牛乳鐵蛋白肽及其衍生肽生物活性與研究進(jìn)展
2017-01-14 02:02:49郭愛(ài)珍呂自力鐘浩胡曼王亮
中國(guó)乳品工業(yè) 2016年5期
關(guān)鍵詞:系統(tǒng)
郭愛(ài)珍,呂自力,鐘浩,胡曼,王亮
(1.江蘇大學(xué)食品與生物工程學(xué)院,江蘇鎮(zhèn)江212013,2.成都中醫(yī)藥大學(xué)第二附屬醫(yī)院成都610041)
牛乳鐵蛋白肽及其衍生肽生物活性與研究進(jìn)展
郭愛(ài)珍1,呂自力2,鐘浩1,胡曼1,王亮1
(1.江蘇大學(xué)食品與生物工程學(xué)院,江蘇鎮(zhèn)江212013,2.成都中醫(yī)藥大學(xué)第二附屬醫(yī)院成都610041)
牛乳鐵蛋白肽(LfcinB)是牛乳鐵蛋白N-端在正常的消化環(huán)境下水解產(chǎn)生的25個(gè)氨基酸殘基的短肽,具有廣譜高效的抗細(xì)菌性,抗病毒,抗腫瘤等多種生物活性。作為一種新型抗菌肽,LfcinB及其衍生肽具有很大的研究?jī)r(jià)值和潛在的應(yīng)用價(jià)值。對(duì)其結(jié)構(gòu)及生物學(xué)活性進(jìn)行介紹,并對(duì)近年來(lái)采用基因工程技術(shù)表達(dá)LfcinB及其衍生肽的方法技術(shù)進(jìn)行了綜述。
牛乳鐵蛋白肽;衍生肽;生物活性;基因工程
0 引 言
乳鐵蛋白是廣泛存在于哺乳動(dòng)物乳腺的天然轉(zhuǎn)鐵糖蛋白,與血漿中鐵離子的傳遞密切相關(guān)。牛乳鐵蛋白(bovine lactoferrin,LfB)是鐵傳遞蛋白家族成員之一,由689個(gè)氨基酸殘基組成,分子量約為80 ku,具有高效抗菌,消炎,抑制腫瘤細(xì)胞生長(zhǎng)及調(diào)節(jié)機(jī)體免疫等生物功能。牛乳鐵蛋白肽(bovine Lactoferricin,LfcinB)最初是由日本學(xué)者Bellamy W[1,2]等人從牛乳鐵蛋白N端17-41處獲得的陽(yáng)離子抗菌肽,分子量3126.4 u,疏水殘基比例48%,電荷數(shù)+8,顯示其抗菌活性是天然LfB的400多倍。LfcinB是一種新型的兩親性陽(yáng)離子抗菌肽,疏水性與正電荷數(shù)是決定其抗菌效果的主要因素[3]。通過(guò)調(diào)整LfcinB正電荷數(shù)以及疏水性比例獲得的牛乳鐵蛋白肽衍生肽LfcinBD越來(lái)越多的運(yùn)用于基因工程技術(shù)中使其高效大量表達(dá)。由于LfcinB及其衍生肽的廣譜抗菌性,抗病毒,抗腫瘤,良好的耐熱性,無(wú)抗原性,對(duì)腸道有益菌無(wú)害等眾多生物學(xué)活性,作為食品保存劑、抗氧化劑、營(yíng)養(yǎng)強(qiáng)化劑、免疫強(qiáng)化劑、免疫調(diào)節(jié)劑等在食品領(lǐng)域得到廣泛的應(yīng)用[4]。
1 牛乳鐵蛋白肽及其結(jié)構(gòu)
安美忱等人利用高效液相色譜-基質(zhì)輔助激光解吸電離串聯(lián)飛行時(shí)間質(zhì)譜分離得到高活性的LfcinB,相對(duì)分子質(zhì)量為3124.89,蛋白質(zhì)量濃度為18.20μg/mL[5]。LfcinB含25個(gè)氨基酸殘基:FKCRRWQWRMKKLGAPSITCVRRAF,具有廣譜抗菌、抗病毒、免疫調(diào)節(jié)等作用。包括2個(gè)色氨酸,5個(gè)精氨酸和3個(gè)賴氨酸等正電荷氨基酸,p I約為11.84。其中在兩個(gè)半胱氨酸殘基處通過(guò)二硫鍵連接成環(huán)狀,如圖1所示。LfcinB在牛乳鐵蛋白分子中呈α螺旋結(jié)構(gòu),當(dāng)解離下來(lái)后α螺旋結(jié)構(gòu)消失轉(zhuǎn)變成以β折疊為主的兩親性結(jié)構(gòu),利用RCSB Protein Data Bank-RCSB PDB數(shù)據(jù)庫(kù)分析LfcinB結(jié)構(gòu)(圖2)也表明,LfcinB是兩親性分子結(jié)構(gòu),疏水性和親水性氨基酸分別排列于β折疊片層的兩側(cè),而這種結(jié)構(gòu)對(duì)于LfcinB發(fā)揮其強(qiáng)大的抗菌功能是至關(guān)重要的[6]。
