企鵝珍珠貝Dmrt 2基因的克隆及表達分析
2017-02-09 09:25:36潘珍妮余祥勇王梅芳曲炳良陳耀輝唐小玉于非非
海洋科學 2017年10期
潘珍妮, 余祥勇, 王梅芳, 曲炳良, 陳耀輝, 唐小玉, 于非非
?
企鵝珍珠貝2基因的克隆及表達分析
潘珍妮1, 余祥勇2, 王梅芳2, 曲炳良1, 陳耀輝1, 唐小玉1, 于非非1
(1. 廣東海洋大學水產學院, 廣東 湛江 524088; 2. 華南農業大學海洋學院, 廣東 廣州 510642)
本研究利用RACE-PCR技術獲得企鵝珍珠貝()一個2基因cDNA的全長序列, 通過熒光定量PCR分析2基因在各組織中的表達特征, 以及在早期卵巢、成熟期卵巢、早期精巢、成熟期精巢和排放期精巢中的表達變化。結果表明,2基因全長1 257bp, 其中開放閱讀框(open reading frame, ORF)為951 bp, 編碼316個氨基酸, 5′非編碼區(untranslated region, UTR)為52 bp, 3′UTR為 254 bp, 第20 位到第73位氨基酸為DM結構域。預測其分子質量為36.61ku, 等電點為9.80。氨基酸序列比對顯示該企鵝珍珠貝2基因與黑蝶真珠蛤()和馬氏珠母貝()的2基因同源性(identity)最高, 分別為46.0%和45.7%。其中在DM結構域高度同源。熒光定量PCR分析組織表達特征顯示,2在企鵝珍珠貝的外套膜、鰓、消化盲囊、足、精巢和卵巢均有表達, 其中在精巢中表達量最高(<0.05), 足為其次, 在閉殼肌中沒有檢測到2表達。對性腺發育不同時期的表達量分析發現,2在發育早期和成熟期卵巢中表達量都很低, 在發育早期精巢中表達量較高, 在成熟期精巢檢測到最大表達量(<0.05), 到精巢排放期表達顯著下降, 推測2可能與企鵝珍珠貝精巢的發育有關, 可能參與了企鵝珍珠貝雄性性別分化和性腺發育這一生理過程。
企鵝珍珠貝();2; 基因克隆; 表達分析; 性別決定和性腺發育
(doublesex and mab-3-related transcription factor)基因家族是指與果蠅()性別決定基因()和秀麗隱桿線蟲()性別決定基因()同源的一類基因[1], 是一個古老的與性別決定有關的基因家族[2]。該家族基因編碼的蛋白質含有一個高度保守的DM結構域, 通過鋅指結構識別并結合特定DNA來調控目的基因的轉錄, 從而參與性別決定與發育[3-4]。迄今為止已發現了基因家族多個成員, 其生理作用涉及性別決定、胚胎發育和調控肌肉發育等多個方面[5-6]。2基因是t基因家族的重要組成成員之一, 被認為對胚胎中期體節形成以及胚胎后期器官形成起到重要作用[7-8],但是否參與了性別決定和分化則沒有定論。在人類基因圖譜中, 基因1、2和3位于9號染色體短臂2區4帶3亞帶(9p2.4.3), 該片段缺失會引起精巢發育不全, 甚至引起雄性胚胎向雌性逆轉; 但在該片段缺失的性逆轉患者中,1和2均缺失, 由于1是性別決定與分化的關鍵基因, 因此無法確定性逆轉中2是否起到關鍵作用[9]。隨著研究的深入, Matsushita等[10]研究發現2在粗皮蛙()精巢中表達, 可能參與精子的形成以及性別的分化。Yu 等[11]通過免疫組化分析發現2基因主要在馬氏珠母貝()的精巢中表達, 在卵巢中未檢測到表達, 暗示2基因參與馬氏珠母貝精子形成過程。
