放牧調控對短花針茅種群年齡及葉性狀的影響
2017-02-15 08:21:11劉文亭衛智軍呂世杰王天樂張爽海松
草業學報 2017年1期
關鍵詞:植物
劉文亭,衛智軍*,呂世杰,王天樂,張爽,海松
(1.內蒙古農業大學草原與資源環境學院, 內蒙古 呼和浩特 010019; 2.內蒙古農業大學理學院, 內蒙古 呼和浩特 010018;3.內蒙古蘇尼特右旗地震局, 內蒙古 蘇尼特右旗 011200)
放牧調控對短花針茅種群年齡及葉性狀的影響
劉文亭1,衛智軍1*,呂世杰2,王天樂1,張爽1,海松3
(1.內蒙古農業大學草原與資源環境學院, 內蒙古 呼和浩特 010019; 2.內蒙古農業大學理學院, 內蒙古 呼和浩特 010018;3.內蒙古蘇尼特右旗地震局, 內蒙古 蘇尼特右旗 011200)
草食動物對植物種群結構和功能的調控機制是放牧生態學研究的重要內容。本研究以內蒙古錫林郭勒盟蘇尼特右旗荒漠草原的長期放牧控制實驗為平臺,通過研究荒漠草原建群種短花針茅種群年齡動態與其相應葉性狀,以期揭示荒漠草地放牧調控下植物響應機制。結果顯示,1)短花針茅種群基徑小于4 mm部分植株存活數少;適度放牧處理短花針茅各年齡階段頻度與不放牧基本一致,重度放牧處理齡級Ⅲ的頻度和比例分別下降了0.79,0.73;2)短花針茅種群齡級Ⅰ與齡級Ⅱ的短花針茅頻度和比例較高,齡級Ⅲ相對較低,屬于比較穩定的種群;Godron指數顯示短花針茅年齡結構的穩定性重度放牧>適度放牧>不放牧;3)放牧調控下,葉高、葉長、葉干重、葉直立度、葉長寬比為年齡型敏感性狀,自然葉寬、完全葉寬、葉卷曲度、葉干物質含量為年齡型保守性狀;4)相關分析認為自然葉寬和葉干物質含量與各年齡型敏感性狀基本呈協同變化;葉卷曲度與葉干重負相關(P<0.05),與葉直立度呈正相關(齡級Ⅲ、齡級Ⅰ)。研究認為,短花針茅種群在亞穩定下,通過控制種群年齡及葉性狀,來完成其生活史對放牧調控的適應。
種群年齡; 葉性狀; 放牧; 短花針茅; 荒漠草地
種群結構是植物種群生態學研究的重要內容[1-2],揭示草地群落優勢種結構與動態對闡明草地生態系統群落的形成、維持和演替規律與種群特征、更新和穩定性具有重要意義[3]。種群的年齡結構不僅反映了植物的更新和個體發育狀態,清晰地描述了不同年齡組分個體在種群內的地位和配置情況,而且體現了植物種群在復雜生境中所表現出的行為策略。研究表明,穩定的種群內部各年齡基本呈“倒J”型分布[3],且與植株生長、存活和再生潛力緊密相連。短花針茅(Stipabreviflora)是亞洲中部暖溫性草原的主要建群種,研究短花針茅種群結構和穩定性對放牧的響應,對揭示荒漠草地資源利用與植物響應機制有很強的代表性。
草食動物作為草地植物組成與多樣性的管理者,其對植物種群的影響直接體現于植株的表型特征[4]。研究認為,植物株高對放牧的反應最為直接和敏感[5],被認為是最有效的指標,并伴隨葉片變短變窄、節間縮短、枝葉硬挺、叢幅變小、種子縮小、花期提前和根系分布淺層化等性狀的改變[6-7]。Cruz等[8]發現長年放牧植物葉干物質含量顯著降低,但比葉面積增大,來進一步提高其光合能力。可見,植物在不同生境下,各器官性狀會根據其功能需求,來實現植物自身可承受的資源配置策略譜[9-10]。
植物種群特征隨植株年齡變化的風險投資和物質分配是植物權衡策略的重要內容[11-12]。Tilman[13]認為,在植物生長早期,葉片會加大同化器官的成本投入,如擴展葉面積,來增大光截獲和碳收益的競爭優勢,補償其建成消耗[9]。而家畜喜食幼時的植物[14],顯然,這會對幼苗的生存帶來巨大的生存威脅,那么短花針茅會采取何種手段來維持其生存?而隨植株年齡的累積,植物會經歷更多更為復雜的脅迫。例如在抗機械脅迫和結構建造方面投資更多的生物量,來維持葉存活時間[15]。那么,綿羊在進行采食等行為的過程中,會對其視若無睹嗎?短花針茅又會采取何種策略來生存呢?
