溫度對木薯葉片葉綠素熒光及Rubisco酶的影響
2017-02-10 03:32:20張振文林立銘余厚美李開綿陳松筆
江西農(nóng)業(yè)學報 2017年1期
張振文,林立銘,余厚美,李開綿,陳松筆
(中國熱帶農(nóng)業(yè)科學院 熱帶作物品種資源研究所/農(nóng)業(yè)部木薯種質(zhì)資源保護與利用重點實驗室/國家薯類加工技術(shù)研發(fā)分中心,海南 儋州 571737)
溫度對木薯葉片葉綠素熒光及Rubisco酶的影響
張振文,林立銘,余厚美,李開綿,陳松筆*
(中國熱帶農(nóng)業(yè)科學院 熱帶作物品種資源研究所/農(nóng)業(yè)部木薯種質(zhì)資源保護與利用重點實驗室/國家薯類加工技術(shù)研發(fā)分中心,海南 儋州 571737)
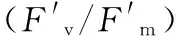
木薯;葉片;葉綠素熒光;溫度;Rubisco
光合生理是淀粉積累和產(chǎn)量形成的生物學基礎(chǔ),其生理生態(tài)過程一直是植物應用基礎(chǔ)研究的一個重要領(lǐng)域。隨著新型的葉綠素熒光動力學技術(shù)的發(fā)展和應用,這種快速、無損傷、能較好地反映植物葉片光合功能的技術(shù),已經(jīng)成為研究植物葉片光合生理過程的重要技術(shù)手段,在植物葉片光合生理和抗逆性研究中得到廣泛的應用[1-4]。植物葉綠色熒光絕大部分是由光合系統(tǒng)Ⅱ天線葉綠素a發(fā)出的,與光合作用過程的各個步驟密切偶聯(lián),任何一步的變化都會影響到光合系統(tǒng)Ⅱ(PSⅡ),從而引起熒光變化,進而反映了光合作用過程的變化,是光合作用的有效探針[5]。研究表明,熒光參數(shù)與溫度關(guān)系密切,在低溫脅迫下木薯葉片葉綠素含量和熒光參數(shù)出現(xiàn)不同程度的差異[6-8],且不同品種間存在顯著性差異[9],同時誘導葉綠素熒光還可以有效地降低葉片花葉病發(fā)病程度[10]。同樣,作為木薯葉光合作用中關(guān)鍵酶之一的1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco),其活性和表達量也受溫度的影響,與凈光合速率基本保持一致[11],但在高溫脅迫下不同品種間的葉片葉綠素熒光參數(shù)和光合系統(tǒng)中所需Rubisco的表達情況如何,尚不清楚。
為此,本文以木薯盆栽苗為研究對象,探討了葉片葉綠素熒光在不同溫度條件下的變化,分析不同品種在不同溫度下其光合生理和Rubisco酶表達情況,有利于了解木薯苗期葉片對溫度的耐受能力,為木薯高產(chǎn)栽培技術(shù)提供理論支撐。