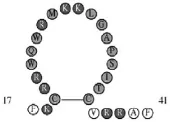
圖1 LfcinB一級(jí)結(jié)構(gòu):紅色代表帶正電荷氨基酸

圖2 LfcinB二級(jí)結(jié)構(gòu)
2 牛乳鐵蛋白肽及其衍生肽的生物學(xué)活性
作為一種有生物活性的短抗菌肽,LfcinB具有普通抗生素所不具有的一些優(yōu)點(diǎn),例如廣譜抗菌性能和不易產(chǎn)生耐藥性問(wèn)題等特點(diǎn)。而且,它在抑制病毒、真菌和腫瘤細(xì)胞的方面也有一定的生物活性,同時(shí),對(duì)真核細(xì)胞幾乎是無(wú)毒性的。
2.1 廣譜抗菌性
LfcinB除不能參與鐵代謝外,具備完整乳鐵蛋白的所有生物學(xué)活性。有研究表明,雖然其抗病毒能力不如完整乳鐵蛋白,但其抗菌性卻比完整乳鐵蛋白強(qiáng)400多倍[1,7]。LfcinB抗菌活性主要表現(xiàn)為廣譜的抑菌和殺菌作用。正電荷氨基酸是抗菌肽與負(fù)電荷的細(xì)菌細(xì)胞壁及細(xì)胞質(zhì)膜直接作用發(fā)揮抗菌效果所必需的,而與脂雙層親脂區(qū)域相互作用需要疏水氨基酸殘基的參與,以產(chǎn)生孔洞或其它的去穩(wěn)結(jié)構(gòu)導(dǎo)致膜的去極化或局部瓦解甚至細(xì)菌死亡。
Bellamy等[8]研究了LfcinB的抗菌譜和抗菌效果,所選擇包括G+和G-、桿菌和球菌、專性需氧菌,兼性厭氧菌和專性厭氧菌,如大腸桿菌(Escherichiacoli)、腸炎沙門氏菌(Salmonella enteritidis)、奇異變形桿菌(Proteus mirabilis)、腸炎沙門氏菌(Salmonella enteritidis)、肺炎克雷伯氏菌(Klebsiella pneumoniae)銅綠假單胞菌(Pseudomonas aeruginosa)、金黃色葡萄球菌(Staphylococcus aureus)、突變鏈球菌(Streptococcus mutans)、白喉棒狀桿菌(Corynebacterium diphtheriae)、雙歧桿菌(Bifidobacterium)等28種菌株。結(jié)果如表1所示。
由表1可以看出,除對(duì)熒光假單胞菌和糞腸道球菌沒(méi)有抑菌作用外,對(duì)其他生長(zhǎng)的檢測(cè)菌株均有不同程度的抑制作用,而且抑菌效果與培養(yǎng)基和菌株有關(guān),其有效抑菌質(zhì)量濃度范圍在0.6~45μg/mL之間,且G+對(duì)LfcinB比G—更為敏感。

表1 牛乳鐵蛋白肽主要抗菌譜
LfcinB除了對(duì)大多數(shù)細(xì)菌有抑殺作用外,還對(duì)某些真菌有抑制作用[9,10],如白色念珠菌等,對(duì)白色念珠菌JCM-2900、白色念珠菌JCM-1542T的最小抑菌濃度(MIC)均為18μg/mL。
2.2 抗病毒生物活性
較完整的乳鐵蛋白來(lái)說(shuō),LfcinB的抗細(xì)菌性和抗真菌性高得多,但LfcinB在體外的抗病毒活性卻較乳鐵蛋白低,大概是完整乳鐵蛋白的1/7。由于分子環(huán)狀結(jié)構(gòu)對(duì)Lfcin抗病毒活性十分重要,而線性化的LfcinB和LfcinH則抗病毒活性喪失[11]。具體的LfcinB抗病毒作用機(jī)制還不是很清楚,可能是LfcinB與細(xì)胞表面的病毒受體或共受體相互作用,從而降低病毒的感染率。但病毒感染包括吸附、侵入、脫殼、生物合成、組裝和釋放多個(gè)階段,而最有效的抗病毒藥劑是能夠影響吸附以及核酸復(fù)制階段從而發(fā)揮作用。Andersen等[12]研究發(fā)現(xiàn),LfcinB與阿昔洛韋聯(lián)合使用,能夠增強(qiáng)阿昔洛韋的抗病毒作用。張曉宇等[13]用pljm 1-LfcinB載體、△8.91載體、pvsvg載體三質(zhì)粒共轉(zhuǎn)染包裝細(xì)胞293T細(xì)胞,包裝產(chǎn)生重組慢病毒載體,得到懸液滴度為2×108TU/mL的濃縮病毒,Western blot檢測(cè)到目的肽的成功表達(dá),為進(jìn)一步研究LfcinB在抗癌細(xì)胞作用機(jī)制和基因治療奠定了基礎(chǔ)。多個(gè)研究均表明,LfcinB具有一定的抗病毒功能,具體的作用機(jī)制還有待進(jìn)一步的探討。
