重引入狗獾釋放后定居巢的生境特征
2017-02-18 07:05:14吳彤褚可龍蔣文忠顧國林袁曉徐宏發
四川動物 2017年1期
吳彤, 褚可龍, 蔣文忠, 顧國林, 袁曉, 徐宏發*
(1. 華東師范大學生命科學學院,上海200241; 2. 上海市奉賢區林業署,上海201400; 3. 上海市野生動物保護管理站,上海200233)
重引入狗獾釋放后定居巢的生境特征
吳彤1, 褚可龍2, 蔣文忠2, 顧國林2, 袁曉3, 徐宏發1*
(1. 華東師范大學生命科學學院,上海200241; 2. 上海市奉賢區林業署,上海201400; 3. 上海市野生動物保護管理站,上海200233)
2014年4月—2016年3月,對上海郊區釋放的狗獾所建立的洞巢進行了調查。結果表明:異地引入的狗獾在大規模城市化的上海郊區仍能找到合適的棲息地,并能很好地適應野外的自然環境。生境特征分析顯示:釋放后的狗獾喜好在有地下空間、地勢較高的竹林、靠近水源、遠離公路的生境中營巢定居;在喬木密度高、草本長勢好、遠離公路的生境中覓食。狗獾食源地調查顯示:上海郊區狗獾冬季主要食物是蚯蚓和土壤昆蟲,其主要來源依次為:果林、落葉闊葉林、針葉林、竹林、常綠闊葉林。在上海郊區,重引入狗獾釋放成功須滿足5個條件:可利用的地下空間,地勢較高長勢好的竹林,成熟的果林、落葉闊葉林、針葉林提供食物,充足的水源以及避免人為干擾。
狗獾;重引入;洞巢;生境
狗獾Melesmeles是上海地區最大的陸生野生動物,20世紀80年代前上海郊區普遍有狗獾分布。近年來,快速的城市化發展致使狗獾適宜棲息地不斷減少和破碎化,目前僅奉賢區和松江區有零星分布,種群瀕臨區域性滅絕。為了挽救這一本土物種,維持上海地區生物多樣性和生態平衡,2007年上海市啟動了狗獾種群恢復項目。考慮到本地現存種群數量過少,已無法通過殘存種群擴大繁殖來復壯,因此只能通過異地引入狗獾來重建種群(IUCN/SSC,2013)。目前國外對狗獾的洞巢、生境、家域、食性已有大量研究(Roperetal.,2001;Macdonaldetal.,2004;Kanekoetal.,2006),包括對狗獾重引入也有詳細報道(Mulder,1996)。但國內對狗獾的研究多集中于洞巢結構、生境選擇、活動節律等方面(徐宏發等,1997;郝輝等,2009;殷寶法等,2011),有關重引入狗獾研究目前也只涉及行為和活動規律(徐循等,2012;崔勇勇等,2013),缺乏有關重引入狗獾的洞巢、生境適應等方面的詳細報道。為了確保狗獾重引入的成功,必須明確以下3個問題:一、選擇什么樣的區域和生境釋放最合適?二、影響釋放后狗獾成功定居的因素有哪些?三、定居后洞巢區的隱蔽性、食源以及其他生存條件能否滿足其生存需要?弄清這些問題對解決重引入所面臨的問題,以及今后在上海地區開展狗獾重引入工作具有重要意義。
1 研究地區概況