草食動物的采食行為是一個非常復雜的過程,在采食過程中,家畜需要在植株個體間重復進行選擇,選擇的結果直接影響植物的存活狀態。長期超載過牧,引發草地生產力持續衰退,生態系統功能衰減。因此,解析放牧對草地生態系統的作用機制,一直是近年來研究的熱點問題[11]。本研究依托于內蒙古錫林郭勒盟蘇尼特右旗荒漠草地建立的長期放牧平臺作為模式生態系統,以暖溫性荒漠草地建群種短花針茅為模式植物,試圖從草地現存植株不同年齡階段這一視角,來探討不同放牧方式各生長階段的短花針茅數量如何變化,其種群穩定性如何,與之相應的短花針茅葉性狀對放牧調控采取何種權衡變化,這將為制定干旱區荒漠草地的放牧管理提供理論基礎與數據支撐。
1 材料與方法
1.1 研究區概況與樣地描述
研究區位于內蒙古錫林郭勒盟蘇尼特右旗朱日和鎮(112°47′11.2″ E, 42°15′48.7″ N)的牧區。實驗區地形較為平坦,有明顯的鈣積層,大部分分布在地下10~80 cm之間,以亞洲中部荒漠草原種占主導地位,植被草層低矮,一般10~35 cm;植被稀疏,短花針茅為建群種,它在群落外貌和群落環境中起決定作用。優勢種為無芒隱子草(Cleistogenessongorica)和堿韭(Alliumpolyrhizum),伴生有寸草苔(Carexduriuscula)、戈壁天門冬(Asparagusgobicus)、狹葉錦雞兒(Caraganastenophylla)、木地膚(Kochiaprostrata)、乳白花黃芪(Astragalusgulactites)、銀灰旋花(Convolvulusammannii)、茵陳蒿(Artemisiacapillaries)、阿爾泰狗娃花(Heteropappusaltaicus)等。
本研究所選用的試驗樣地位于連續的同一地段,地勢平坦,環境相對均勻,有效地控制了本底和空間異質性的差異。放牧試驗于2010年開始,每年5月開始放牧,10月底終止,期間采用連續放牧方式,晚上羊群在樣地不歸牧,不進行補飼。本研究設有3個放牧處理,依次為適度放牧、重度放牧和不放牧處理,每個處理均設有3次重復,每個實驗小區面積約為2.60 hm2。適度放牧處理和重度放牧處理載畜率分別為0.96和1.54只羊/(hm2·年)。放牧的草食動物為“蘇尼特羊”。放牧區綿羊的健康狀況、個體大小、體重、性別基本一致。
1.2 取樣方法與計算
于2015年8月進行野外取樣,在每個試驗小區,隨機設置5個1 m×1 m的樣方,每個1 m×1 m樣方內,使用電子游標卡尺測量所有現存的短花針茅基徑,對應隨機選取該株10片葉片(不足10片的株叢全部測量),測量其葉高(自然狀態下葉片最高點距離地面的垂直高度,leaf height,LH)、自然葉寬(葉片自然狀態的寬度,natural leaf width,NLW),將每個植株用剪刀齊地剪起、編號,帶回室內陰涼處處理。于12 h內用去離子水快速沖洗干凈,吸干表面水分,稱取采集的短花針茅濕重(leaf fresh weight,LFW),并測量葉片完全寬度(葉片展平時的寬度,total leaf width,TLW)、葉長(葉片拉直時的長度,leaf length,LL),待完成后,將新鮮材料置于105 ℃烘箱中殺青10 min,將葉片置于75 ℃干燥箱中烘干至恒重,稱量樣品干重(leaf dry weight,LDW)。通過公式(1~4)計算出短花針茅葉片的卷曲度(leaf rolling index,LRI)、直立度(leaf erection index,LEI)、干物質含量(leaf dry matter content,LDMC)、長寬比(leaf length/leaf width,L/W):
(1)
(2)
(3)
(4)
1.3 數據處理與統計分析
根據短花針茅株叢實際基徑(d)大小,同時結合已有研究[16],將短花針茅劃分為11個生長層次(d≤4 mm, 4 mm
采用Godron穩定性測定方法評價短花針茅種群的穩定性。以試驗小區所有樣方為單位,將種群中不同生長層次的頻度由大到小排序,把頻度換算成相對頻度,按相對頻度由大到小的順序逐步累積相加,與此同時,逐一對應累積短花針茅生長層次的倒數,采用模擬散點所得一元二次平滑曲線的方程與坐標軸第一象限的曲線y=1-x連接的對角直線來判斷累積短花針茅生長層次倒數與累積相對頻度比值的交點坐標,0.2/0.8是群落的穩定點,交點坐標距離越接近0.2/0.8,種群就越穩定[18]。