1 材料與方法
1.1 實驗材料
實驗材料來源于中國熱帶農(nóng)業(yè)科學院熱帶作物品種資源研究所木薯種質(zhì)資源圃保存的3個品種,分別是華南5(ManihotesculentaCrantz. cv. M. South China 205,SC205)、華南8號(ManihotesculentaCrantz. cv. M. South China 8,SC8)和從泰國引進的KU50,以及從國際熱帶農(nóng)業(yè)中心(CIAT)引進的野生木薯(ManihotglazioviiMueller von Argau,Wild)。
1.2 實驗方法
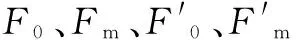
Fv=Fm-F0
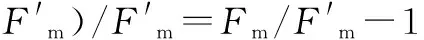

1.2.2 蛋白質(zhì)印跡 塊根總蛋白提取參考莊盈婷[12]的丙酮沉淀法,抗體使用試劑盒(Agrisera公司生產(chǎn))。
1.3 統(tǒng)計與分析
采用Sigmaplot 11.0軟件制作圖表,利用Gel-Pro Analyzer 4.0分析蛋白質(zhì)印跡,并利用SAS 8.1軟件進行方差分析。
2 結(jié)果與分析
2.1 光化學量子效率
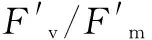

圖1 不同木薯品種最大光量子產(chǎn)量比較
然而,從實際光化學量子效率PhiPS2(ΦPSⅡ)來看,KU50和Wild在不同溫度條件下,其PhiPS2均顯著高于栽培中的SC205和SC8(圖2),且達到極顯著差異(表1),可見國內(nèi)主要栽培品種SC205和SC8的實際光能利用率不高,遠低于KU50和Wild。此外,在25 ℃條件下,SC205和SC8葉片的PhiPS2極顯著高于其他溫度條件,說明25 ℃是該品種木薯苗期葉片光合作用較為適應的溫度,適應我國木薯種植地的氣溫條件。
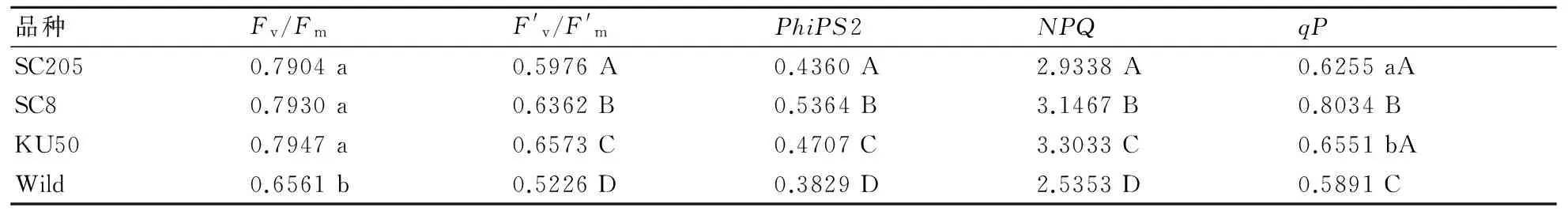
表1 不同品種木薯苗期葉片熒光參數(shù)方差分析

表2 不同溫度木薯苗期葉片熒光參數(shù)方差分析
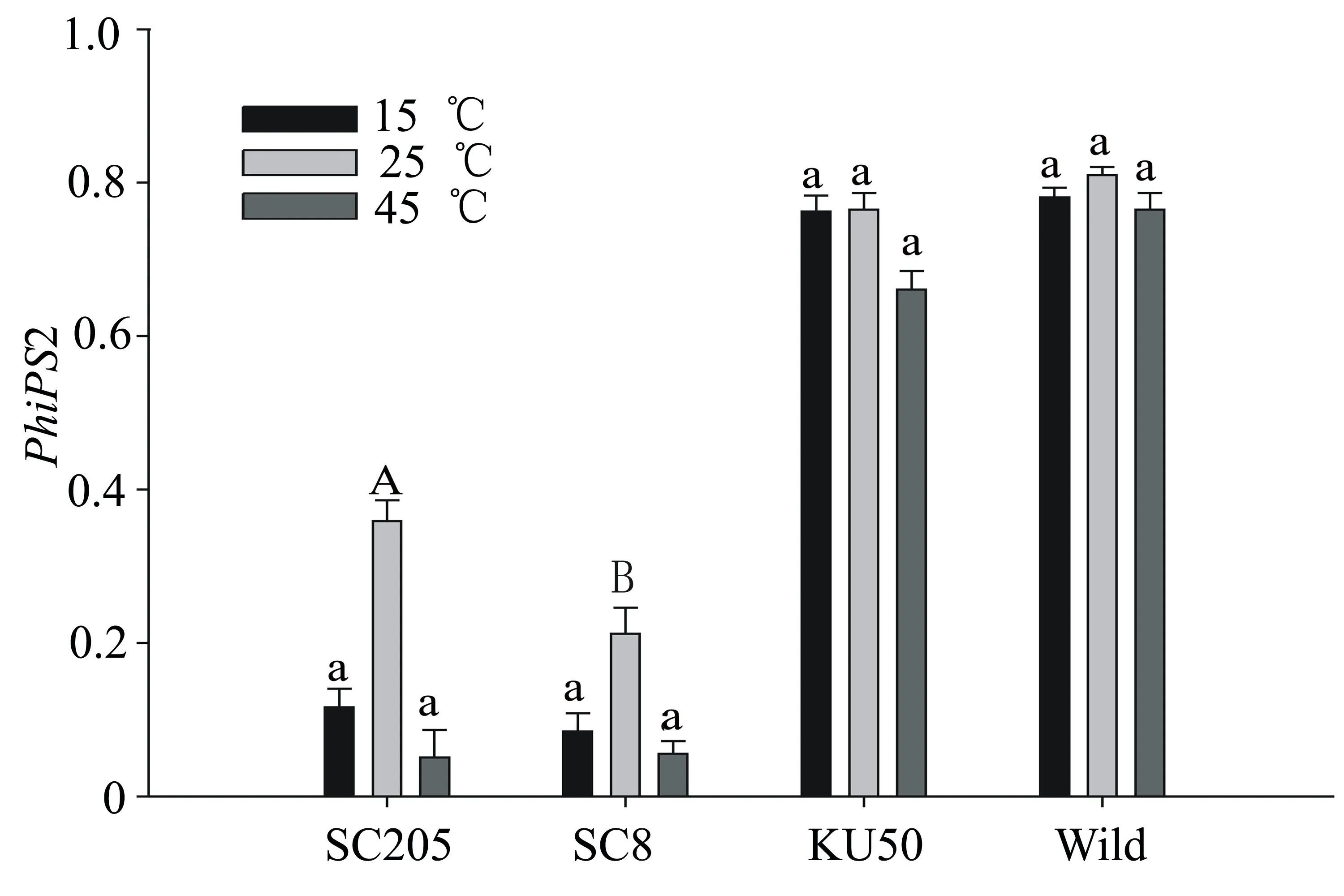
圖2 不同木薯品種實際光量子產(chǎn)量比較
2.2 光化學淬滅系數(shù)