2.3 抗腫瘤生物活性
關(guān)于LfcinB抗腫瘤作用的研究主要都集中于探討其誘導(dǎo)腫瘤細(xì)胞凋亡作用機(jī)制[14],作為一種陽(yáng)離子抗菌肽,其可通過(guò)與腫瘤細(xì)胞暴露出的帶凈負(fù)電荷的磷脂酰絲氨酸PS結(jié)合,以及腫瘤細(xì)胞質(zhì)膜磷脂頭部成分的丟失導(dǎo)致的不對(duì)稱性,擊穿腫瘤細(xì)胞膜,啟動(dòng)細(xì)胞凋亡途徑最終使其凋亡[15]。
王靜等人分析了LfcinB對(duì)Jurkat細(xì)胞增殖周期、細(xì)胞表面分化抗原表達(dá)水平的作用[16]。用LfcinB處理過(guò)的Jurkat細(xì)胞隨著LfcinB濃度增加,處于G 1/G 0期的Iurkat細(xì)胞百分率明顯上升,S期的細(xì)胞百分率明顯下降,細(xì)胞表面分化抗原CD llb表達(dá)水平顯著升高。研究結(jié)果表明,LfcinB能夠通過(guò)細(xì)胞周期阻滯Jurkat細(xì)胞增殖,進(jìn)而誘導(dǎo)其分化。該研究為L(zhǎng)fcinB抗白血病的研究提供了理論基礎(chǔ)。石皖榮等[17]人研究了牛乳鐵蛋白抗菌肽(LfcinB)抗人肺癌細(xì)胞功能及其作用機(jī)制,他們采用MTT法檢測(cè)LfcinB對(duì)人肺腺癌A 549細(xì)胞增殖的影響作用,通過(guò)流式細(xì)胞儀測(cè)定細(xì)胞周期和凋亡率。結(jié)果表明,LfcinB能顯著抑制人肺腺癌A549細(xì)胞增殖,使細(xì)胞阻滯于S期,并誘導(dǎo)細(xì)胞凋亡。其中,當(dāng)LfcinB質(zhì)量濃度為10-2g/L時(shí)抑制率最高。Shusheng Wang[18]等人的研究發(fā)現(xiàn)LfcinB能夠在轉(zhuǎn)錄和翻譯水平抑制肺癌H 460細(xì)胞VEGF(血管內(nèi)皮細(xì)胞生長(zhǎng)因子)及凋亡抑制基因的表達(dá),并在小鼠體內(nèi)成功抑制H460腫瘤細(xì)胞生長(zhǎng);利用雙變量流式細(xì)胞儀通過(guò)Annexin V/PI雙染色法得到的散點(diǎn)圖表明LfcinB能夠有效的誘導(dǎo)細(xì)胞凋亡,當(dāng)LfcinB質(zhì)量濃度為80μg/mL時(shí),H460細(xì)胞凋亡率約達(dá)71.7%。結(jié)果表明LfcinB有可能是治療肺癌的潛在藥物。類似地,minjia Sheng[19]等研究了LfcinB9(WQWRMKKLG)誘導(dǎo)人卵巢癌SK-OV-3細(xì)胞凋亡的機(jī)制,研究發(fā)現(xiàn)LfcinB9成功的抑制了小鼠體內(nèi)SK-OV-3細(xì)胞的增殖,并得出LfcinB9通過(guò)上調(diào)活性氧成分以及在轉(zhuǎn)錄和翻譯水平上激活半胱氨酸蛋白酶-3,半胱氨酸蛋白酶-9的表達(dá)來(lái)誘導(dǎo)細(xì)胞凋亡。
綜上所述,一方面,牛乳鐵蛋白肽LfcinB的高效廣譜抗菌性,抗腫瘤以及免疫調(diào)節(jié)等功能使得其在食品添加劑領(lǐng)域以及嬰幼兒配方奶粉的功能性因子方面有很大的應(yīng)用價(jià)值;另一方面,近年來(lái)全球關(guān)于抗生素耐藥性的威脅已經(jīng)亟需控制,找到新的抗菌性藥物已經(jīng)迫在眉睫[20],LfcinB則是極具潛力的無(wú)耐藥性威脅的新型抗生素。因此,LfcinB及其衍生肽在食品,藥品,化妝品,動(dòng)物飼料等諸多領(lǐng)域有著廣闊的應(yīng)用前景[21]。
3 牛乳鐵蛋白肽及其衍生肽基因工程表達(dá)研究進(jìn)展