重引入狗獾釋放地位于上海市奉賢區金匯鎮申亞生態林(121°31′31″E,30°59′59″N)。該地地勢平坦,水系豐富,地下水位高(高程3~5 m)。年均氣溫16.6 ℃,夏季暖濕多雨,年均降水量1 328.3 mm,屬亞熱帶季風性濕潤氣候。1—2月氣溫最低(6.1~6.5 ℃),7—8月為高溫期(26.2~27.1 ℃)。該地原為農田,經植樹造林形成以闊葉喬木為主,配以果樹和竹林等有江南特色的本地植被。主要喬木有樟Cinnamomumcamphora、紫葉李Prunuscerasifera、雪松Cedrusdeodara、水杉Metasequoiaglyptostroboides等;果樹主要為梨Pyruspyrifolia、桃Amygdaluspersica;竹林以雷竹Phyllostachyspraecox為主;灌木包括八角金盤Fatsiajaponica、龜甲冬青Ilexcrenatavar.convexa等種類。冬季地表雜草主要為婆婆納Veronicadidyma、馬蘭Kalimerisindica、天胡荽Hydrocotylesibthorpioides等。該地原有狗獾棲息于地勢較高的竹林、河堤、墓穴等地,現已滅絕(上海市奉賢縣縣志修編委員會,1987;謝一民,2002)。
2 研究方法
2.1 洞巢分布及生境因子調查
2014年4月—2016年3月,對2007年秋季釋放的成年狗獾挖掘定居的洞巢進行調查,并采用夜間紅外探頭對發現的洞巢進行監測并用手持GPS定位。
狗獾洞巢一般由多個洞口組成,最外圍洞口圍成的區域稱為洞巢區,用格子法計算洞巢區面積,并測量洞口大小(長徑×短徑)(徐宏發等,1997;李偉等,2010),統計洞巢中總洞口數、利用洞口數(洞壁有抓爬踏蹬痕跡)、利用管道數(有狗獾棲息的地下廢棄管道)和建巢時間。統計狗獾洞巢在不同地形、植被、坡度和坡位中的洞口數量。
狗獾會隨著居住時間的增長和家族成員的增加不斷挖洞擴大巢穴(Roper,1992;徐宏發等,1997),以單位面積洞口數反映狗獾對營巢地的利用強度和喜好程度(徐宏發等,1997;殷寶法等,2011)。狗獾通過挖掘土壤獲取食物,留下大小(10 cm×8 cm~25 cm×16.5 cm)和深度(3~13.5 cm)不等的土坑,稱為食坑。利用單位面積食坑數可以衡量狗獾在生境中的取食強度。
生境因子調查采用樣方法。在洞巢區隨機設置10個10 m×10 m樣方,測量記錄樣方內的洞口數、食坑數、喬木密度、喬木基蓋度(地上1.3 m處喬木截面積占樣方面積百分比)、灌木密度、灌木蓋度、灌木基蓋度(地上3 cm處灌木基干截面積占樣方面積百分比)、竹密度和竹基蓋度(地上1.3 m處竹林截面積占單位面積百分比)、地下管道數。草本植物的調查采用在樣方中心和4角分別設置1 m×1 m小樣方,記錄小樣方的草本種類和蓋度;測量樣方中心距水源、公路和釋放點的距離(王正寰等,2003;Kanekoetal.,2006;李偉等,2010;楊會濤等,2010;李路云等,2015)。