根據樣方內短花針茅不同生長層次植株密度來計算各生長層次短花針茅種群比例及頻度。采用SPSS 19.0進行雙因素方差分析(two-way ANOVA)檢驗放牧方式和年齡階段及其交互作用對荒漠草原短花針茅葉性狀的影響,并進行多重比較(Duncun)檢驗不同放牧方式不同年齡階段間短花針茅葉功能性狀的顯著性差異。將數據標準化后使用Pearson相關分析來解釋短花針茅不同年齡階段葉性狀間的關系,在0.05和0.01水平進行顯著性檢驗。
2 結果與分析
2.1 短花針茅種群年齡結構特征
2.1.1 短花針茅年齡結構數量特征對放牧方式的響應 短花針茅種群的年齡結構直接體現了不同年齡階段的個體在種群內的比例或配置情況,野外調查結果顯示(圖1),齡級Ⅰ與齡級Ⅱ的短花針茅頻度和權重整體較高,齡級Ⅲ相對較低,年齡結構基本呈金字塔型,即幼苗儲備豐富,成年植株生長狀況較好,屬于比較穩定的種群。具體而言,短花針茅種群基徑小于4 mm部分植株存活數少;適度放牧和不放牧處理各年齡階段短花針茅皆有分布,且適度放牧草地各年齡階段短花針茅頻度與不放牧草地短花針茅頻度基本一致,重度放牧短花針茅齡級Ⅰ頻度較不放牧下降了0.29,齡級Ⅱ頻度下降了0.41,齡級Ⅲ下降了0.79。各草地短花針茅年齡結構基本呈單峰曲線,基徑在4~41 mm這一范圍比值最高,重度放牧草地短花針茅齡級Ⅰ、齡級Ⅱ比例下降了0.06, 0.08,但齡級Ⅲ下降了0.73,適度放牧草地齡級Ⅰ、齡級Ⅱ短花針茅與不放牧草地短花針茅比例變幅較小(不超過0.12)。
2.1.2 短花針茅年齡結構穩定性對放牧方式的響應 本研究采用Godron指數來評價短花針茅年齡結構的穩定性。將短花針茅不同年齡階段累積相對頻度與總個體數的倒數一一對應,繪制個體數倒數累積-相對頻度累積的散點圖(圖2),之后完成平滑曲線的模擬,其結果見表1。重度放牧草地、適度放牧草地、不放牧草地相應的交點坐標分別為(0.27, 0.72)、(0.29, 0.71)、(0.32, 0.68)。離穩定點(0.2, 0.8)距離由近到遠分別是重度放牧草地0.11、適度放牧草地0.13、不放牧草地0.17。
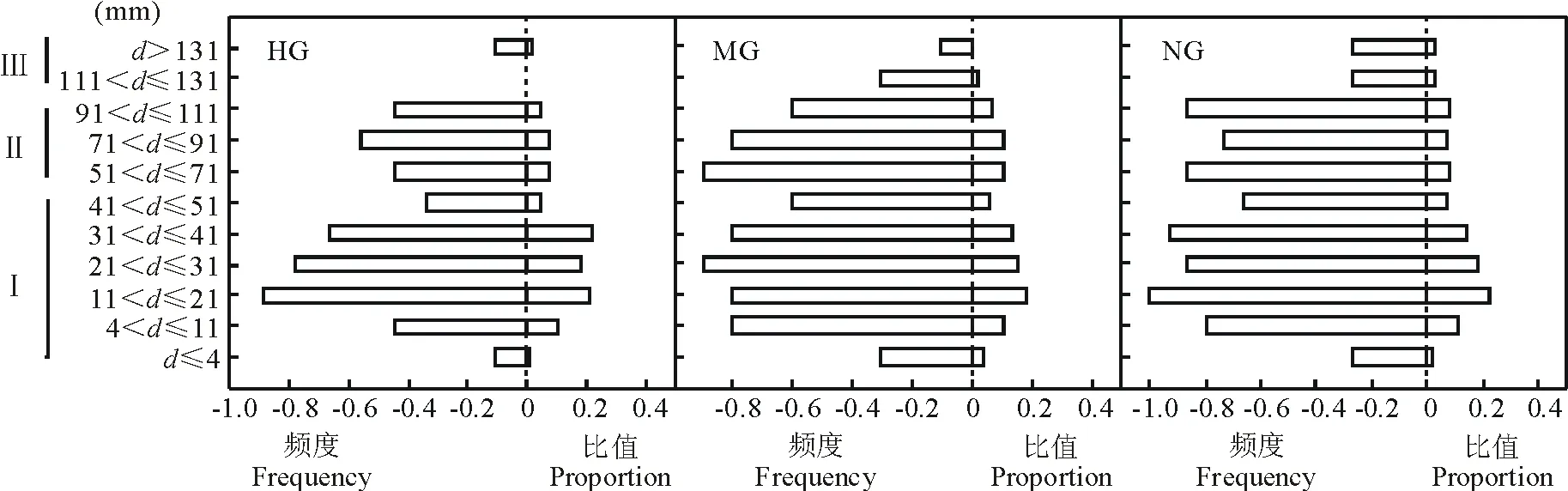
圖1 短花針茅種群組成對放牧方式的響應特征Fig.1 Response characteristics of S. breviflora age composition to the grazing systems HG: 重度放牧Heavy grazing; MG: 適度放牧Moderate grazing; NG: 不放牧No grazing.
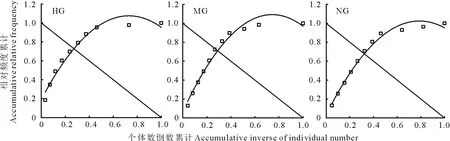
圖2 短花針茅年齡結構Godron圖Fig.2 Godron scatter graphs of S. breviflora age composition in desert grassland
2.2 不同放牧方式短花針茅異齡葉性狀
雙因素方差分析表明(表2),不同放牧方式短花針茅葉性狀均存在顯著性差異(P<0.05),從年齡結構分析,葉高、葉長、葉干重、葉直立度、葉長寬比亦存在顯著性差異(P<0.05);自然葉寬、完全葉寬、葉卷曲度、葉干物質含量無明顯差異,但放牧方式和年齡結構的交互作用結果顯示(表2),自然葉寬、完全葉寬、葉高、葉長、葉干重、葉直立度和葉干物質含量存在顯著性差異(P<0.05)。因此,在放牧調控下可將短花針茅葉性狀按年齡響應模式分為2類:年齡型敏感性狀和年齡型保守性狀。在年齡敏感性狀中(圖3),葉高、葉長、葉干重、葉直立度、葉長寬比基本為適度放牧草地>不放牧與重度放牧草地,其葉長、葉干重、葉長寬比均為齡級Ⅲ>齡級Ⅱ>齡級Ⅰ。

表1 短花針茅年齡結構穩定性分析結果Table 1 Stability of S. breviflora age composition in desert grassland
y:相對頻度累積;x:個體數倒數累計。
y: Accumulative relative frequency;x:Accumulative inverse of individual number.
