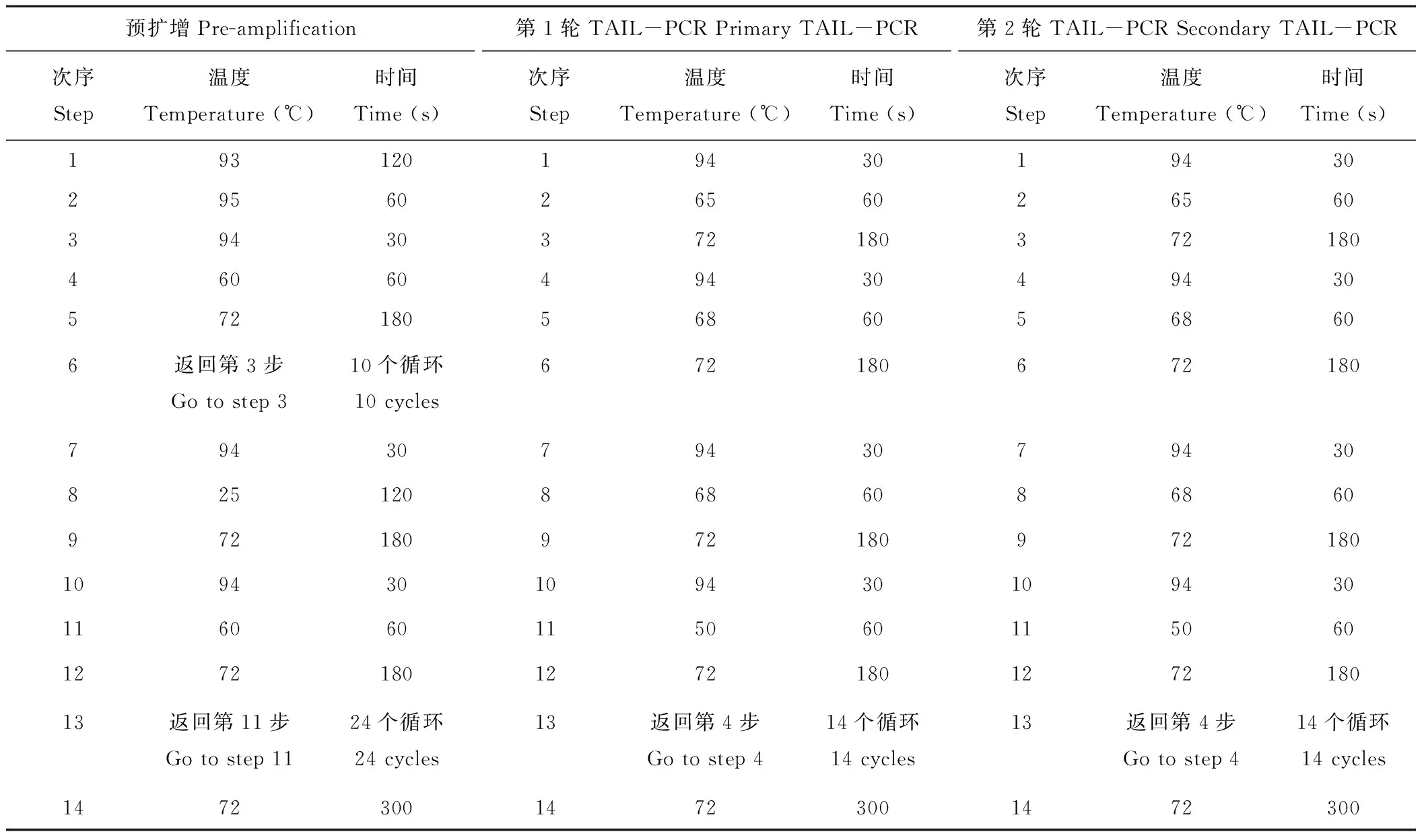
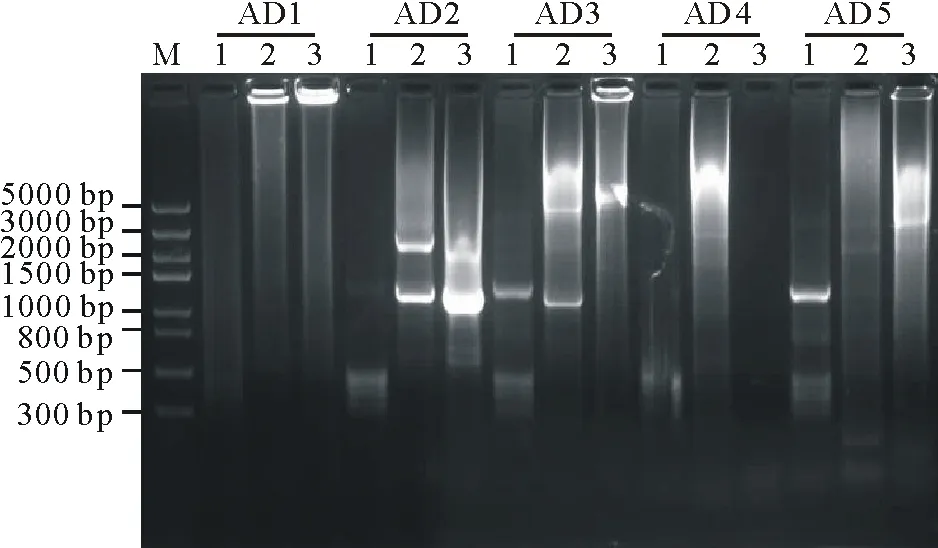
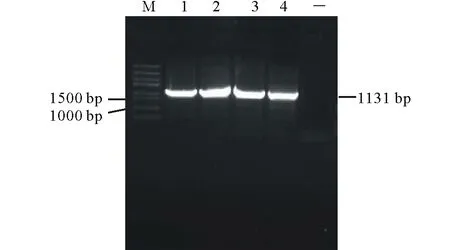
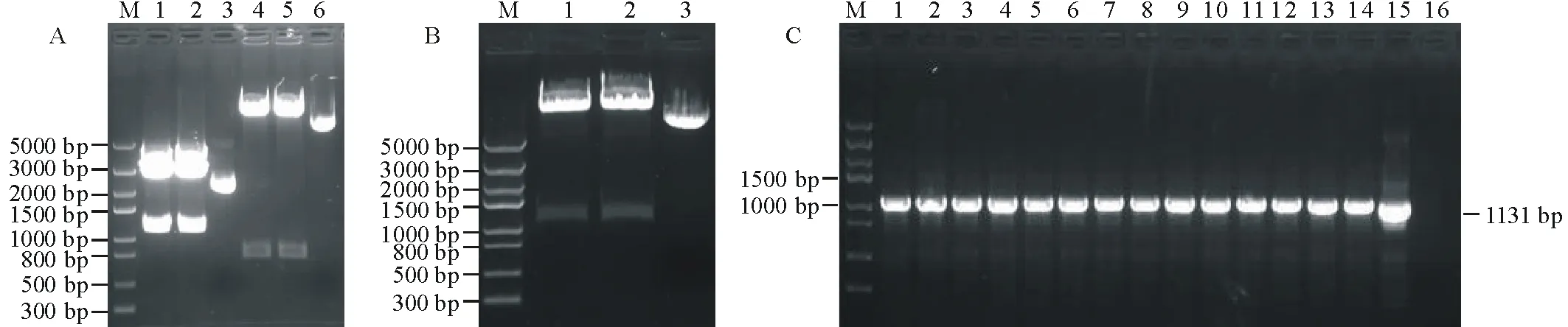
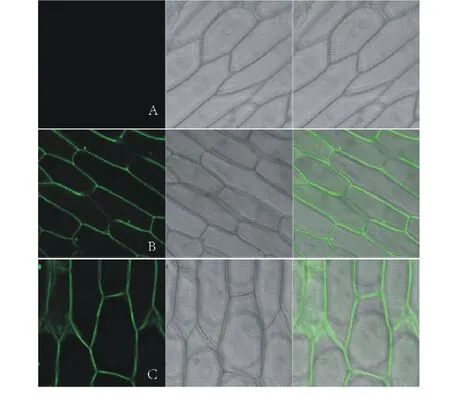
橡膠草(Taraxacumkok-saghyz)是一種多年生的產膠草本植物,作為產膠代謝機理研究的生物反應器,對橡膠草產膠相關功能基因的研究尤為重要。牻牛兒基牻牛兒基焦磷酸合酶是產膠代謝途徑的一個上游基因,它能夠催化烯丙基焦磷酸合成GGPP。橡膠分子合成起始是一個十分緩慢的過程,是橡膠生物合成的關鍵限速步驟;增加橡膠產量的遺傳控制戰略應集中在橡膠分子合成的起始上[2]。有研究者報道,在橡膠體外合成試驗中,二甲基烯丙基焦磷酸(dimethylallyl diphosphate,DMAPP)、香葉基焦磷酸(geranyl diphosphate,GPP)、法呢基焦磷(farnesyl diphosphate,FPP)和牻牛兒基牻牛兒基焦磷酸(geranylgeranyl pyrophosphate,GGPP)都可作為橡膠分子合成的前體物質且各起始物都以一個恒定的速度啟動橡膠的合成。但是,隨著起始物碳鏈的增長,橡膠合成速度也增加(起始物鏈長效應),即DMAPP(C5) 因此,本研究通過熱不對稱性PCR(thermal asymmetric interlaced PCR,TAIL-PCR)技術,克隆橡膠草中GGPPS基因的特異性啟動子,并對其進行活性研究;根據啟動子所含有的順式調控元件,預測該啟動子的功能,為提高橡膠草橡膠合成奠定理論基礎。 1.1 材料 2014年9月,橡膠草采自新疆石河子市北泉鎮附近。 1.2 方法 1.2.1 橡膠草TkGGPPS基因啟動子的克隆 根據已克隆的橡膠草TkGGPPS基因序列設計3個嵌套的特異性引物:sp1、sp2、sp3,5個短的簡并AD引物:AD1、AD2、AD3、AD4和AD5(表1)。用CTAB法提取橡膠草葉片基因組DNA,3個特異引物和5個短的AD引物組合進行TAIL-PCR擴增。反應體系:模板(基因組DNA)1 μL,2.5 mmol/L dNTPs 2 μL,引物各0.5 μL,10×LA-Taq buffer 2.5 μL,LA-Taq DNA polymerase(TaKaRa公司)0.5 μL,用ddH2O補足至25 μL。第2輪和第3輪PCR分別以上一輪PCR的產物稀釋100倍作為模板,反應體系同上。擴增程序參考文獻[5-9],具體擴增程序見表1。用1.0%的瓊脂糖凝膠電泳檢測3輪PCR擴增產物,用瓊脂糖凝膠DNA回收試劑盒(Tiangen公司)回收第3輪PCR中單一條帶,連接到pMD19-T載體(TaKaRa公司)上,連接產物轉化EscherichiacoliTop10感受態細胞,陽性單克隆交由北京華大基因股份有限公司進行測序。 根據TAIL-PCR所得序列設計特異引物:pTkGGPPS-F和pTkGGPPS-R。以橡膠草基因組DNA為模板,以pTkGGPPS-F和pTkGGPPS-R為上下游引物,擴增橡膠草TkGGPPS基因啟動子序列,擴增程序為94 ℃ 5 min;(94 ℃ 30 s,57 ℃ 30 s,72 ℃ 90 s)30個循環;72 ℃ 10 min。用1.0%的瓊脂糖凝膠電泳檢測,用DNA瓊脂糖凝膠回收試劑盒回收PCR產物。將純化產物與pMD19-T載體連接,連接產物轉化E.coliTop10感受態細胞,將陽性單克隆進行測序(表2)。 1.2.2 pCAMBIA1304-pTkGGPPS-GFP植物表達載體構建 用EcoRⅠ與NcoⅠ(TaKaRa公司)雙酶切pMD19-Tector-pTkGGPPS重組質粒與pCAMBIA1304質粒,分別回收小片段和大片段。將pTkGGPPS小片段與pCAMBIA1304載體大片段用T4-DNA連接酶(TaKaRa公司),16 ℃連接過夜,將重組質粒轉化E.coliTop10感受態細胞,經卡那霉素(Kan)篩選,進行PCR和雙酶切鑒定,經鑒定正確的陽性重組子,用電擊轉化法轉入根癌農桿菌GV3101,獲得陽性轉化子。命名為pCAMBIA1304-pTkGGPPS-GFP/GV3101。 