企鵝珍珠貝(), 屬軟體動物門(Mollusca), 雙殼綱(Bivalvia), 珍珠貝科(Pteriidae), 珍珠貝屬()[12]。企鵝珍珠貝具有生長速度快、珍珠質分泌速度快的特點, 是培育優質海水珍珠的大型珍珠貝[13]。本研究建立了企鵝珍珠貝轉錄組數據庫, 利用RACE-PCR技術克隆到企鵝珍珠貝2基因的cDNA全長序列, 利用熒光定量PCR技術分析2基因在企鵝珍珠貝各組織中的表達差異以及性腺發育不同時期中的表達差異, 以闡述2在企鵝珍珠貝性腺發育中的重要作用。
1 材料與方法
1.1 材料
本實驗所用企鵝珍珠貝采自廣西北海潿洲島野生群體, 300~350 g。
1.2 企鵝珍珠貝Dmrt2基因cDNA全長克隆
使用上海生工UNIQ-10柱式Trizol總RNA抽提試劑盒分別提取企鵝珍珠貝各組織(外套膜、鰓、閉殼肌、消化盲囊、足、性腺)的總RNA, 按照SMARTer?RACE 5′/3′Kit說明書合成企鵝珍珠貝2的3′RACE模板和5′RACE模板。根據企鵝珍珠貝轉錄組測序結果, 設計企鵝珍珠貝2基因5′RACE的特異性引物和3′RACE的特異性引物(表1)。采用巢式PCR擴增法進行5′RACE和3′RACE擴增, 5′RACE和3′RACE PCR反應體系為2×SeqAmp Buffer 10 μL, SeqAmp DNA Polymerase 0.5μL, 5′RACE或3′RACE cDNA模板(100 ng/μL) 1μL, 10×UPM (10×Universal Primer Mix) 2 μL , 5′RACE或3′RACE outer引物(見表1)(10μmol/L) 0.5 μL, PCR-Grade H2O 6 μL。使用touch-down PCR程序擴增: 94℃ 30 s, 72℃ 2 min, 5個循環; 94℃ 30 s, 70℃ 30 s, 72℃ 2 min, 5個循環; 94℃ 30 s, 68℃ 30 s, 72℃ 2 min, 25個循環。第二輪巢式PCR擴增反應體系為50μL: 第一輪PCR擴增產物2μL, 5′RACE或3′RACE inner引物(10μmol/L) 2μL, NUP引物(10μmol/L) 2μL, 2×EasyTaq?PCR SuperMix 25μL, PCR-Grade H2O 19μL。得到的PCR產物經電泳分離、純化后與pEASY?-T1 Cloning Vector連接, 轉化, 過夜培養。篩選陽性克隆測序, 將獲得的 3′端序列與 5′端序列進行拼接。根據拼接序列設計3′和5′驗證引物, PCR驗證拼接序列的正確性, 從而獲得2 cDNA 全長序列。利用VecScreen軟件(http: //www.ncbi.nlm.nih.gov/VecScreen/VecScreen. html)去除載體序列, 獲得企鵝珍珠貝2基因的cDNA全長序列。
1.3 Dmrt2基因的生物信息學分析
通過ORF Finder工具, 推導企鵝珍珠貝2基因編碼的氨基酸序列; 通過Expasy網站(http: // web.expasy.org/compute_pi/)分析其相對分子質量和等電點; 經ProtScale軟件分析氨基酸序列的疏水性; 經SignalP 4.1、TMHMM和SoftBerry-Psite分別預測其信號肽、跨膜結構及氨基酸基本功能位點; 經SMART對其進行功能結構域分析; 利用Clustal X (1.8)軟件進行氨基酸同源比對, 用MEGA6軟件構建系統進化樹。