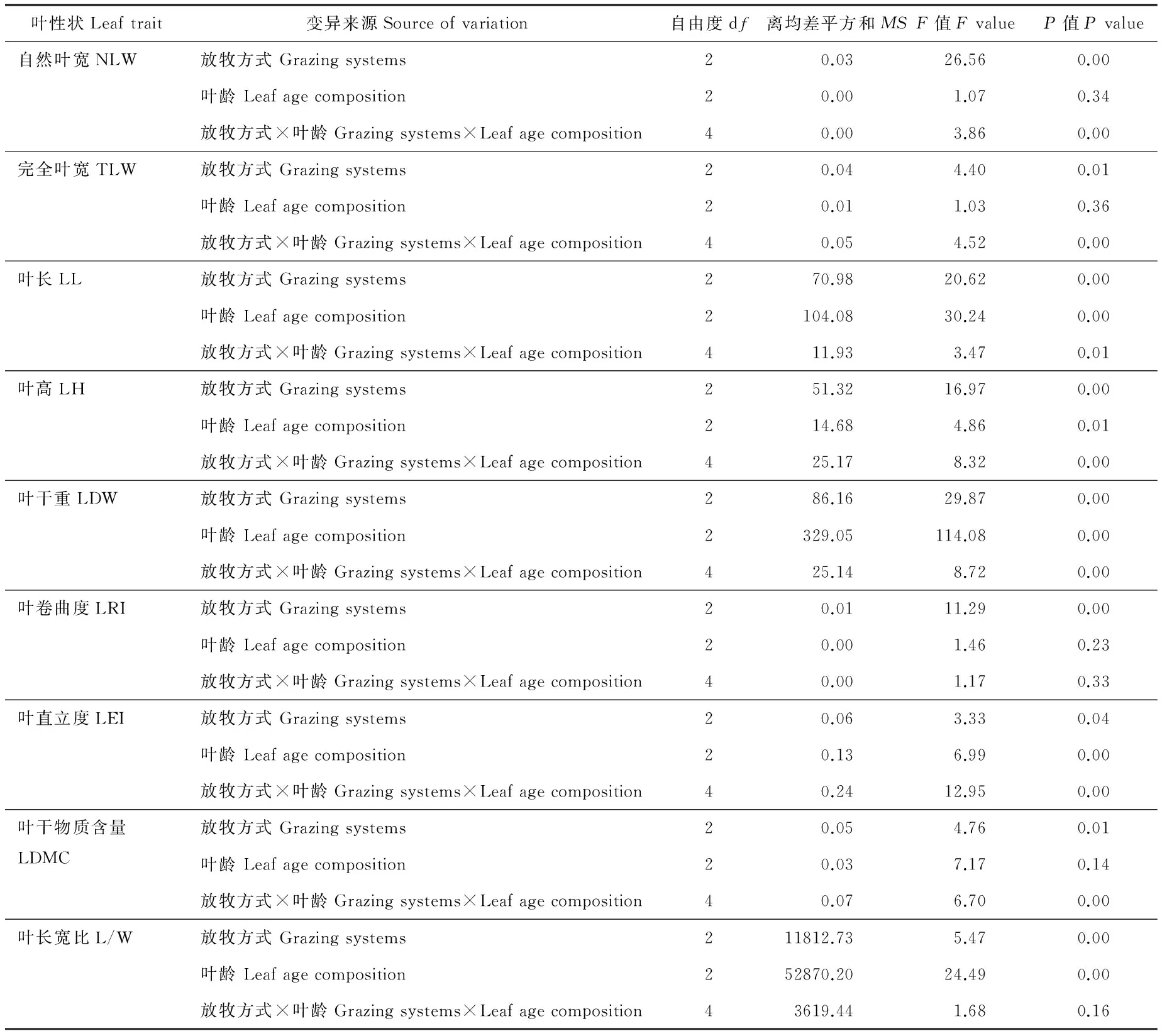
表2 放牧方式與葉齡對短花針茅葉性狀影響的雙因素方差分析Table 2 Two-way ANOVA for plant traits affected by grazing system and age composition
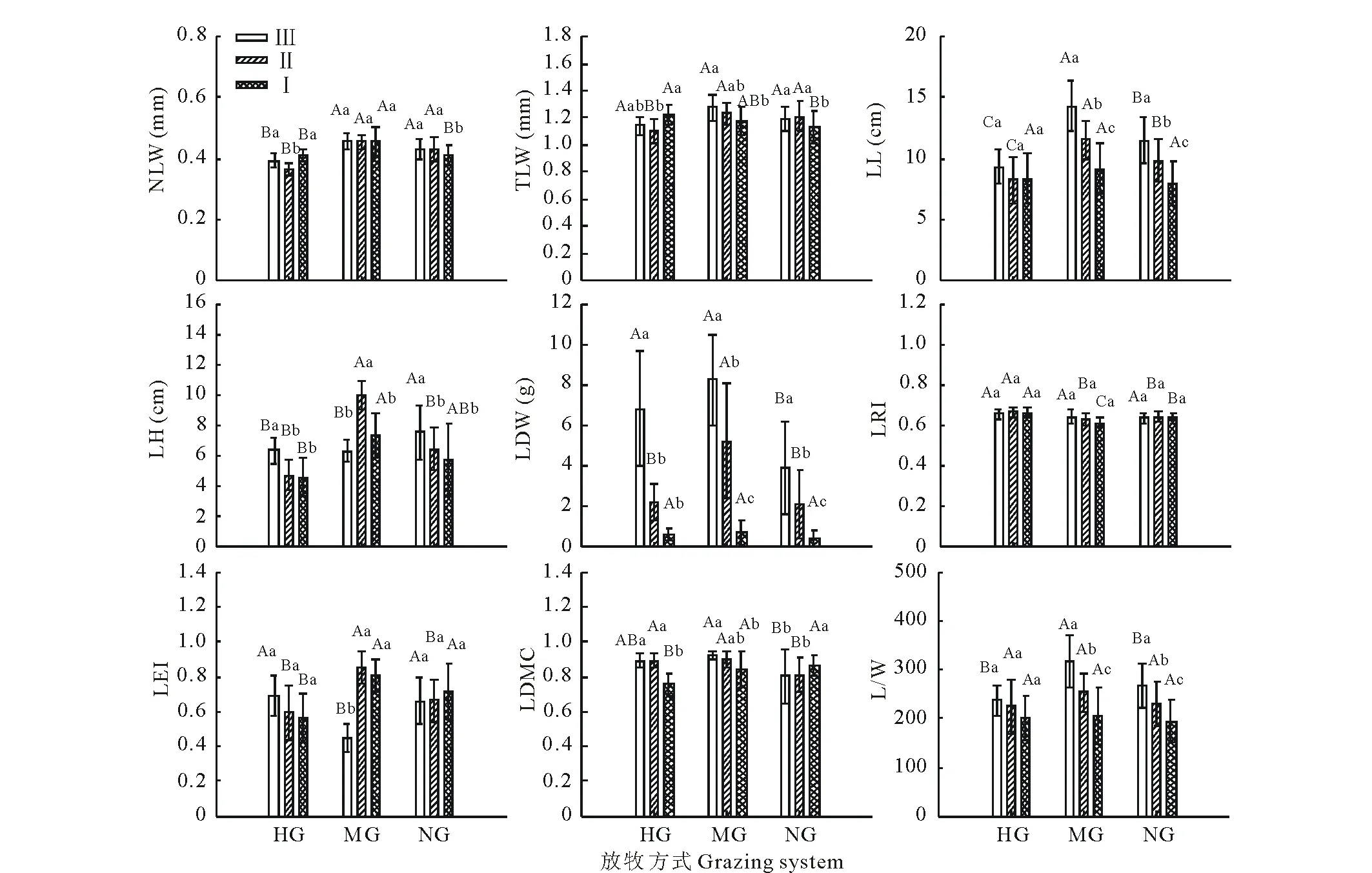
圖3 不同放牧方式不同葉齡短花針茅的葉性狀(平均值±標準誤差)Fig.3 Plant traits of S. breviflora in different grazing system and different age composition in desert grassland (mean±SE) 不同大寫字母表示不同放牧方式葉性狀存在顯著差異,不同小寫字母表示不同葉齡葉性狀存在顯著差異。Different capital letters indicate significant differences in grazing systems, and different lowercase letters indicate significant differences in the age compositions.