由于天然分離LfcinB產(chǎn)量低,工藝復(fù)雜,費(fèi)用較高,人工合成LfcinB及其衍生物成本更高,無(wú)法大規(guī)模生產(chǎn),成為L(zhǎng)fcinB廣泛應(yīng)用的制約因素。因此,利用轉(zhuǎn)基因技術(shù)將LfcinB及其衍生肽轉(zhuǎn)入其他生物表達(dá),是解決生產(chǎn)生活中LfcinB來(lái)源不足最具潛力的途徑。目前表達(dá)LfcinB或LfcinB衍生肽應(yīng)用最多的表達(dá)系統(tǒng)包括兩大類,分別是大腸桿菌表達(dá)系統(tǒng)和畢赤酵母表達(dá)系統(tǒng)。對(duì)兩大系統(tǒng)以及國(guó)內(nèi)外學(xué)者利用兩大系統(tǒng)表達(dá)LfcinB及衍生肽進(jìn)行介紹。
3.1 大腸桿菌表達(dá)系統(tǒng)
大腸桿菌作為表達(dá)外源基因受體菌,有著許多優(yōu)點(diǎn):大腸桿菌培養(yǎng)簡(jiǎn)單,繁殖迅速,遺傳穩(wěn)定,可利用大量的商業(yè)化表達(dá)載體,操作方便。而且對(duì)其在遺傳學(xué),生物化學(xué)和生理學(xué)方面都有了較為充分的認(rèn)識(shí),全基因組測(cè)序,共有4405個(gè)開(kāi)放型閱讀框架,被美國(guó)FDA批準(zhǔn)為安全的基因工程受體生物。因此已廣泛用于外源蛋白的生產(chǎn),且蛋白表達(dá)量較高。
而利用大腸桿菌構(gòu)建重組牛乳鐵蛋白或LfcinB的表達(dá)系統(tǒng)已有研究,Isui和Jose等[22]人對(duì)比了人乳鐵蛋白和牛乳蛋白的三維結(jié)構(gòu)的差別,并成功克隆了牛乳鐵蛋白的cDNA基因,與硫氧還蛋白組成融合蛋白,構(gòu)建pET 32-bLf重組表達(dá)載體,轉(zhuǎn)入E.coliBL21 (DE3)進(jìn)行表達(dá),經(jīng)過(guò)鎳柱分離純化后,獲得的重組牛乳鐵蛋白的產(chǎn)量為15.3 mg/L,純度為90.3%。由于LfcinB抗菌效果大大強(qiáng)于牛乳鐵蛋白,因此更不乏大腸桿菌表達(dá)LfcinB的研究。例如,為了能夠獲得成本低的LfcinB,姚海燕等[23]利用大腸桿菌表達(dá)系統(tǒng)克隆表達(dá)牛乳鐵蛋白十肽及其突變體,為基因工程抗真菌肽的制備奠定了基礎(chǔ)。Z i-gang Tian[24]等人設(shè)計(jì)合成LfcinB15-W 4,10衍生肽多聚體序列,插入表達(dá)載體pET32a,IPTG誘導(dǎo)表達(dá),其中LfcinB15-W 4,10蛋白表達(dá)量最高,高于10 mg/L,回收純度在99%以上。Xingjun Feng[25]等利用大腸桿菌表達(dá)融合了谷胱甘肽轉(zhuǎn)移酶GST標(biāo)簽的LfcinB-W 10,表達(dá)量為20 mg/L,純度為90%。分離純化后的LfcinB-W 10產(chǎn)量達(dá)300μg/L,且對(duì)金黃色葡萄球菌有明顯的抑菌活性。他們還合成雜合的牛乳鐵蛋白肽-天蠶素A,即(LF15-CA 8)n(n=1-6)的多聚體,構(gòu)建pGEX-4T-LCn(n=1–6)載體,轉(zhuǎn)入大腸桿菌BL21 (DE3)表達(dá)。IPTG誘導(dǎo)表達(dá)后,獲得含有? GST-LF15-CA8和GST-(LF15-CA8)2的包涵體,純化后獲得10.0 mg/L,純度高于97%。LF15-CA 8對(duì)革蘭氏陽(yáng)性菌(金葡菌)的抗性高于LfcinB以及CA-8,并且在有效抗菌濃度范圍內(nèi)對(duì)人紅細(xì)胞沒(méi)有顯示出明顯的溶血活性[26]。
雖然大腸桿菌表達(dá)系統(tǒng)是目前掌握最為成熟的表達(dá)系統(tǒng),能夠在較短時(shí)間內(nèi)獲得基因表達(dá)產(chǎn)物,且所需成本相對(duì)較低。但是大腸桿菌缺乏對(duì)真核生物蛋白質(zhì)的復(fù)性及修飾加工系統(tǒng),例如,在原核表達(dá)系統(tǒng)中抗菌肽的二硫鍵無(wú)法正確形成,影響了蛋白的正確折疊使得大腸桿菌表達(dá)系統(tǒng)常常無(wú)法獲得有活性的目的產(chǎn)物;抗菌肽無(wú)法得到酰胺化修飾,從而降低產(chǎn)物活性[27]。另外,內(nèi)源性的蛋白酶降解空間構(gòu)象異常的外源蛋白,加上細(xì)胞周質(zhì)內(nèi)含有種類繁多的內(nèi)毒素,均會(huì)對(duì)目的蛋白的表達(dá)產(chǎn)生不利影響。