1.4 熒光定量PCR檢測目的基因的表達特征
分別提取企鵝珍珠貝外套膜、鰓、閉殼肌、消化盲囊、足、精巢和卵巢的RNA, 反轉錄合成第一鏈cDNA, 進行熒光定量PCR檢測。以18S rRNA基因作為內參, 以企鵝珍珠貝的外套膜為對照, 采用2–DDCt法計算2基因在不同組織中的表達水平, 每組設3個重復, 組間重復性和差異顯著性用SPSS16.0分析。分別提取企鵝珍珠貝發育早期卵巢、成熟期卵巢、發育早期精巢、成熟期精巢和排放期精巢的RNA, 反轉錄合成第一鏈cDNA, 進行熒光定量PCR。方法同上。

表1 企鵝珍珠貝Dmrt2基因克隆及表達分析引物
2 結果與分析
2.1 企鵝珍珠貝Dmrt2基因的克隆和序列分析
運用RACE-PCR快速擴增技術得到2基因的cDNA全長序列并分析。企鵝珍珠貝的2基因cDNA全長1257bp, 其中開放閱讀框(ORF)為951 bp, 編碼316個氨基酸, 5′非編碼區(UTR)為52 bp, 3′UTR為 254 bp, 包含30 bp的polyA尾巴, 如圖1所示。登錄號為: KY670720.1。
2.2 Dmrt2基因蛋白理化性質預測和分析
經ExPASy預測企鵝珍珠貝2蛋白分子質量為36.61ku, 等電點為9.80。疏水性分析得知2氨基酸序列總平均親水系數為–0.843, 預測該蛋白屬于親水性蛋白。經TMHMM 2.0進行跨膜結構域預測, 顯示沒有跨膜區。SignalP4.1 預測其不存在信號肽, 屬于非分泌型蛋白質。SoftBerry-Psite在線預測該序列功能位點, 發現酪蛋白激酶II磷酸化位點6個, N-糖基化位點和N-豆蔻酰化位點各一個, 蛋白激酶C磷酸化位點9個, CAAX box 1個, C-末端定位信號序列微體6個。同時使用SMART軟件分析得出第20位到第73位氨基酸為DM結構域(圖1)。
2.3 Dmrt2氨基酸序列同源性分析
采用Clustal X軟件將企鵝珍珠貝的2基因與其他物種的2基因編碼氨基酸進行多序列比對, 結果顯示, 該基因與黑蝶真珠蛤(, AIE16095.1)和馬氏珠母貝(, ADD97887.1)的2基因同源性(identity)最高, 分別為46.0%和45.7%; 與人(, AAF86295.1)和小鼠(, EDL41636.1)的2基因同源性最低, 都為27.5%。其中, 保守位點主要集中在DM結構域(圖2)。將該企鵝珍珠貝2基因命名為2。

圖1 企鵝珍珠貝Dmrt2基因序列分析
小寫字母代表非編碼序列(UTR); 大寫字母代表開放閱讀框(ORF); 方框代表起始密碼子和終止密碼子; 灰色框代表DM結構域
5′UTR and 3′UTR regions are shown in small letters; open reading frame is shown in capital letters; initiation codon and termination codon are shown in boxes; DM domain is shown in gray boxes