光化學淬滅(qP)反映PSⅡ天線色素分子吸收光能后,用于光化學電子傳遞的份額,因此也反映了色素天線吸收的光能用于光合電子傳遞的變化,由光合作用引起的熒光淬滅,反映了光合活性的高低[14];同時,qP也反映PhiPS2初級電子受體(QA)氧化還原狀態(tài)的變化。要保持高的光化學淬滅,就要使PSⅡ反應中心處于開放狀態(tài)。本研究發(fā)現(xiàn),qP在KU50和Wild的葉片中極顯著高于SC205和SC8(圖3-A),與PhiPS2的變化基本一致,且不同溫度條件下qP差異不顯著,說明KU50和Wild對環(huán)境適應性較強。SC205和SC8這2個品種卻在25 ℃時才表現(xiàn)出較高的光化學活性,并在45 ℃時達到最低水平(圖3-A),說明高溫對這2個品種苗期葉片光合十分不利,處于光抑制狀態(tài)。
在非光化學猝滅(NPQ)系數(shù)中,NPQ反映了由于內(nèi)囊體膜內(nèi)基質(zhì)酸化而引起的內(nèi)囊體膜能化的能量耗散,能量耗散主要發(fā)生在天線色素上,跨類囊體膜質(zhì)子梯度的形成是引起NPQ變化的必要條件,反映了植物過剩光能。本研究發(fā)現(xiàn),25 ℃下,葉片剩余的光能較多,而45 ℃時相對較少,說明葉片吸收后的光能未能被充分利用,其中KU50和Wild葉片的NPQ在不同溫度條件下均極顯著高于SC205和SC8(圖3-B,表1),說明這2個品種的光補償點低于常規(guī)栽培品種SC205和SC8。

圖3 光化學和非光化學淬滅系數(shù)
2.3 Rubisco差異表達分析
一直以來,核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)是探討作物光合機理的重要限速酶,存在于葉綠體基質(zhì)中,是C3植物光合作用的關(guān)鍵羧化酶,催化CO2固定和參與光呼吸。研究表明:增強Rubisco酶活力是提高光合作用的重要途徑,而環(huán)境溫度是影響其活化狀態(tài)的主要原因[15]。木薯是一種高光效的熱帶作物,含有一定的量Rubisco酶。本研究表明,隨著溫度的升高4個品種對溫度的響應存在差異。在45 ℃高溫下,KU50和野生品種葉片中Rubisco酶具有較高的表達,且野生品種(Wild)在45 ℃條件下,其表達量極顯著高于25 ℃,但栽培種SC8和SC205在45 ℃時Rubisco表達量最低;在15 ℃低溫下,SC205的Rubisco表達量最高,并呈現(xiàn)隨著溫度升高而降低的趨勢(圖4)。
3 結(jié)論與討論
3.1 溫度對熒光參數(shù)的影響
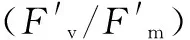