表1 TAIL-PCR的擴增程序 Table 1 Amplification program of TAIL-PCR 預擴增Pre-amplification次序Step溫度Temperature(℃)時間Time(s)第1輪TAIL-PCRPrimaryTAIL-PCR次序Step溫度Temperature(℃)時間Time(s)第2輪TAIL-PCRSecondaryTAIL-PCR次序Step溫度Temperature(℃)時間Time(s)19312019430194302956026560265603943037218037218046060494304943057218056860568606返回第3步Gotostep310個循環10cycles672180672180794307943079430825120868608686097218097218097218010943010943010943011606011506011506012721801272180127218013返回第11步Gotostep1124個循環24cycles13返回第4步Gotostep414個循環14cycles13返回第4步Gotostep414個循環14cycles147230014723001472300 1.2.3 根癌農桿菌介導的方法轉化洋蔥表皮細胞 根癌農桿菌介導轉化洋蔥表皮細胞的方法,參照文獻[10-12]。在超凈工作臺中,將洋蔥表皮用手術刀劃成1 cm×1 cm的方塊,用尖頭鑷子從一角輕輕撕下,于MS培養基中預培養24 h,農桿菌28 ℃振蕩培養至OD260≈0.6,收集菌液,用等體積MS液體培養基重懸,將洋蔥表皮在重懸液中侵染30 min,用濾紙吸去多余的菌液后,重新接種到鋪有一層濾紙的培養皿中,共培養24 h后,激光共聚焦顯微鏡檢測熒光信號。 2.1 TAIL-PCR結果 經電泳檢測3輪PCR產物,AD2簡并引物和3條特異引物組合在第3輪PCR產物中有清晰的擴增條帶(圖1),經測序,該片段長1185 bp。 表2 PCR擴增使用的引物 Table 2 Primers used in PCR amplification 引物Primers引物序列Sequences(5'→3')sp15'-GATTTCCCCCAGAGTTTCCTGCTC-CTCCTTCGTA-3'sp25'-GGGTTTACTGCATTTCATGGGT-GTTTTGCAGATCA-3'sp35'-TCCGTTGAATATGGAGCATGTTT-GAACCCAAGAAT-3'AD15'-NTCGASTWTSGWGTT-3'AD25'-NGTCGASWGANAWGAA-3'AD35'-WGTGNAGWANCANAGA-3'AD45'-TGWGNAGWANCASAGA-3'AD55'-AGWGNAGWANCAWAGG-3'pTkGGPPS-F5'-TGGTCGAGTGAGATGAACC-3'pTkGGPPS-R5'-ATGTTTATCCAGGAAGATA-3' S: CG; W: AT; B: GTC; D: GAT; V: ACG; N: ACGT. 2.2 橡膠草TkGGPPS基因啟動子序列的克隆及分析以橡膠草基因組DNA為模板,以TAIL-PCR的測序結果設計特異引物pTkGGPPS-F和pTkGGPPS-R,進行PCR擴增,電泳檢測發現單一的特異條帶(圖2),與推測的理論值相符。測序發現該產物為橡膠草TkGGPPS基因的5′側翼序列,全長為1131 bp。命名為pTkGGPPS(GenBank登錄號:KT901796)。 圖1 TAIL-PCR擴增產物的電泳分析Fig.1 Analysis of TAIL-PCR products byagarose electrophoresisM: Trans5K DNA Marker(TaRaKa公司); 1: 第1輪PCR產物; 2: 第2輪PCR產物; 3: 第3輪PCR產物; AD1、AD2、AD3、AD4和AD5: 在TAIL-PCR中, 與特異引物組合的5個簡并引物。M: Trans5K DNA Marker; 1: Products of primary PCR; 2: Products of the secondary PCR; 3: Products of the tertiary PCR; AD1, AD2, AD3, AD4 and AD5: Five arbitrary degenerate primers in the TAIL-PCR. 圖2 橡膠草pTkGGPPS啟動子序列的電泳分析Fig.2 Analysis of TkGGPPS promoter on agarose electrophoresis M: Trans5K DNA Marker; 1-4: TkGGPPS啟動子的PCR產物PCR Product of TkGGPPS promoter; -: 陰性Negative. 