4.2 巴斯德畢赤酵母表達(dá)系統(tǒng)
巴斯德畢赤酵母表達(dá)系統(tǒng)是20世紀(jì)80年代初期發(fā)展起來(lái)的一種新型的外源蛋白表達(dá)系統(tǒng),能高效表達(dá)重組蛋白。作為真核生物,畢赤酵母具有高等真核表達(dá)系統(tǒng)的許多優(yōu)點(diǎn),如蛋白加工、折疊及翻譯后修飾等特點(diǎn)[27]。它比桿狀病毒或哺乳動(dòng)物組織培養(yǎng)等其他真核表達(dá)系統(tǒng)更快捷、簡(jiǎn)單、廉價(jià),且表達(dá)水平更高,已越來(lái)越廣泛的應(yīng)用于DNA重組技術(shù)中。
畢赤酵母是甲醇營(yíng)養(yǎng)型酵母,可以利用甲醇作為唯一碳源。圖1顯示了甲醇代謝途徑[28],最初的甲醇代謝在專門微體膜,過(guò)氧化物酶體內(nèi)進(jìn)行,隨后的代謝步驟是在細(xì)胞質(zhì)中進(jìn)行的。中間產(chǎn)物甲醛是在畢赤酵母甲醇和甲胺通路共有的。甲醛在AOX 1通路中是由酒精氧化酶(AOX)與過(guò)氧化氫(H2O2)作用產(chǎn)生的。在甲胺通路中是由胺氧化酶(AmOX)與H2O2和NH3的作用產(chǎn)生的。

圖3 甲醇代謝途徑
畢赤酵母表達(dá)系統(tǒng)具有一系列優(yōu)點(diǎn):首先,醇氧化酶(A1coholoxidase,AOXⅠ)基因的強(qiáng)啟動(dòng)子特別適合于外源基因的調(diào)控表達(dá);其次,所應(yīng)用的表達(dá)載體為整合型穿梭質(zhì)粒,外源基因可以在大腸桿菌中進(jìn)行擴(kuò)增,之后通過(guò)質(zhì)粒整合到畢赤酵母基因組,不易丟失,能夠穩(wěn)定表達(dá);最后,畢赤酵母自身分泌到培養(yǎng)基中的蛋白較少,簡(jiǎn)化了后期蛋白質(zhì)的分離純化操作[29]。下表列舉了用巴斯德畢赤酵母表達(dá)系統(tǒng)生產(chǎn)的外源蛋白產(chǎn)品[28]。

表2 巴斯德畢赤酵母表達(dá)系統(tǒng)生產(chǎn)的外源蛋白產(chǎn)品

(續(xù)表2)
眾多研究表明,在以畢赤酵母為宿主菌的抗菌肽外源表達(dá)中,多數(shù)研究都實(shí)現(xiàn)了目的蛋白的生產(chǎn)且產(chǎn)物對(duì)工程菌沒(méi)有明顯的抑制作用,且可以得到有生物活性的目標(biāo)蛋白。然而由于畢赤酵母自身蛋白酶的降解作用以及所表達(dá)目標(biāo)蛋白的不穩(wěn)定性等因素,制約著畢赤酵母表達(dá)系統(tǒng)的應(yīng)用。為了提高表達(dá)水平,研究學(xué)者采用了各種不同的表達(dá)策略,如融合表達(dá)、改造后表達(dá)等,都有一定的預(yù)期結(jié)果。已有利用巴斯德畢赤酵母表達(dá)系統(tǒng)表達(dá)LF或LFcin的研究。Choi等[33]用畢赤酵母表達(dá)人乳鐵蛋白來(lái)評(píng)估N末端多聚糖結(jié)構(gòu)對(duì)免疫反應(yīng)的影響。Sue-Hong Wang[34]等人利用畢赤酵母表達(dá)重組豬乳鐵蛋白,獲得約12 mg/L的表達(dá)量。Chen[35]等構(gòu)建四種重組山羊乳鐵蛋白肽,GLFcin,GLFcin-(H is)6-Tag,GLFcin II和GLFcin II-(H is)6-Tag,克隆到畢赤酵母表達(dá)載體pPICZαC上,轉(zhuǎn)化畢赤酵母SM D 1168H,這四種肽均獲得表達(dá)。用CM-Sepharose(不含H is-tg),H isT rap(含H is-tg)以及Sephadex G-25 chrom atographies分離純化后可獲得0.15 mg/mL的重組蛋白,對(duì)大腸桿菌BCRC 11549、銅綠假單胞菌BCRC 12450、蠟狀芽孢桿菌BCRC 10603、金黃色葡萄球菌BCRC 25923、痤瘡丙酸桿菌BCRC 10723以及產(chǎn)單核細(xì)胞李斯特菌? BCRC 14845均有較高的抑菌活性,最小抑菌濃度從4.07到16.00 mg/mL不等。