2.3 短花針茅葉性狀間的關系
植物葉性狀間的關系體現了植物一系列彼此聯系、協調變化且有規律地資源權衡策略。為了進一步揭示年齡型敏感性狀和年齡型保守性狀間的關系,相關分析結果顯示(圖4),短花針茅自然葉寬和葉干物質含量與各年齡型敏感性狀基本呈正相關,其中自然葉寬與葉長、葉干重(齡級Ⅲ、齡級Ⅱ)顯著正相關(P<0.05),葉干物質含量僅與葉直立度、葉高(齡級Ⅰ)呈顯著正相關。各年齡型敏感性狀與葉卷曲度基本呈負相關(圖4C),其中與各齡葉干重呈顯著負相關(P<0.05),與葉直立度呈正相關(齡級Ⅲ、齡級Ⅰ)。葉全寬與各年齡型敏感性狀則無顯著相關關系。
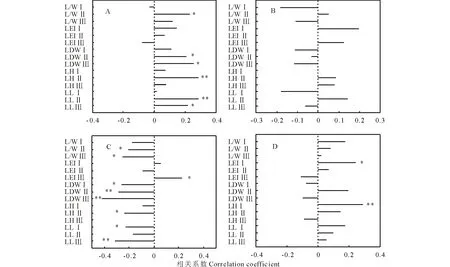
圖4 短花針茅年齡型保守性狀和年齡型敏感性狀的相關分析Fig.4 Correlation analysis of S. breviflora age inert traits and different age sensitive traits A: 自然葉寬NLW; B: 完全葉寬TLW; C: 葉卷曲度LRI; D: 葉干物質含量LDMC. *, P<0.05; **, P<0.01.