分別選取馬氏珠母貝、黑蝶真珠蛤、光滑雙臍螺(, XP_013083870 )creb、鴨嘴海豆芽(, XP_013397304.1)、尾曳鰓蟲(, XP_014675559.1)和熱帶爪蟾(AAI35448.1)等物種的2基因編碼的氨基酸進行聚類分析, Neighbor-joining(NJ)樹如圖3所示: 企鵝珍珠貝、馬氏珠母貝和黑蝶真珠蛤都屬于軟體動物瓣鰓綱, 其2編碼的氨基酸聚為一個小的進化分支; 光滑雙臍螺、鴨嘴海豆芽和尾曳鰓蟲聚為一個小的進化分支; 以上所有的動物聚為一個大的進化分支, 而熱帶爪蟾這一脊椎動物作為外群物種單獨列為一個進化分支。
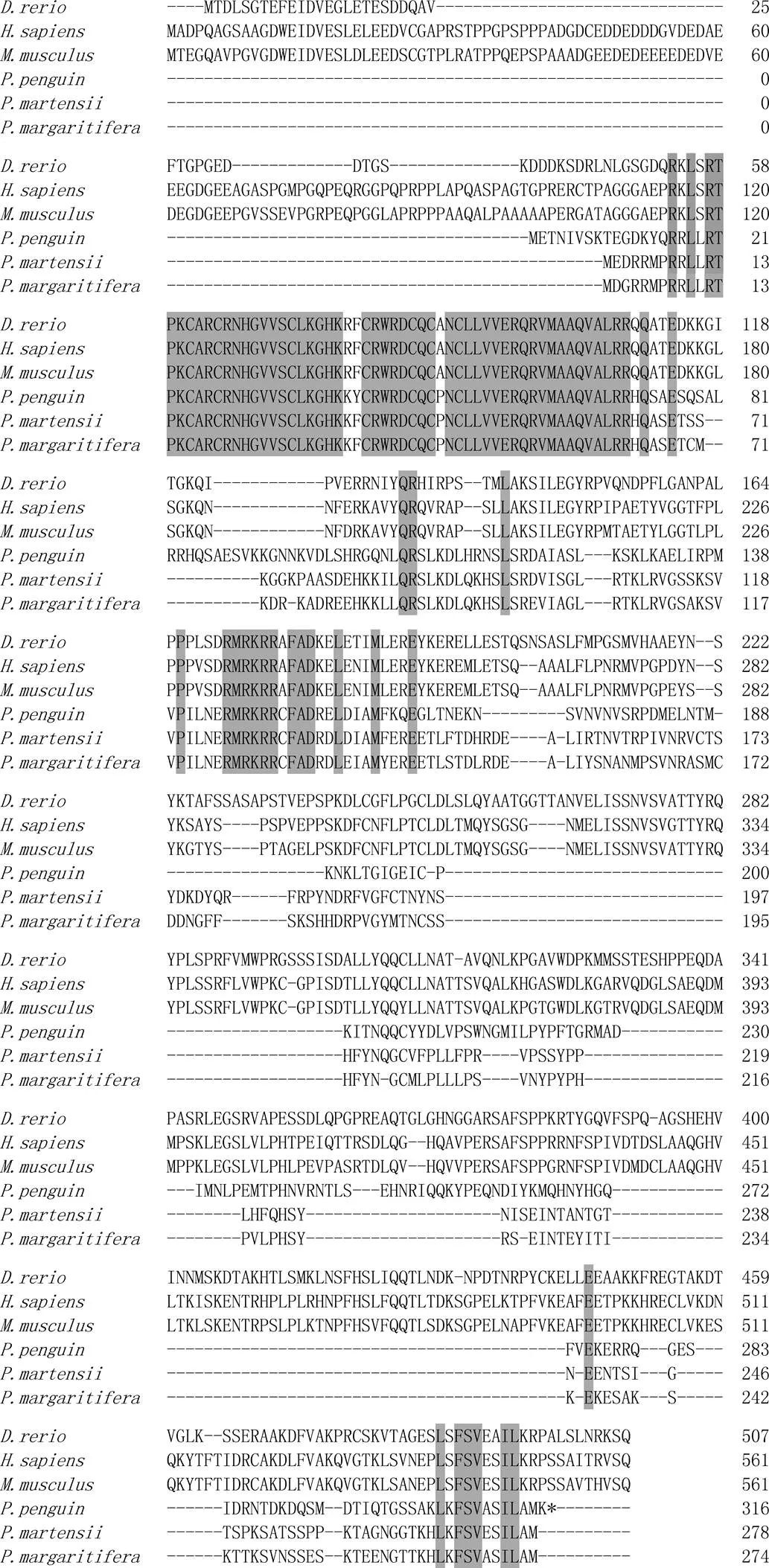
圖2 企鵝珍珠貝與其他物種Dmrt2基因編碼氨基酸比對
采用Clustal W軟件對企鵝珍珠貝()、斑馬魚(NP_571027.1)、人(AAF86295.1)、小鼠(EDL41636.1)、馬氏珠母貝(ADD97887.1)和黑蝶真珠蛤(AIE16095.1)的2基因氨基酸序列進行排序, 相同的氨基酸涂灰色表示
The alignment of2 among,, andby ClustalW. Those amino acids with 100% identity are marked in gray
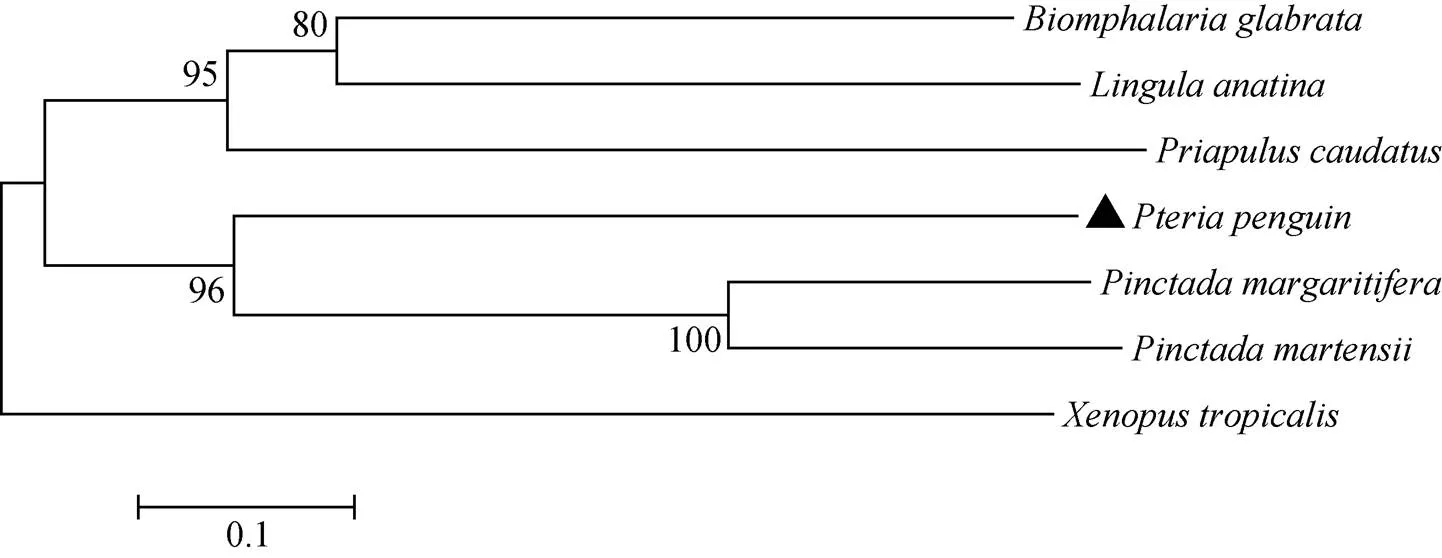
圖3 企鵝珍珠貝與其他物種Dmrt2氨基酸序列的系統進化樹(NJ)
2.4 基因的組織定量分析
采用qRT-PCR技術分析2基因在企鵝珍珠貝外套膜、鰓、閉殼肌、消化盲囊、足、精巢和卵巢中的組織表達差異, 結果顯示(圖4), 除在閉殼肌未檢測到2基因的表達外, 在企鵝珍珠貝各個組織中均檢測到2基因表達。其中, 在精巢中表達量最高(<0.05), 其次是足和消化盲囊, 在外套膜、鰓和卵巢中的表達量較低。
不同發育時期性腺的qRT-PCR分析發現, 在發育早期雌性性腺、成熟期雌性性腺、發育早期雄性性腺、成熟期雄性性腺、排放期雄性性腺中都可檢測到2基因轉錄本的存在, 但在雄性性腺中表達水平顯著高于各期雌性性腺, 其中成熟期雄性性腺中表達量最高, 顯著高于發育早期和排放期雄性性腺(<0.05, 圖5)。這暗示著2基因可能參與了企鵝珍珠貝的雄性分化和性腺發育。
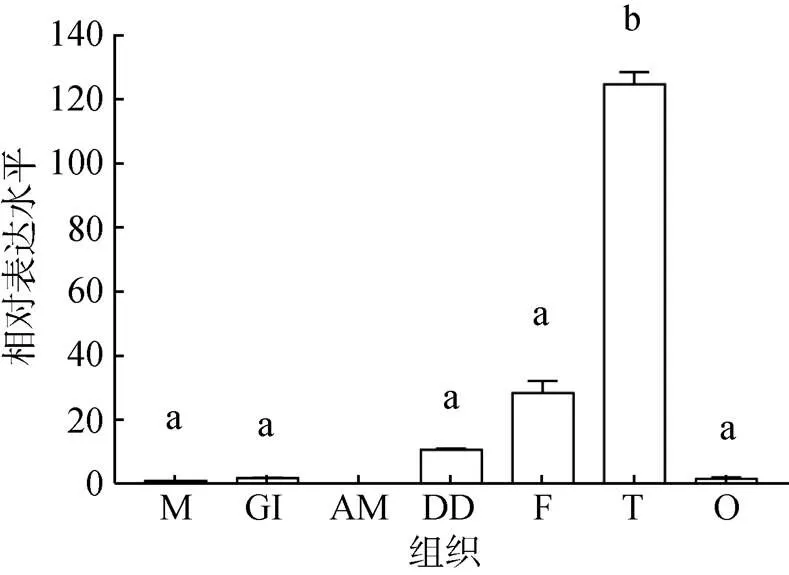
圖4 Dmrt2基因在企鵝珍珠貝各組織的表達水平
M. 外套膜, G. 鰓, AM. 閉殼肌, DD. 消化盲囊, F. 足, T. 精巢, O. 卵巢相同字母表示差異不顯著(>0.05), 不同字母表示差異顯著(<0.05)
M. mantle; G. gill; AM. adductor muscle; DD. digestive diverticulum; F. foot; T. testis; O. ovary The same letters indicate no significant difference (>0.05); different letters indicate significant difference (<0.05)
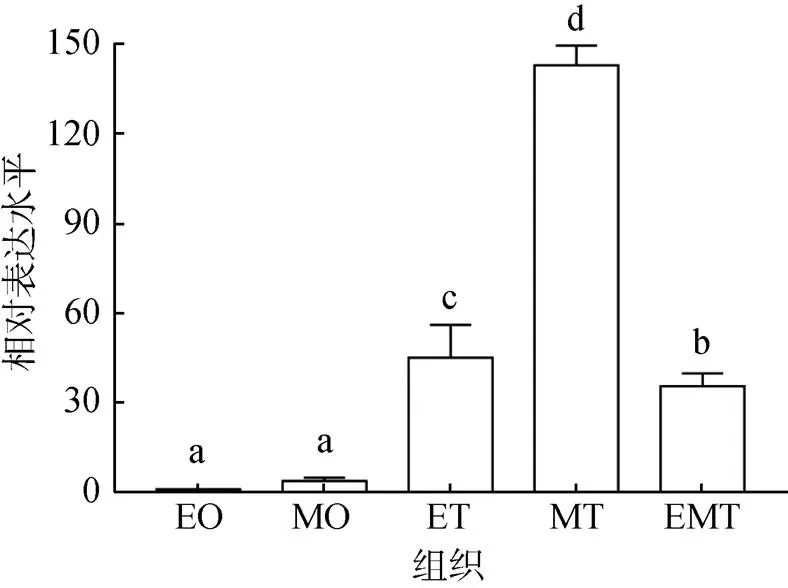