林松濤[36]等成功克隆了牛乳鐵蛋白基因的N-lobe的堿基序列,并成功構(gòu)建了酵母表達(dá)載體,為下一步轉(zhuǎn)化進(jìn)行酵母表達(dá)奠定了基礎(chǔ)。易俊波等人[37]將LfcinB基因片段克隆到分泌型表達(dá)載體pPIC 9K中,得到重組質(zhì)粒,線性化電轉(zhuǎn)畢赤酵母SM D 1168。通過(guò)遺傳霉素G 418篩選獲得多拷貝陽(yáng)性轉(zhuǎn)化子,經(jīng)甲醇誘導(dǎo)后,LfcinB在酵母中獲得了表達(dá),表達(dá)產(chǎn)物具有較強(qiáng)的殺菌作用。ChahardooliM[38]等構(gòu)建pH IL-LFcinC表達(dá)載體,利用畢赤酵母表達(dá)系統(tǒng)生產(chǎn)重組亞洲駱駝乳鐵蛋白肽,結(jié)果顯示對(duì)大腸桿菌,金黃色葡萄球菌,枯草芽孢桿菌,綠膿桿菌,白色念珠菌均有顯著的抑菌活性。
5 展 望
而LfcinB作為一種新型的抗菌肽,于食品行業(yè),LfcinB是天然的抑菌和抗氧化成分,可作為嬰幼兒奶粉添加劑,亦可作為調(diào)節(jié)免疫功能因子,添加到食品保健品中;于醫(yī)藥行業(yè),LfcinB可治療疾病的輔助藥物,發(fā)揮其抗病毒抗腫瘤活性;此外,在生化研究方面,動(dòng)物飼料行業(yè),化妝品領(lǐng)域等也有著廣闊的應(yīng)用價(jià)值。利用基因工程技術(shù)手段表達(dá)LfcinB及其衍生肽業(yè)已成為趨勢(shì),而如何使LfcinB及其衍生肽高效表達(dá)成為研究重點(diǎn)。本研究擬通過(guò)分析LfcinB空間結(jié)構(gòu)與生物活性的關(guān)系,優(yōu)化設(shè)計(jì)LfcinB基因獲得LfcinB衍生肽,利用真核表達(dá)系統(tǒng)進(jìn)行表達(dá)活性分析,,并對(duì)培養(yǎng)溫度,甲醇濃度,培養(yǎng)時(shí)間等發(fā)酵條件進(jìn)行優(yōu)化使其能夠高效表達(dá),為提高LfcinB衍生肽的外源表達(dá)量作出有益的探索。
[1]BELLAMY W,TAKASE M,YAMAUCH I K,et al.Identification of the bactericidal domain of lactoferrin[J].Biochim Biophys Acta, 1992,1-2(1121):130-136.
[2]WAKABAYASHI H,TAKASE M,TOM ITA M.Lactoferricin derived from milk protein lactoferrin[J].Current Phamaceutical Design, 2003,9(16):1277-1287.
[3]PANTELEEV P V,BOLOSOV IA,BALANDIN SV,et al.Structure and Biological Functions ofβ-Hairpin Antimicrobial Peptides [J].Acta Naturae,2015,7(1):37-47.
[4]韓兆蓮,許曉曦.新型食品添加劑LfcinB對(duì)腫瘤細(xì)胞生長(zhǎng)的抑制作用[J].食品研究與開(kāi)發(fā),2014,35(23):17-20.
[5]安美忱,劉寧.高效液相色譜-基質(zhì)輔助激光解吸電離串聯(lián)飛行時(shí)間質(zhì)譜分離鑒定牛乳鐵蛋白素[J].色譜,2010,28(2):180-184.
[6]FARNAUD S,EVANSR W.Lactoferrin-amultifuncitional protein w ith antim icrobial properties[J].M olecular Immunology,2003,40(7): 395-405.