2.3 橡膠草TkGGPPS基因啟動子序列的生物信息學分析 經啟動子分析軟件PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)和PLACE(https://sogo.dna.affrc.go.jp/cgi-bin/sogo.cgi?lang=en&pj=640&action=page&page=newplace)分析擴增得到pTkGGPPS啟動子序列(圖3)。分析表明,TkGGPPS基因啟動子序列不僅具有植物啟動子基本的順式作用元件(CAAT-box和TATA-box),還有與光、植物激素、逆境脅迫等相關的順式作用元件,比如I-box(光調節響應的順式作用元件(cis-acting elements involved in light-regulated responsiveness)、GARE(赤霉素應答相關的元件GA-responsive element)、GT-1 box(病原菌和鹽脅迫響應的順式作用元件cis-acting elements Plays a role in pathogen- and salt- induced responsiveness)、W-box(水楊酸誘導響應的WRKY結合位點WRKY binding wite involved in salicylic acid-induced responsiveness)、ACGT-motif(黃化誘導相關的順式作用元件cis-acting elements involved in etiolation-induced responsiveness)和LTRE(低溫應答元件low temperature responsive element)等。各個元件所具有的生物學功能如表3所示。 2.4 pCAMBIA1304-pTkGGPPS-GFP植物表達載體構建 用EcoRⅠ與NcoⅠ雙酶切pMD19-Tector-pTkGGPPS重組質粒和pCAMBIA1304質粒,結果如圖4A所示。分別回收目的小片段和載體大片段,將pTkGGPPS小片段與pCAMBIA1304載體大片段用T4-DNA連接酶,16 ℃,連接2 d,將重組質粒轉化E.coliTop10感受態細胞,涂布到含Kan抗性的LB篩選平板上,過夜培養。經PCR檢測和雙酶切雙重鑒定,結果如圖4B所示。用EcoRⅠ與NcoⅠ雙酶切,有一個約1131 bp的目的片段和一個大片段,說明 pCAMBIA1304-pTkGGPPS-GFP載體構建成功。將鑒定成功的重組質粒,轉化根癌農桿菌GV3101,PCR檢測結果如圖4C所示。 2.5 根癌農桿菌介導的洋蔥表皮細胞的瞬時轉化 將經PCR鑒定成功的陽性單克隆,利用根癌農桿菌介導的方法,對洋蔥表皮細胞進行瞬時轉化(圖5B),經質量分數為40%蔗糖溶液浸泡30 min發生質壁分離(圖5C)。結果表明,TkGGPPS基因的啟動子能夠驅動GFP基因的表達,表明該啟動子具有活性。 圖3 橡膠草GGPPS啟動子序列Fig.3 The promoter sequence of GGPPS in T. kok-saghyz 表3TkGGPPS啟動子順式作用元件的生物學功能分析 Table 3 Biological function analysis of cis-acting element ofTkGGPPSpromoter 調控元件Regulatoryelement序列Sequence生物學功能BiologicalfunctionLIPsGAGTGAG光誘導的根特異性順式作用元件。Cis-regulatoryelementinvolvedinlight-inducedroot-specificresponsiveness.TATA-boxTATA據轉錄起始位點30個堿基的啟動子中心元件。Corepromoterelementaround-30oftranscriptionstart.CCA1-BSAAMAATCTMYB相關的轉錄因子的結合位點。Bindingsitewithmyb-relatedtranscriptionfactor.W-boxTTGAC水楊酸誘導響應的WRKY結合位點。WRKYbindingwiteinvolvedinsalicylicacid(SA)-inducedresponsiveness.GATAboxGATAASF-2結合位點。