圖5 Dmrt2基因在企鵝珍珠貝不同時期性腺的表達水平
EO. 發育早期卵巢, MO. 成熟期卵巢, ET. 發育早期精巢, MT. 成熟期精巢, EMT. 排放期精巢相同字母表示差異不顯著(>0.05), 不同字母表示差異顯著(<0.05)
EO. early ovary; MO. mature ovary; ET. early testis; MT. mature testis; EMT. emission testis.The same letters indicate no significant difference (>0.05); different letters indicate significant difference (<0.05)
3 討論
目前已在不同物種中發現了多個家族成員[14],其中哺乳動物發現了8個基因, 分別命名為1~8[15], 鳥類中發現了14共4個基因[16], 爬行類發現了16共6個基因[17], 兩棲類發現了15共5個基因[18], 魚類中發現了15共5種[19]。軟體動物主要發現了15共5個基因[20]。在已發現的基因家族中, 除了人類與小鼠的8基因編碼蛋白質的DM結構域不完整外, 其他基因均含有保守的DM結構域, 且DM結構域的序列成為家族成員命名的有力佐證。Wang等[21]發現12基因只含有DM結構域, 而35同時含有DM結構域和DMA結構域。本研究獲得的企鵝珍珠貝基因編碼的蛋白質序列只含有DM結構域; 同源性分析顯示, 該企鵝珍珠貝基因與馬氏珠母貝2基因具有最高的同源性, 兩者相互印證, 故確定了該企鵝珍珠貝基因為2。
近年來對多個物種2基因的組織表達特征進行了分析, 許寶紅等[22]研究發現2在中國大鯢()的精巢和肌肉中高表達, Yamaguchi等[23]報道在成年的紅鰭東方鲀中()的眼、鰓、心臟、腸、頭腎、肝臟、肌肉和脾都有表達, 但在性腺中表達最高。2在斑馬魚中的精巢、卵巢、腦、肌肉和肝中都有表達, 其中在肌肉中表達量最高, 其次是精巢, 且2在精巢中的表達量高于卵巢中的表達[24]。成魚青鳉()2基因在精巢和卵巢中表達[25]。基因在黑蝶真珠蛤中的精巢中的高表達, 而在卵巢中只有微弱表達[26]。本文利用qRT-PCR技術檢測2基因在不同組織中以及在雄性和雌性性腺中的表達情況, 結果顯示,2基因在精巢中的表達量最高, 尤其在成熟精巢中表達量顯著高于發育期精巢和排放期精巢, 而在卵巢及其他組織中的表達量低, 這與已有報道相吻合。
因2和1在染色體上的位置較近, 而已確定1是魚類[27]、兩棲類[28]、爬行類[29]、鳥類[30]、哺乳類[31]的重要性別決定和分化基因, 因此2的功能曾經是有爭議的, 有的學者認為是因為1的突變或缺失影響了2的功能才導致動物由雄性轉為雌性,2本身并不是性別分化和性腺發育的關鍵基因。但是, 越來越多的研究表明2參與了性別分化和性腺發育, 如Yu等[11]對馬氏珠母貝中雌雄性腺發育不同階段的原位雜交實驗顯示,2在馬氏珠母貝雄性性腺成熟和性別分化過程中起了重要作用。結合我們的實驗結果和已有報道, 我們推測在與馬氏珠母貝親緣關系很近的企鵝珍珠貝的雄性性別分化和性腺發育過程中,2可能與起到了重要作用。