3 討論與結論
3.1 不同放牧方式下短花針茅種群結構特征
種群的年齡結構是指不同年齡組的個體在種群內的比例或配置情況,一個給定的生物個體不僅反映了其實際年齡特征,同時還反映了該個體的發育狀態[16]。短花針茅作為亞洲中部偏暖氣候區域草原亞區荒漠草原帶的建群種,其個體存活數量及年齡階段對維持群落結構及地區的畜牧業生產具有重要作用。從短花針茅種群的年齡結構來看,短花針茅種群的年齡結構整體呈金字塔形,處于相對平衡的狀態。荒漠草地各年齡級短花針茅數量特征基本一致,但d≤4 mm階段短花針茅存活的數量最少(圖1),說明若其幼苗“挺過”這一瓶頸期,短花針茅的數量特征則明顯上升。產生這種現象的可能原因是:在生境惡劣且異質性較高的荒漠草地,較年長的植株為了規避死亡風險,故其產生許多幼苗分布在年長植株周圍[12]。這樣,該植株被家畜采食的概率即為年長植株和所有幼苗個體死亡概率的乘積。由于植物的被采食導致死亡的概率小于1,理論上成年植株產生幼苗越多,短花針茅越有可能采取這種平攤風險的策略,來降低個體被采食的風險。此外,荒漠草原環境條件惡劣,幼苗的生存率遠小于成年植株[19],致使短花針茅種群現存幼苗存活率低的現象。
已有研究認為,過度放牧會導致年長針茅株叢的破碎化[20],使其破碎化為幾個較小的株叢,致使重度放牧草地齡級Ⅲ部分缺失(圖1)。針茅屬植物屬于多年生叢生禾草,在相同的環境條件下,短花針茅基徑大小和年齡特征對環境等條件的反應規律基本一致[12],那么,年長植株的破碎化應導致青年植株數量的上升[20]。本研究結果顯示,重度放牧處理使各齡級短花針茅的頻度和權重均下降。這是因為重度放牧處理中,牧畜的過度踐踏改變了土壤表層環境[21],使土壤表層變得緊實,通透性變差,降低了土壤滲透速率,影響了根系的生長[22];加之植物地上部分被家畜反復啃食,使株叢生活力減弱,生長發育受抑制[20],大多數短花針茅的植株不能抽穗、開花和結實,使開花結實株叢的密度相應減少,致使各齡級短花針茅的頻度和權重下降。也有研究認為,動物過度干擾,植物的誘導抵抗會顯著降低動物對幼嫩株叢的傾向性取食[23]。
穩定性指數分析表明,重度放牧草地短花針茅種群穩定性最高,適度放牧草地次之,不放牧草地最低(圖2)。研究認為,放牧條件會改變草地物種間的競爭強度,使一些在競爭上占劣勢的植物種因競爭而被排除[24],加之短花針茅荒漠草原物種多樣性隨放牧梯度下降而上升,短花針茅重要值不隨放牧壓力顯著改變[25],致使短花針茅對現有資源享有絕對統治地位,種群穩定性最高;相反,不放牧處理草地物種多樣性最高,使之加劇了其種內與種間競爭,物種的資源利用競爭性較強,導致單個種群的穩定性降低。
3.2 不同放牧方式下短花針茅葉性狀權衡
現存植物的葉性狀是植物權衡外界環境來改變自身的綜合表現[26],也是整個生物策略的最終環節。傳統研究認為,株高因具直觀和便捷的操作性,被認為是最有效的指標,植物個體矮小化是放牧導致草原生態系統結構和功能衰退的重要觸發機制[7]。本研究結果顯示,僅齡級Ⅲ出現隨放牧梯度增加株高(葉高)下降的現象(圖3)。這是因為短花針茅是多年生叢生禾草,在其營養生長旺季植株無明顯莖葉分化,葉高即為株高,葉片卷成長筒狀條形[27],故葉面積的增大意味著一定程度的葉高的增加,而老葉組織密度和厚度的增大使得光合能力的衰退[26, 28],葉片通過繼續增大面積這一策略來進行的光合收益-支出的邊際效應已甚小,繼續投資葉面積顯然不符合葉經濟譜原理[9]。這一定程度證明了短花針茅葉性狀對放牧的響應伴隨種群年齡的權衡。可見,單一指標作為衡量草地放牧管理有效指標并不能完全指示和表征植被的異化,因此種群的年齡結構在放牧管理中應當被充分考慮。
本研究結果顯示,短花針茅葉性狀存在年齡響應模式(表2),葉高、葉長、葉干重、葉直立度、葉長寬比為年齡型敏感性狀,而自然葉寬、完全葉寬、葉卷曲度、葉干物質含量為年齡型保守性狀,可見,在放牧干擾下,短花針茅葉性狀具有不同的適應對策。一般來說,在生境脅迫下,植物性狀之間具有明顯的協同變化現象[11],而這種協同變化現象也表現在放牧調控下種群不同年齡響應模式葉性狀間(圖4),如短花針茅自然葉寬和葉干物質含量與各年齡型敏感性狀基本呈正相關,其中自然葉寬與葉長、葉干重(齡級Ⅲ、齡級Ⅱ)顯著正相關(P<0.05),葉干物質含量僅與葉挺立度、葉高(齡級Ⅰ)呈顯著正相關。雖然植物對放牧調控存在協同變化,但亦有部分結構性性狀不遵從上述規律,如短花針茅葉卷曲度。短花針茅葉片呈針狀并向內卷曲,極少數縱折、扁平,研究認為卷葉通過改變其葉片的電導率和有效葉面積來減少其攔截的輻射量及降低蒸騰作用,進而有效防止水分的喪失[29-30]。同時葉片卷曲程度的增加可以有效地增強葉片直立程度,減少披垂現象的發生,起到抵抗草地多風、動物踐踏等作用。與此相對的,葉生物量的積累會降低短花針茅的直立程度,這就解釋短花針茅葉卷曲度與各齡葉干重呈顯著負相關。這一定程度說明干旱脅迫是限制荒漠草地植物生長的另一因素,植物的多重生活策略是其克服環境脅迫的重要途徑。
綜上分析得出,1)荒漠草地短花針茅種群在不同放牧強度下處于相對穩定狀態,齡級Ⅰ與齡級Ⅱ的短花針茅頻度和權重整體較高,齡級Ⅲ相對較低,重度放牧降低了齡級Ⅲ數量;種群穩定性呈重度放牧草地>適度放牧草地>不放牧草地;2)葉性狀對放牧的響應應充分考慮植株多性狀間的權衡和其年齡狀態;3)短花針茅葉高、葉長、葉干重、葉直立度、葉長寬比為年齡型敏感性狀,自然葉寬、完全葉寬、葉卷曲度、葉干物質含量為年齡型保守性狀。
References:
[1] Boopathi N M, Hoffmann L V. Genetic Diversity, Erosion, and Population Structure in Cotton Genetic Resources[M]. Switzerland: Springer International Publishing, 2016.