ASF-2bindingsite.CACTmotifYACT羧化反應響應的順式調控元件。Cis-regulatoryelementinvolvedincarboxylaseresponsiveness.CArGmotifCWWWWWWWWGA/T豐富區。AlongerA/T-richcore.OSEmotifCTCTT組織特異性元件。Organ-specificelements.I-boxGATAA光調節響應的順式作用元件。Cis-actingelementsinvolvedinlight-regulatedresponsiveness.MYB1ATWAACCA脫水響應相關的MYB識別位點。MYBrecognitionsitefoundinthedehydration-responsive.CAAT-box1CAAT啟動子和增強子普遍存在的順式作用元件。Commoncis-actingelementsinpromoterandenhancerregion.ARR1-BSNGATT細胞色素調節響應的ARR1結合元件。ARR1-bindingelementinvolvedinthecytokinin-regulatedresponsiveness.GT-1motifGRWAAW光調節響應的GT-1結合位點。GT-1bindingsiteinlight-regulatedresponsiveness.CAREsCAACTC赤霉素相應相關的順式作用元件。Cis-actingelementsnecessaryandsufficientforgibberellin-regulatedresponsiveness.GTGAmotifGTGA后期花粉形成相關的順式作用元件。Cis-regulatoryelementsinvolvedinlatepollen.GT-1boxGAAAAA病原菌和鹽脅迫響應的順式作用元件。Cis-actingelementsPlaysaroleinpathogen-andsalt-inducedresponsiveness.CACT-motifYACTC4植物葉肉組織特異性的順式調控元件。Cis-Regulatoryelementsformesophyll-specificintheC4plant.Low-CO2GANTTNCCO2響應相關的順式作用元件。Cis-actingelementsinvolvedinCO2-responsive.A-boxTACGTA蔗糖響應的順式作用元件。Cis-actingelementsareresponsibleforsugarrepression.ACGT-motifACGT黃化誘導相關的順式作用元件。Cis-actingelementsinvolvedinetiolation-inducedresponsiveness.O2-REGTAC氧氣脅迫響應的順式作用元件。Cis-actingelementsinvolvedinoxygen-response.GARETAACAAR赤霉素應答相關的元件。GA-responsiveelement.LTRECCGAAA低溫應答元件。Low-temperature-responsiveelement.CircadianCAANNNNATC晝夜節律順式作用元件。Cis-actingregulatoryelementinvolvedincircadiancontrol.GTACGTAC銅和氧氣響應的順式作用元件。Cis-actingregulatoryelementinvolvedcopper-andoxygen-responsive. 圖4 pCAMBIA1304-pTkGGPPS-GFP/GV3101植物表達載體的構建Fig.4 Construction of prokaryotic expression vector of pCAMBIA1304-pTkGGPPS-GFP/GV3101 A: pMD19-T-pTkGGPPS質粒和pCAMBIA1304質粒雙酶切電泳圖 (1~2: pMD19-T-pTkGGPPS質粒雙酶切結果; 3: 陽性質粒對; 4~5: pCAMBIA1304質粒雙酶切結果; 6: 陽性質粒對照);B: pCAMBIA1304-pTkGGPPS-GFP質粒雙酶切電泳圖 (1~2: pCAMBIA1304-pTkGGPPS-GFP質粒雙酶切結果; 3: 陽性質粒對照);C: pCAMBIA1304-pTkGGPPS-GFP/GV3101菌液PCR鑒定電泳圖 (1~14: pCAMBIA1304-pTkGGPPS-GFP/GV3101; 15: 陽性對照; 16: 陰性對照)。