此外,2基因在企鵝珍珠貝的足中也有較高的表達, 足是貝類的運動器官, 足中通常有4對收縮肌, 即前后收足肌, 前伸足肌和和中舉舉足肌各一對[32]。這可能與在肌肉中的高表達相一致[22, 24]。
[1] Raymond C S, Shamu C E, Shen M M, et al. Evidence for evolutionary conservation of sex-determining genes[J].Nature, 1998, 391(6668): 691-695.
[2] Picard M A, Cosseau C, Mouahid G, et al. The roles of(Double sex/Male-abnormal-3 Related Transcription factor) genes in sex determination and differentiation mechanisms: Ubiquity and diversity across the animal kingdom[J]. Comptes Rendus Biologies, 2015, 338(7): 451-462.
[3] Erdman S E, Burtis K C. The Drosophila doublesex proteins share a novel zinc finger related DNA binding domain[J]. The EMBO Journal, 1993, 12(2): 527-535.
[4] Matson C K, Zarkower D. Sex and the singular DM domain: insights into sexual regulation, evolution and plasticity[J]. Nature reviews. Genetics, 2012, 13(3): 163-174.
[5] Ottolenghi C, Veitia R, Barbieri M, et al. The human doublesex-related gene,, Is homologous to a gene involved in somitogenesis and encodes a potential bicistronic transcript[J]. Genomics, 2000, 64(2): 179- 186.
[6] Sato T, Rocancourt D, Marques L, et al. A Pax3/Dmrt2/ Myf5 regulatory cascade functions at the onset of myogenesis[J]. PLOS Genetics, 2010, 6(4): e1000897.
[7] Liu S, Li Z, Gui J F. Fish-specific duplicated dmrt2b contributes to a divergent function through Hedgehog pathway and maintains left-right asymmetry establishment function[J]. PLOS One, 2009, 4(9): e7261.