[7]GIFFORD JL,HUNTER H N,VOGEL H J.Lactoferricin:a Lactoferrin-derived peptidew ith antimicrobial,antiviral,antitumor and immunological properties[J].Cellular and M olecular Life Science,2005, 62(22):2588-2598.
[8]BELLAMYW,TAKASEM,WAKABASHIK.Antibacteria spectrum of lactoferricin B,a potent bactericldal peptide derived from the N-terminal region of bovine lactoferrin[J].Applied Bacteriology, 1992,73:472-479.
[9]YAMAUCHIK,TOM ITA M.,GIEHL T J,et al.Antibacterial activity of lactoferrin and a pepsin-derived lactoferrin peptide fragment[J]. Infect.Immun,1993,61:719-728.
[10]BELLAMY W,YAMAUCHIK,WAKABAYASHIH,et al.Antifungal propertiesof lact-oferricinB,apeptide derived from the N-terminal region of bovine lactoferrin[J].Lett.Appl.M icrobiol,1994,18: 230-233.
[11]賈洪鋒,賀雅非,劉麗娜,等.乳鐵蛋白及其生理功能.糧食與油脂, 2006,2:44-47.
[12]ANDERSEN J,GUTTEBERG T J.Lactoferrin and lactoferricin inhibitherpessimplex 1 and 2 infection and exhibit synergy when combined w ith acyclovir[J].AntiviralRes,2003,58(3):209-215.
[13]張曉宇,秦宜德,張文曉,等.牛乳鐵蛋白肽基因LfcinB的慢病毒載體的構(gòu)建及鑒定[J].安徽醫(yī)科大學(xué)學(xué)報(bào),2011,46(7):709-712.
[14]張鐵男,楊巍,劉寧.牛乳鐵蛋白素對(duì)Jurkat細(xì)胞和HFL-I細(xì)胞生物學(xué)特性的影響[J].營(yíng)養(yǎng)學(xué)報(bào),2009,31(53):465-469.
[15]FADOK V,CATHELINEAU A,DALEKE D,et al.Loss of phospholipid asymmetry and surface exposure of phosphatidylserine is required for phagocytosis of apoptotic cells by macrophages and fibroblasts[J].BiolChem,2001,276(2):1071-1077.
[16]王靜,趙寧,李萌,等.流式細(xì)胞術(shù)分析牛乳鐵蛋白素對(duì)Jurkat細(xì)胞增殖、分化的影響[J]中國(guó)乳品工業(yè),2014,42(5):9-12.
[17]石皖榮,鄧松華,秦宜德,等.牛乳鐵蛋白抗菌肽抗肺癌細(xì)胞增殖及其作用機(jī)制研究[J].安徽醫(yī)科大學(xué)學(xué)報(bào),2013,48(8):877-881.
[18]WANG S,TU J,ZHOU C,et al.The effect of Lfcin-B on non-small cell lung cancer H 460 cells is mediated by inhibiting VEGF expression and inducing apoptosis[J].Archives of pharmacal research,2014,38(2):261-271.
[19]SHENG M,ZHAO Y,ZHANG A,et al.The effect of LfcinB9 on human ovarian cancer cell SK-OV-3 ismediated by inducing apoptosis[J].Journal of Peptide Science,2014,20(10):803-810.
[20]ROCA I,AKOVA M,BAQUERO F.The global threat of antimicrobial resistance:science for intervention[J].New Microbes New
[21]JENSSEN H.Anti herpes simplex virus activity of lactoferrin/lactoferricin-an example of antiviral activity of antimicrobial protein/peptide [J].Cell Mol Life Sci,2005,62(24):3002-3013.
[22]GARAIA M I,SALAZAR M J,AREVALO G S,et al.Expression and characterization of recombinant bovine lactoferrin in E.coli[J]. Biometals,2013,26(1):113-122.
[23]姚海燕,韓躍武,張雪燕,等.牛乳鐵蛋白十肽的基因改造、克隆及表達(dá)[J].中國(guó)生物制品學(xué)雜志,2008,21(7):568-571.
[24]TIAN Z,TENG D,YANG Y,et al.Multimerization and fusion expression of bovine lactoferricin derivative LfcinB15-W4,10 in Escherichiacoli[J].Applied microbiology and biotechnology,2007,75(1): 117-124.
[25]FENG X,LIU C,GUO J,et al.Expression and purification of an antimicrobial peptide,bovine lactoferricin derivative LfcinB-W10 in Escherichia coli[J].Current microbiology,2010,60(3):179-184. [26]FENG X J,XING LW,LIU D,et al.Design and high-level-expression of a hybrid antimicrobial peptide LF15-CA8 in Escherichia coli[J].Journal of industrial microbiology&biotechnology,2014,41 (3):527-534.