M: Trans5K DNA Marker;A: The electrophoregram of pMD19-T-pTkGGPPS plamid digested (1-2: pMD19-T-pTkGGPPS plasmid through double enzyme cut; 3: The positive plamid; 4-5: pCAMBIA1304 plasmid through double enzyme cut; 6: The positive plamid);B: The electrophoregram of pCAMBIA1304-pTkGGPPS-GFP plamid digested (1-2: pCAMBIA1304-pTkGGPPS-GFP plasmid through double enzyme cut; 3: The positive plamid);C: PCR electrophoregram of pCAMBIA1304-pTkGGPPS-GFP/GV3101(1-14: pCAMBIA1304-pTkGGPPS-GFP/GV3101; 15: Positive control; 16: Negative control). 圖5 pTkGGPPS-GFP融合蛋白的活性檢測 Fig.5 Activity detection of pTkGGPPS-GFP fusion protein A: 正常洋蔥表皮細胞; B: 轉染pTkGGPPS-GFP融合基因的洋蔥表皮細胞; C: 轉染pTkGGPPS-GFP融合基因, 經質量分數為40%蔗糖溶液質壁分離后的洋蔥表皮細胞。A: CK; B: Onion epidermal cells of transfectional pTkGGPPS-GFP fusion gene; C: transfectional pTkGGPPS-GFP fusion gene, after a 40% sucrose solution plasmolysis onion epidermal cells. 在分子生物學的研究中,基因克隆和分子雜交探針的制備等操作常需分離與已知DNA序列相近的未知序列;而TAIL-PCR技術作為一種用來分離已知序列旁側的未知序列的一種方法,該技術簡單易行,反應高效靈敏,產物的特異性高,重復性好,能夠在較短的時間內獲得目的片段,已經成為分子生物學研究中一項有用的技術。TAIL-PCR是Liu等[8]1995年設計并對其進行不斷發展完善。Liu等[6,8]用這種方法成功分離了YAC、P1和BAC載體的插入序列和擬南芥T-DNA插入的側翼序列。梁成真等[13]利用優化TAIL-PCR方法,成功地克隆棉花抗逆相關轉錄因子編碼基因。仇艷光等[14]對TAIL-PCR進行改良,并成功地分離小麥基因啟動子。啟動子作為轉錄調控中心一直是研究的熱點,獲得具有功能的啟動子是研究的基礎。本研究隨機選取5對簡并引物,利用改良的TAIL-PCR方法,成功地擴增得到橡膠草GGPPS基因5′側翼序列,其中,引物AD2的特異性較高。 通過分析啟動子序列,發現該啟動子除含有植物啟動子所具有CAAT-box和TATA-box等基本的順式作用元件外,還包括I-box、GARE、W-box、GT-1 box和LTRE等多個與光、植物激素、逆境脅迫等相關的順式作用元件。最為重要的是,該啟動子具有根、葉肉和胚乳特異性的順式作用元件,這與GGPPS基因主要在根、葉和種子的表達的結果相一致[15]。另外,Hefner等[16]研究表明,經茉莉酸甲酯誘導后的紅豆杉細胞GGPPS的RNA水平要明顯高于非誘導的對照細胞;化文平等[17]研究表明,SmGGPPS3基因在丹參不同發育時期不同器官中表達差異顯著,同時受茉莉酸甲酯和病原菌的誘導。這說明該啟動子區域應該有茉莉酸甲酯響應的順式作用元件或相關的轉錄因子,但是該啟動子的生物信息學的預測并沒有發現,原因需要做進一步的研究。 以綠色熒光蛋白為報告基因,利用根癌農桿菌介導的瞬時轉化法對該啟動子進行活性檢測,結果表明本研究克隆獲得的啟動子具有轉錄活性。GGPPS基因在橡膠草中表達具有組織特異性,該啟動子是否是組織特異性啟動子,還需進一步驗證。 本研究利用TALL-PCR成功克隆到橡膠草GGPPS基因5′側翼序列,大小為1131 bp。利用啟動子分析軟件分析,表明該序列不僅含有植物啟動子基本的順式作用元件,還包含相應逆境、植物激素等順式作用元件。