[8] Matsui T, Sasaki A, Akazawa N, et al. Celf1 regulation of dmrt2a is required for somite symmetry and left-right patterning during zebrafish development[J]. Development, 2012, 139(19): 3553-3560.
[9] Quinonez S C, Park J M, Rabah R, et al. 9p partial monosomy and disorders of sex development: Review and postulation of a pathogenetic[J]. American Journal of Medical Genetics Part A, 2013, 161(8): 1882-1896.
[10] Matsushita Y, Oshima Y, Nakamura M. Expression of DMRT genes in the gonads of Rana rugosa during sex determination[J]. Zoological Science, 2007, 24(1): 95- 99.
[11] Yu F F, Wang M F, Zhou L, et al. Molecular Cloning and Expression Characterization ofin Akoya Pearl Oysters, Pinctada martensii[J]. Journal of Shellfish Research, 2011, 30(2): 247-254.
[12] 蒙釗美, 李有寧, 邢孔武. 珍珠養殖理論與技術[M]. 北京: 科學出版社, 1996: 39-41. Meng Zhaomei, Li Youning, Xing Kongwu. The theory and technoligy of the pearl cultivation[M]. Beijing: Science Press, 1996: 39-41.
[13] 馮永勤, 梁盛, 梁曉春, 等. 企鵝珍珠貝游離有核珍珠培育技術[J]. 水產科技情報, 2016, 43(1): 10-14. Feng Yongqin, Liang Sheng, Liang Xiaochun, et al. The cultivation technology of round pearl in Pteria penguin[J]. Fisheries Science & Technology Infomation, 2016, 43(1): 10-14.
[14] Wexler J R, Plachetzki D C, Kopp A. Pan-metazoan phylogeny of the DMRT gene family: a framework for functional studies[J]. Development Genes and Evolution, 2014, 224(3): 175-181.
[15] Hong C S, Park B Y, Saint J P. The function of Dmrt genes in vertebrate development it is not just about sex[J]. Developmental Biology, 2007, 10(1): 1-9.
[16] 汪海, 王婷茹, 袁靜, 等. 脊椎動物Dmrt基因的研究進展[J]. 貴州農業科學, 2012, 40(5): 148-152. Wang Hai, Wang Tingru, Yuan Jing, et al. Advances in Dmrt gene family of vertebrate[J]. GuizhouAgricultural Sciences, 2012, 40(5): 148-152.
[17] Wang Z, Miyake T, Edwards S V, et al. Tuatara (Sphenodon) genomics BAC library construction, sequence survey, and application to the DMRT gene family[J]. Journal of Heredity, 2006, 97(6): 541–548.
[18] Bewick A J, Anderson D W, Evans B J. Evolution of the closely related, sex-related genes DM-W and DMRT1 in African clawed frogs (Xenopus)[J]. Evolution, 2011, 65(3): 698-712.
[19] 曹謹玲, 陳劍杰, 甘西. 魚類DMRT基因的研究進展[J].廣東海洋大學學報, 2011, 31(01): 94-98. Cao Jinling, Chen Jianjie, Gan Xi. Advances in the DMRT genes research of fishes[J]. Journal of GuangdongOceanUniversity, 2011, 31(1): 94-98.
[20] 李法君, 付春鵬, 羅永巨. Dmrt基因在水生生物中的研究進展[J]. 水生生物學報, 2016, 40(5): 1068-1077. Li Fajun, Fu Chunpeng, Luo Yongju. Research progress of Dmrt genes in hydrobiontes[J]. Acta Hydrobiologica Sinica, 2016, 40(5): 1068-1077.
[21] Wang F, Yu Y, Ji D, et al. The DMRT gene family in amphioxus[J]. Journal of Biomolecular Structure and Dynamics, 2012, 30(2): 191-200.
[22] 許寶紅, 肖真明, 肖調義, 等. 大鯢2基因cDNA的克隆與表達分析[J]. 水產學報, 2012, 36(8): 1179-1184. Xu Baohong, Xiao Zhenming, Xiao Diaoyi, et al. Molecular cloning and expression analysis of2 gene in[J]. Journal of Fisheries of China, 2012, 36(8): 1179-1184.
[23] Yamaguchi A, Lee K H, Fujimoto H, et al. Expression of the DMRT gene and its roles in early gonadal deve-lop-ment of the Japanese pufferfish[J]. Comparative Biochemistry and Physiology Part D: Ge-nomics Proteomics, 2006, 1(1): 59-68.
[24] Zhou X, Li Q, Lu H, et al. Fish specific duplication of2: Characterization of zebrafish2b[J]. Biochimie, 2009, 90(6): 878-887.
[25] Winkler C, Hornung U, Kondo M, et al. Developmentally regulated and non-sex-specific expression of autoso-mal dmrt genes in embryos of the Medaka fish ()[J]. Mechanisms of Development, 2004, 121(7- 8): 997-1005.