圖4 不同溫度下木薯葉片Rubisco差異表達分析
3.2 溫度對Rubsico酶活性的影響
Rubisco酶是光合碳同化作用的關(guān)鍵酶,是調(diào)節(jié)光合和光呼吸,從而決定凈光合的一個關(guān)鍵酶,具有熱不穩(wěn)定性,其羧化、加氧活性受環(huán)境溫度影響十分明顯[20]。多數(shù)研究表明,影響植物光合作用的各種生態(tài)因子都通過Rubisco而起作用[21]。本研究表明,隨著溫度的升高,野生品種葉片中Rubisco表達的量增加,說明野生種適應高溫條件,這與野生種高溫條件葉片的高熒光效率相一致。對于栽培種,一般認為光合最適溫度為25 ℃,但3個栽培品種僅有SC8的Rubisco表達量在25 ℃時達最高,而KU50甚至出現(xiàn)最低,這很可能是不同品種Rubisco活化酶(RCA)對溫度的響應并不一致[21]。
由此可知,木薯葉片葉綠素熒光效率與溫度和品種密切相關(guān),而葉片葉綠素熒光效率可以間接地反映RCA含量的變化情況,也間接反映出Rubisco酶的活性。然而,木薯葉片中還存在另一個碳同化的途徑,不同溫度條件下,這2個碳同化途徑是如何協(xié)同調(diào)節(jié)木薯葉片的光合作用有待進一步研究。
[1] 李曉,馮偉,曾曉春.葉綠素熒光分析技術(shù)及應用進展[J].西北植物學報,2006,26(10):2186-2196.
[2] 湯文仲,李信書,黃海燕,等.不同光強和溫度對長石莼(緣管滸苔)光合作用和葉綠素熒光參數(shù)的影響[J].水產(chǎn)學報,2009,33(5):762-769.
[3] 吳韓英,壽森炎,朱祝軍,等.高溫脅迫對甜椒光合作用和葉綠素熒光的影響[J].園藝學報,2001,28(6):517-521.
[4] 胡文海,喻景權(quán).低溫弱光對番茄葉片光合作用和葉綠素熒光參數(shù)的影響[J].園藝學報,2001,28(1):41-46.
[5] Govindjee, George C P. Chlorophyll a fluorescence: a signature of photosynthesis[J]. Photosynthesis Research, 2005, 83(1): 101-105.
[6] 陳霆,李開綿,安飛飛,等.華南系列木薯葉綠素熒光參數(shù)及光系統(tǒng)Ⅱ:相關(guān)蛋白表達水平分析[J].江西農(nóng)業(yè)大學學報,2014,36(3):514-519.
[7] 孟凡超.木薯抗寒性的表達譜分析[D].海口:海南大學,2012.