以綠色熒光蛋白為報告基因,構建融合表達載體,瞬時轉化洋蔥表皮細胞,發現該啟動子能夠驅動GFP基因的表達,說明該啟動子具有生物活性。橡膠草中橡膠主要在根部合成,利用該啟動子驅動橡膠合成相關功能基因在橡膠草根中過表達,以提高橡膠合成量,為橡膠草產業化生產橡膠提供有力的工具。 [1] Zeng L H,Wang H Y, Xie C C. Clone and identification of granule-bound starch synthase gene promoter in Maize. Plant Physiology Journal, 2015, 51(9): 1433-1439. [2] Archer B L, Audley B G. New aspects of rubber biosynthesis. Botanical Journal of the Linnean Society, 1987, 94: 181-196. [3] Cornish K, Siler D J. Effect of different allylic diphosphates on the initiation of new rubber molecules and on cis-1,4-polyisoprene biosynthesis in guayule (PartheniumargentatumGray). Journal of Plant Physiology, 1995, 147(3-4): 301-305. [4] Liu B H, Wei X D. Tissue culture ofTaraxacumkok-saghyzRodin. Tropical Agricultural Engineering, 2009, 37(13): 5950-5951. [5] Liu Y G, Chen Y L. High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences. Short Technical Reports, 2007, 43(5): 649-656. [6] Liu Y G, Huang N. Efficient amplification of insert end sequences from bacterial artificial chromosome clones by thermal asymmetric interlaced PCR. Plant Molecular Biology Reporter, 1998, 16: 175-181. [7] Miao J, Huo Y M, Yang Y Y,etal. Improvement of hiTAIL-PCR and its application in onion. Shandong Agricultural Sciences, 2009, 10: 1-4. [8] Liu Y G, Whittier R F. Thermal asymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from PI and YAC clones for chromosome walking. Genomics, 1995, 25: 674-681. [9] Zeng J, Huang F Z, Li B C,etal. An optimized method ofAgrobacteriumtumefaciens-mediated GFP subcelluar localization. Genomics and Applied Biology, 2014, 33(1): 159-162. [10] Zhao W T, Wei J H, Meng D,etal. Subcellular localization of a sesquiterpene synthase fromAquilariasinensisutilizing three transient expression system. Chinese Traditional and Herbology Drugs, 2013, 44(23): 3379-3385. [11] Liu X F, Liang W H. Studies on subcellular localization GFP in onion epidermal cells mediated byAgrobacterium. Journal of Henan Normal University (Natural Science), 2009, 37(1): 123-125, 150. [12] Liu H Y, Feng D R, Liu B,etal. Studies on subcellular localizationMpASRin onion epidermal cells mediated byAgrobacterium. Journal of Tropical and Subtropical Botany, 2009, 17(3): 218-222. [13] Liang C Z, Zhang R, Sun G Q,etal. Cloning of sress-related transcription factor gene fromCottonby optimized TAIL-PCR. Cotton Science, 2010, 22(3): 195-201. [14] Qiu Y G, Tian J H, Ge R C,etal. A modified TAIL-PCR and its application in isolating gene promoter of Wheat. Chinese Journal of Biotechnology, 2008, 24(4): 695-699. [15] Li Y M, Feng Y J, Li J,etal. Cloning and expression analysis of theTkGGPPSgene fromTaraxacumkok-saghyz. Plant Physiology Journal, 2016, 52(3): 303-311. [16] Hefner J, Ketchum R E B, Croteau R. Cloning and functional expression of a cDNA encoding geranylgeranyl diphosphate synthase fromTaxuscanadensisand assessment of the role of this prenyltransferase in cells induced for taxol production. Arch Biochem Biophys, 1998, 360(1): 62-74. [17] Hua W P, Song S H, Zhi Y,etal. Cloning and expression analysis ofSmGGPPS3 gene fromSalviamiltiorrhiza. Plant Science Journal, 2014, 32(1): 50-57. [1] 曾禮華, 汪瀚宇, 謝程程, 等. 玉米淀粉合成酶基因GBSS啟動子的克隆與鑒定. 植物生理學報, 2015, 51(9): 1433-1439. [7] 繆軍, 霍雨猛, 楊妍妍, 等. 高效TAIL-PCR的改良及在洋蔥中的應用. 山東農業科學, 2009, 10: 1-4. [9] 曾潔, 黃鳳智, 李本昌. 等. 一種根癌農桿菌介導的GFP亞細胞定位方法的優化. 基因組學與應用生物學, 2014, 33(1): 159-162. [10] 趙文婷, 魏建和, 孟冬, 等. 3種瞬時表達體系研究1個白木香倍半萜合酶的亞細胞定位. 中草藥, 2013, 44(23): 3379-3385. [11] 劉肖飛, 梁衛紅. 根癌農桿菌介導的GFP在洋蔥表皮細胞定位研究. 河南師范大學學報, 2009, 37(1): 123-125, 150. [12] 劉海燕, 馮冬茹, 劉兵. 等. 農桿菌介導的MpASR蛋白在洋蔥表皮細胞的定位研究. 熱帶亞熱帶植物學報, 2009, 17(3): 218-222. [13] 梁成真, 張銳, 孫國清, 等. 優化TAIL-PCR方法克隆棉花抗逆相關轉錄因子編碼基因. 棉花學報, 2010, 22(3): 195-201. [14] 仇艷光, 田景漢, 葛榮朝, 等. TAIL-PCR的改良及其在分離小麥基因啟動子中的應用. 生物工程學報, 2008, 24(4): 695-699. [15] 李永梅, 馮玉杰, 李錦, 等.能源植物橡膠草TkGGPPS基因的克隆及表達分析.植物生理學報, 2016, 52(3): 303-311. [17] 化文平, 宋雙紅, 智媛, 等. 丹參SmGGPPS3基因的克隆及表達分析. 