[26] Teaniniaraitemoana V, Huvet A, Levy P, et al. Gonad transcriptome analysis of pearl oysteridentification of potential sex differentiation and sex determining genes[J]. BioMed Central Genomics, 2014, 15(1): 1-20.
[27] 楊東, 余來寧. 魚類性別與性別鑒定[J]. 水生生物學報, 2006, 30(2): 221-226. Yang Dong, Yu Laining. Sex and sex identification of fish[J]. Acta Hydrobiologica Sinica, 2006, 30(2): 221- 226.
[28] Osawa N, Oshima Y, Nakamura M. Molecular cloning of Dmrt1 and its expression in the gonad of Xenopus[J]. Zoological Science, 2005, 22(6): 681-687.
[29] Sreenivasulu K, Ganesh S, Raman R. Evolutionarily conserved, DMRT1, encodes alternatively spliced transcripts and shows dimorphic expression during gonadal differentiation in the lizard, Calotes versicolor[J]. Mechanisms of Development, 2002, 119 Suppl 1(7574): S55.
[30] Shetty S, Kirby P, Zarkower D, et al. DMRT1 in a ratite bird: evidence for a role in sex determination and discovery of a putative regulatory element[J]. Cytogenetic & Genome Research, 2002, 99(1-4): 245-251.
[31] Pask A J, Behringer R R, Renfree M B. Expression of DMRT1 in the mammalian ovary and testis--from marsupials to mice[J]. Cytogenetic & Genome Research, 2003, 101(3-4): 229-236.
[32] 梁象秋, 方紀祖, 楊和荃. 水生生物學(形態和分類)[M]. 北京: 中國農業出版社, 2002: 250-251.Liang Xiangqiu, Fang Jizu, Yang Hequan. Aquatic biology (morphology and classification)[M]. Beijing: China Ag-ri-cuture Press, 2002: 250-251.
(本文編輯: 梁德海)
Molecular cloning and expression analysis of2 gene from
PAN Zhen-ni1, YU Xiang-yong2, WANG Mei-fang2, QU Bing-liang1, CHEN Yao-hui1, TANG Xiao-yu1, YU Fei-fei1
(1. Fisheries College, Guangdong Ocean University, Zhanjiang 524088, China; 2. School of Marine Science, South China Agricultural University, Guangzhou 510642, China)
2 (doublesex and mab-3-related transcription factor 2) is an important member of thegene family with a highly conserved zinc finger-like DM domain and plays an important role in somite formation, organ formation, and skeletal patterning. This study was conducted to investigate the role of2 in sex determination and gonadal development in. The full-length cDNA of2 gene was characterized fromby RACE-PCR, and real-time PCR was performed to assess the expression of2 in different tissues and gonads of different growth periods. The results showed that the full-length cDNA of2 was 1257 bp, including a 5′UTR of 52 bp, a 3′UTR of 254 bp, and an open reading frame of 951 bp, which encoded a deduced protein of 317 amino acids. The DM domain was from 20 to 73 amino acids. The predicted molecular weight was 36.61 ku, and the isoelectric point was 9.80. The amino acid sequence alignment showed that2 ofshared 46.0% and 45.7% sequence identity with2 ofand, respectivelyThe real-time PCR results showed that2 was constitutively expressed in the mantle, gill, digestive diverticulum, foot, testis, and ovary, except in the adductor muscle. The expression level was highest in the testis (< 0.05), followed by the foot. The expression analysis in gonads of different growth periods revealed that2 had lower expression in early ovary and mature ovary, higher expression in early testis and emission testis, and the highest expression in mature testis (< 0.05). This study showed that2 might play a role in sex determination and gonadal development in
;2; gene cloning; sex determination and gonadal development
Feb. 17, 2017
Q341
A
1000-3096(2017)10-0117-08
10.11759/hykx20170417002
2017-02-17;
2017-04-12
廣東省科技發展專項(2016A020210115); 廣東省漁港建設和漁業發展專項(B201601-Z08, Z2014005); 廣東海洋大學優秀青年教師項目(2014004); 廣東海洋大學博士科研啟動項目(E15041); 廣東海洋大學創新強校重大科研項目(GDOU2016050248)
[Guangdong Provincial Science and Technology Program, No. 2016A020210115; Guangdong Marine Fishery Development Foundation No. B201601-Z08, No.Z2014005; Outstanding Young Teacher Foundation, No.2014004; Doctoral Scientific Research Foundation of GuangdongOceanUniversity, No.E15041; Guangdong Major Project of Innovation School, No.GDOU2016050248]
潘珍妮(1990-), 女, 碩士研究生, 研究方向為海洋經濟動物發育生物學, E-mail: pzn52020@163.com; 于非非, 通信作者, 副教授, E-mail: yufeifei2000@163.com