[27]TOWNESC L,MICHAILIDISG,NILEC J,et al,Induction of cationic chicken liver-expressed antimicrobial peptide 2 in response to Salmonella enterica infection[J].Infect Immum,2004,72(12): 6987-6993.
[28]MACAULEY P S,FAZENDA ML,MCNEILl B,et al.Heterologous protein production using the Pichia pastoris expression system [J].Yeast,2005,22(4):249-270.
[29]PANG S Z,OBERHAUS S M,RASMUSSEN J L,et al.Expression of a gene encoding a Scorpion insectotoxin peptide in yeast,bacteria and plants[J].Gene,1992,116(2):165-172.
[30]PARAMSIVAM M,SARAVANAN K,UMA K,et al.Expression, purification and characterization of equine lactoferrin inPichia pastoris[J].Prot Exp Purif,2002,26(1):28–34.
[31]WANG SH,YANG T S,LIN SM,et al.Expression,characterisation and purification of recombinant porcine lactoferrin in Pichia pastoris[J].Prot Exp Purif,25(1):41–49.
[32]GARCIA M I,SALAZAR M J,AREVALO G S,et al.Expression and characterization of recombinant bovine lactoferrin in E.coli[J]. Biometals,2013,26(1):113-122.
[33]CHOI B K,ACTOR J K,RIOSS,et al.Recombinant human lactoferrin expressed in glycoengineered Pichia pastoris:effect of terminal N-acetylneuraminic acid on in vitro secondary humoral immune response[J].Glycoconjugate journal,2008,25(6):581-593.
[34]WANG SH,YANG T S,LIN SM,et al.Expression,Characterization,and Purification of recombinant Porcine Lactoferrin in Pichia pastoris[J].Protein Expr Purif.2002,25(1):41-49.
[35]CHEN G H,CHEN W M,HUANG G T,et al.Expression of recombinant antibacterial lactoferricin-related peptides from Pichia pastoris expression system[J].Journal of agricultural and food chemistry, 2009,57(20):9509-9515.
[36]林松濤,張居農(nóng),王亮,等.牛乳鐵蛋白基因N-lobe的cDNA克隆及畢赤酵母表達(dá)載體的構(gòu)建[J].家畜生態(tài)學(xué)報(bào),2008,29(5):13-18.
[37]易俊波,黃德新,李凌云,等.抗菌肽牛乳鐵多肽素(LfcinB)在畢赤酵母中的表達(dá)及活性鑒定[J].微生物學(xué)報(bào),2007,34(2):265-269.
[38]CHAHARDOOLI M,NIAZI A,ARAM F,et al.Expression of recombinant Arabian camel lactoferricin-related peptide in Pichia pastoris and its antimicrobial identification[J].Journal of the Science of Food and Agriculture,2015.
Biological activity and research progress of Bovine Lactoferricin and derived pep tide
GUO Ai-zhen1,LV Zi-li2,ZHONG Hao1,HU Man1,WANG Liang1
(1.School of Food and Biological Engineering,Jiangsu University,Zhen jiang 212013,China)
(2.The Second Affiliated Hospital of Chengdu University of Traditional Chinese Medicine,Chengdu 610041,china)
Abstrcat:Bovine Lactoferricin(LfcinB)is a short peptide containing 25 amino acid residues and under normal digestive environment it can be released from the N-terminal of Bovine Lactoferrin(LfB).It has many probiotic features,such as broad-spectrum antibacterial,antiviral, antitumor and other biological activities.As a new antibacterial peptide,has a great deal of research value and potential application value.The structure and biological activity of LfcinB and its derived peptides are Introduced,and the expression methods of Lfcin B and its derived peptides using genetic engineering technology in recent years are reviewed.
Bovine Lactoferricin;derived peptides;biological activities;genetic engineering
TS252.1
B
1001-2230(2016)05-0022-06
2015-09-11
四川省科研院所科技成果轉(zhuǎn)化項(xiàng)目(15010116),江蘇大學(xué)高級(jí)專業(yè)人才科研啟動(dòng)資金(12JDG069)。
郭愛(ài)珍(1990-),女,碩士,主要從事牛乳鐵蛋白肽的研究。
王亮
猜你喜歡
工業(yè)設(shè)計(jì)(2022年8期)2022-09-09 07:43:20
軍民兩用技術(shù)與產(chǎn)品(2021年10期)2021-03-16 06:05:30
北京測(cè)繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(shù)(2019年12期)2019-12-25 03:06:46
制造技術(shù)與機(jī)床(2019年10期)2019-10-26 02:47:06
中國(guó)洗滌用品工業(yè)(2019年4期)2019-05-11 09:27:34
鐵道通信信號(hào)(2018年5期)2018-06-28 03:06:24
家庭影院技術(shù)(2017年9期)2017-09-26 03:41:45
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(shù)(2016年6期)2016-04-20 06:21:32