植物科學學報, 2014, 32(1): 50-57. Cloning and transient expression of theGGPPSgene promoter from the energy plantTaraxacumkok-saghyz LI Yong-Mei, FENG Yu-Jie, CAO Xin-Wen, ZHAO Li-Jing, ZHU Jian-Bo, YAN Jie* CollegeofLifeScience,ShiheziUniversity,Shihezi832000,China Based on the known sequence ofGGPPS, which encodes geranylgeranyl pyrophosphate synthase, theGGPPSpromoter sequence was amplified from the root of the perennial herbTaraxacumkok-saghyzby thermal asymmetric interlaced PCR (TAIL-PCR) using nested specific primers. Analyses of the full-length 1131-bp fragment by PlantCare and PLACE software showed that the promoter sequence contained basic cis-acting elements, multiple organ-specific expression elements, and a number of stress-relatedcis-acting elements. The promoter sequence was designated as pTkGGPPS (GenBank: KT901796). The plant expression vector pCAMBIA1304-pTkGGPPS-GFP was constructed using this sequence to replace the CaMV 35S promoter sequence in the plasmid of pCAMBIA1304 withGFPas the reporter gene. The construct was transformed into onion epidermal cells using Agrobacterium tumefaciens, and the promoter successfully drove expression of the GFP gene. The cloning and transient expression of theGGPPSgene promoter fromT.kok-saghyzprovides a reference for further studies on the mechanisms and tissue-specificity of rubber synthesis. Taraxacumkok-saghyz;GGPPSgene; TAIL-PCR; promoter; sequence analysis; transient expression 10.11686/cyxb2016034 http://cyxb.lzu.edu.cn 2016-01-21;改回日期:2016-04-28 國家自然科學基金(No.31360060)資助。 李永梅(1988-),女,山西呂梁人,在讀碩士。E-mail: liyongmeishzu@163.com*通信作者Corresponding author. E-mail:jiey@shzu.edu.cn 李永梅, 馮玉杰, 曹新文, 趙李靖, 祝建波, 閆潔. 能源橡膠草GGPPS基因啟動子的克隆及瞬時表達研究. 草業學報, 2016, 25(12): 180-187. LI Yong-Mei, FENG Yu-Jie, CAO Xin-Wen, ZHAO Li-Jing, ZHU Jian-Bo, YAN Jie. Cloning and transient expression of theGGPPSgene promoter from the energy plantTaraxacumkok-saghyz. Acta Prataculturae Sinica, 2016, 25(12): 180-187.1 材料與方法
2 結果與分析



3 討論
4 結論