[2] Sampaio-E-Silva T A, Tiberio F C S, Dodonov P,etal. Differences in allometry and population structure between native and invasive populations of a tropical tree. New Zealand Journal of Botany, 2015, 53(2): 90-102.
[3] Agren J, Zackrisson O. Age and size structure ofPinussylvestrispopulations on mires in central and northern Sweden. Journal of Ecology, 1990, 78(4): 1049-1062.
[4] García C A M, Schellberg J, Ewert F,etal. Response of community-aggregated plant functional traits along grazing gradients: insights from African semi-arid grasslands. Applied Vegetation Science, 2014, 17(3): 470-481.
[5] Díaz S, Lavorel S, McIntyre S,etal. Plant trait responses to grazing-a global synthesis. Global Change Biology, 2007, 13(2): 313-341.
[6] Louault F, Pillar V D, Aufrère J,etal. Plant traits and functional types in response to reduced disturbance in a semi-natural grassland. Journal of Vegetation Science, 2005, 16(2): 151-160.
[7] Wang W, Liang C Z, Liu Z L,etal. Analysis of the plant individual behavior during the degradation and restoring succession in steppe community. Chinese Journal of Plant Ecology, 2000, 24(3): 268-274. 王煒, 梁存柱, 劉鐘齡, 等. 草原群落退化與恢復演替中的植物個體行為分析. 植物生態學報, 2000, 24(3): 268-274.
[8] Cruz P, De Quadros F L F, Theau J P,etal. Leaf traits as functional descriptors of the intensity of continuous grazing in native grasslands in the south of Brazil. Rangeland Ecology Management, 2010, 63(3): 350-358.
[9] Wright I J, Reich P B, Westoby M,etal. The worldwide leaf economics spectrum. Nature, 2004, 428: 821-827.
[10] An H. Effects of grazing disturbance on leaf traits and their interrelationships of plants in desert steppe. Chinese Journal of Applied Ecology, 2012, 23(11): 2991-2996. 安慧. 放牧干擾對荒漠草原植物葉性狀及其相互關系的影響. 應用生態學報, 2012, 23(11): 2991-2996.
[11] Li X L, Liu Z Y, Hou X Y,etal. Plant functional traits and their trade-offs in response to grazing: a review. Chinese Bulletin of Botany, 2015, 50(2): 159-170. 李西良, 劉志英, 侯向陽, 等. 放牧對草原植物功能性狀及其權衡關系的調控. 植物學報, 2015, 50(2): 159-170.
[12] Dong M. Cloning Plant Ecology[M]. Beijing: Science Press, 2011. 董鳴. 克隆植物生態學[M]. 北京: 科學出版社, 2011.
[13] Tilman D. Plant Strategies and the Dynamics and Structure of Plant Communities[M]. Princeton: Princeton University Press, 1998.
[14] Ritchie M E, Tilman D, Knops J M H. Herbivore effects on plant and nitrogen dynamics in Oak Savanna. Ecology, 1998, 79(1): 165-177.
[15] Westoby M, Falster D S, Moles A T,etal. Plant ecological strategies: some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 2002, 33: 125-159.
[16] Bai Y F, Xu Z X, Li D X,etal. Study on age and bunch structure of fourStipaspecies in Inner Mongolia Plateau. Chinese Bulletin of Botany, 1999, 41(10): 1125-1131. 白永飛, 許志信, 李德新, 等. 內蒙古高原四種針茅種群年齡與株叢結構的研究. 植物學報, 1999, 41(10): 1125-1131.
[17] Lv S J, Liu H M, Wu Y L,etal. Effects of grazing on spatial distribution relationships between constructive and dominant species inStipabrevifloradesert steppe. Chinese Journal of Applied Ecology, 2014, 25(12): 3469-3474. 呂世杰, 劉紅梅, 吳艷玲, 等. 放牧對短花針茅荒漠草原建群種與優勢種空間分布關系的影響. 應用生態學報, 2014, 25(12): 3469-3474.
[18] Zheng Y R. Comparison of methods for studying stability of forest community. Scientia Silvae Sinicae, 2000, 36(5): 28-32. 鄭元潤. 森林群落穩定性研究方法初探. 林業科學, 2000, 36(5): 28-32.
[19] Lu W H, Zhu J Z, Jin G L. Spatial and temporal pattern ofSerihidiumseedling in small-scale under two gradient degradation. Acta Prataculturae Sinica, 2011, 20(5): 272-277. 魯為華, 朱進忠, 靳瑰麗. 小尺度條件下退化絹蒿種群幼苗更新時空格局. 草業學報, 2011, 20(5): 272-277.