[8] Oyetunji O J, Ekanayake I J, Osonubi O, et al. Chlorophyll fluorescence analysis for assessing water deficit and arbuscular mycorrhizal fungi (AMF) inoculation in Cassava (ManihotesculentaCrantz)[J]. Advances in Biological Research, 2007, 1 (3/4): 108-117.
[9] 張振文,黎良賢,簡純平,等.不同溫度對不同木薯品種盆栽苗葉片光合特性的影響[J].熱帶作物學報,2015,36(1):1-7.
[10] Anderson B, Eghan M J, Asarebediako E. Violet diode laser-induced chlorophyll fluorescence: a tool for assessing mosaic disease severity in cassava (ManihotesculentaCrantz) cultivars [J]. Environmental Technology, 2012, 33(3): 367-372.
[11] 姜振升,孫曉琦,艾希珍,等.低溫弱光對黃瓜幼苗Rubisco與Rubisco活化酶的影響[J].應用生態(tài)學報,2010,21(8):2045-2050.
[12] 莊盈婷.木薯葉片和葉綠體應答鹽脅迫的比較蛋白質(zhì)組學研究[D].海口:海南大學,2014.
[13] Powles S B, Berry J A, Bj?rkman O. Interaction between light and chilly temperature on the inhibition of photosynthesis in chilling-sensitive plants[J]. Plant Cell & Environment, 2006, 6(2): 117-123.
[14] 張守仁.葉綠素熒光動力學參數(shù)的意義及討論[J].植物學通報,1999,16(4):444-448.
[15] 李衛(wèi)芳.小麥Rubisco和Rubisco活化酶對光合作用的影響[D].揚州:揚州大學,2002.
[16] Papageorgiou G C, Govindjee. Photosystem Ⅱ fluorescence: slow changes-scaling from the past[J]. Journal of Photochemistry & Photobiology B Biology, 2011, 104(1/2): 258-270.
[17] 張順堂,張桂蓮,陳立云,等.高溫脅迫對水稻劍葉凈光合速率和葉綠素熒光參數(shù)的影響[J].中國水稻科學,2011,25(3):335-338.
[18] 王瑞華,郭峰,李海峰,等.高溫脅迫對不同葡萄品種葉綠素熒光參數(shù)的影響[J].中外葡萄與葡萄酒,2014(1):33-35.
[19] 張振文,李開綿,葉劍秋,等.木薯光合作用特性研究[J].云南農(nóng)業(yè)大學學報:自然科學版,2007,29(6):628-632.
[20] 陳為鈞,趙貴文,顧月華.RubisCO的研究進展[J].生物化學與生物物理進展,1999;26(5):433-436.
[21] 常阿麗,毛曉芳,韓蓉,He-Ne激光和增強UV-B輻射對小麥幼葉葉綠素熒光和Rubisco活化酶的影響[J].西北植物學報,2013,33(9):1823-1829.
(責任編輯:曾小軍)
Effects of Temperature on Chlorophyll Fluorescence Parameters and Rubisco Activity in Cassava Leaves
ZHANG Zhen-wen, LIN Li-ming, YU Hou-mei, LI Kai-mian, CHEN Song-bi
(Institute of Tropical Crop Genetic Resources, Chinese Academy of Tropical Agricultural Sciences / Key Laboratory of Conservation and Utilization of Cassava Genetic Resources, Ministry of Agriculture / National Center of Potato Processing Technique Research and Development, Danzhou 571737, China)

Cassava; Leaf; Chlorophyll fluorescence; Temperature; Rubisco
2016-07-05 基金項目:國家自然科學基金項目(No.31371684);現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系(CARS-12);農(nóng)業(yè)部木薯種質(zhì)資源創(chuàng)新利用重點實驗室 共同資助項目。
張振文,副研究員,主要從事木薯光合和采后生理研究。*通訊作者:陳松筆。
S533
A
1001-8581(2017)01-0001-05