[20] Bai Y F, Li D X, Xu Z X,etal. Growth and reproduction ofStipakryloviipopulation on a grazing gradient. Acta Ecologica Sinica, 1999, 19(4): 479-484. 白永飛, 李德新, 許志信, 等. 牧壓梯度對克氏針茅生長和繁殖的影響. 生態學報, 1999, 19(4): 479-484.
[21] Gao Y Z, Han X G, Wang S P. The effects of grazing on grassland soils. Acta Ecologica Sinica, 2004, 24(4): 790-797. 高英志, 韓興國, 汪詩平. 放牧對草原土壤的影響. 生態學報, 2004, 24(4): 790-797.
[22] Yavuz T, Karadag Y. The effect of fertilization and grazing applications on root length and root biomass of some rangeland grasses. Turkish Journal of Field Crops, 2015, 20(1): 38-42.
[23] Gómez S, Onoda Y, Ossipov V,etal. Systemic induced resistance: a risk-spreading strategy in clonal plant networks. New Phytologist, 2008, 179(4): 1142-1153.
[24] Liu X Y. Effects of Grazing on the Relationships between Plant Diversity and Community Stability on Meadow Steppe[D]. Changchun: Northeast Normal University, 2013. 劉曉媛. 放牧方式對草地植被多樣性與穩定性關系的影響[D]. 長春: 東北師范大學, 2013.
[25] Sun S X, Wei Z J, Lv S J,etal. Characteristics of plant community and its functional groups in desert grassland under effects of seasonal regulation of grazing intensity. Chinese Journal of Ecology, 2013, 32(10): 2703-2710. 孫世賢, 衛智軍, 呂世杰, 等. 放牧強度季節調控下荒漠草原植物群落與功能群特征. 生態學雜志, 2013, 32(10): 2703-2710.
[26] Field C, Mooney H A. Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub. Oecologia, 1983, 56(2/3): 348-355.
[27] The Editorial Committee of Inner Mongolia Flora. Flora Intramongolica[M]. Huhhot: Inner Mongolia Peoples Publishing House, 1989. 內蒙古植物志編輯委員會. 內蒙古植物志[M]. 呼和浩特: 內蒙古人民出版社, 1989.
[28] He J S, Wang X P, Flynn D F B,etal. Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence. Ecology, 2009, 90(10): 2779-2791.
[29] Kadioglu A, Terzi R. A dehydration avoidance mechanism: leaf rolling. The Botanical Review, 2007, 73(4): 290-302.
[30] Price A H, Young E M, Tomos A D. Quantitative trait loci associated with stomatal conductance leaf rolling and heading date mapped in upland rice. New Phytologist, 1997, 137(1): 83-91.
Effect of grazing on Stipa breviflora age dynamics and leaf traits
LIU Wen-Ting1, WEI Zhi-Jun1*, LV Shi-Jie2, WANG Tian-Le1, ZHANG Shuang1, HAI Song3
1.CollegeofGrassland,ResourcesandEnvironmental,InnerMongoliaAgriculturalUniversity,Huhhot010019,China; 2.CollegeofScience,InnerMongoliaAgriculturalUniversity,Huhhot010018,China; 3.SeismologicalBureau,SonidRightBanner011200,China
The regulation mechanism of plant population structure and function by herbivores is an important part of grazing ecology. A long-term grazing study in Xilingol was used to reveal the mechanisms of the regulation of plant responses to grazing through population age dynamics and leaf traits ofStipabreviflorain desert grassland. The result showed that fewS.brevifloraplants with base diameter of less than 4 mm survived. There was little difference between moderate grazing and no grazing. The frequency and proportion of age class Ⅲ plants was reduced 0.79 and 0.73 respectively by heavy grazing.S.breviflorapopulation were relatively stable. Two way ANOVA showed that leaf height, leaf length, leaf dry weight, leaf erectness and leaf length-width ratio were sensitive to grazing in different age classes. By contrast, leaf width, leaf rolling index and leaf dry matter content were not affected by age class. Correlation analysis suggested that leaf width and leaf dry matter content changed synergistically with the age sensitive traits. Leaf rolling index was significantly negatively correlated with leaf dry weight, and was positively related with leaf erectness (e.g. age class Ⅲ and class Ⅰ). The study suggested thatS.brevifloracompleted its life history and adapted to grazing by controlling population age and leaf traits in a metastable state.
population age; leaf trait; grazing;Stipabreviflora; desert grassland
10.11686/cyxb2016277
http://cyxb.lzu.edu.cn
2016-07-14;改回日期:2016-09-07
國家自然科學基金項目(31460126),國家重點基礎研究發展計劃(2014CB138805),內蒙古自治區博士研究生科研創新項目(B20161012906Z)和內蒙古自然科學基金(2015MS0349)資助。
劉文亭(1990-),男,內蒙古呼和浩特人,在讀博士。E-mail: nmgliuwenting@163.com*通信作者Corresponding author. E-mail: nmndwzj@163.com
劉文亭, 衛智軍, 呂世杰, 王天樂, 張爽, 海松. 放牧調控對短花針茅種群年齡及葉性狀的影響. 草業學報, 2017, 26(1): 63-71.
LIU Wen-Ting, WEI Zhi-Jun, LV Shi-Jie, WANG Tian-Le, ZHANG Shuang, HAI Song. Effect of grazing onStipabrevifloraage dynamics and leaf traits. Acta Prataculturae Sinica, 2017, 26(1): 63-71.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