2.2 冬季食源地分布調查
上海郊區的狗獾不冬眠(郝輝等,2009;徐循等,2012)。冬季是一年內食物最少的季節,為了解食物匱乏期的食物量和食源分布,判斷冬季食物能否滿足狗獾的需要,2015年12月—2016年2月,根據當地實際情況,選擇5種植被類型(果林、竹林、常綠闊葉林、落葉闊葉林、針葉林)進行狗獾食源調查。每隔30 d在各植被類型中隨機設置6個樣方(0.5 m×0.5 m),每個樣方深挖 20 cm,將土壤中所有狗獾可食用的部分收集起來,稱鮮重,然后在烘箱中50 ℃烘至恒重,稱干重(Goszczyńskietal.,2000;Kanekoetal.,2006;李峰等,2013)。
2.3 數據統計分析
利用單個樣本Kolmogorov-Smirnov Test檢驗生境因子和食源數據是否符合正態分布。經檢驗,生境因子、蚯蚓密度、總食源密度不符合正態分布(P<0.05),故采用Spearman相關分析檢驗樣方內洞口密度和食坑密度與生境因子間相關性、食坑密度與非正態分布食源數據間相關性。采用Mann-WhitneyUtest檢驗不同植被類型之間食坑密度、食源的差異,利用Pearson相關分析檢驗樣方內食坑密度與正態分布食源數據間相關性。數據均使用SPSS 22.0處理。
3 研究結果
3.1 洞巢分布及建立時間
結合實地調查和夜間紅外監測,確認申亞生態林及其周邊地區共有10個狗獾洞巢(表1)。10個洞巢大多在距原釋放點400 m范圍內,最近的洞巢在原釋放地(1、2號),最遠分布到林地的邊緣(5、7、10號)。
據每年的調查記錄,2008年僅發現零星深度不足1 m的臨時性洞口。2009年狗獾最先在距釋放點300 m的茂密竹林中建立洞巢(6號),2010年向北擴散建立洞巢(9號)。2010—2011年釋放地栽植的成片竹林逐漸繁茂,狗獾在此建立洞巢(1、2、3號),2012年向西、北擴散建立洞巢(5、10號)。2013年釋放地周邊新種植小片竹林,當年即有狗獾活動,次年建立洞巢(4號),2015年狗獾向東、西擴散建立了洞巢(7、8號)。
3.2 洞巢的特征
10個洞巢的特征測量統計結果見表1,最大的洞巢面積為2 580 m2,洞口83個(1號),最小的洞巢僅1個洞口(8號)(表1)。根據監測記錄,2013—2016年共4個洞巢(1、2、5、6號)發現有幼獾活動,可見釋放后狗獾已成功定居。調查并測量了10個洞巢的192個洞口,洞口最大為50 cm×44 cm,最小為19 cm×12 cm,平均大小(31.2±7.2) cm×(24.7±5.7) cm。
夜間監測發現,釋放后的狗獾首先棲息在廢棄的管道中,夜晚出管道在附近土坡挖掘洞口。絕大部分洞巢中的狗獾至少利用1個管道。
3.3 洞巢的生境類型
對10個洞巢、192個洞口的地形和植被進行了測量和統計,結果見表2。
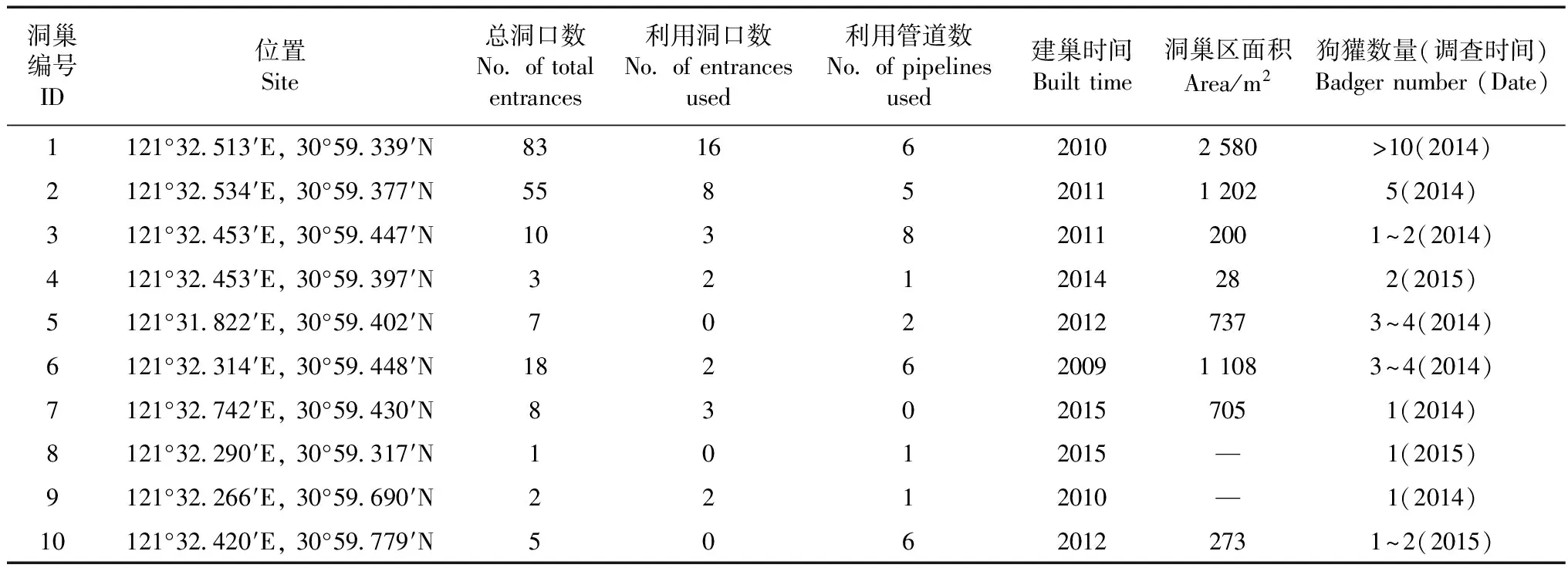
表1 狗獾洞巢的特征
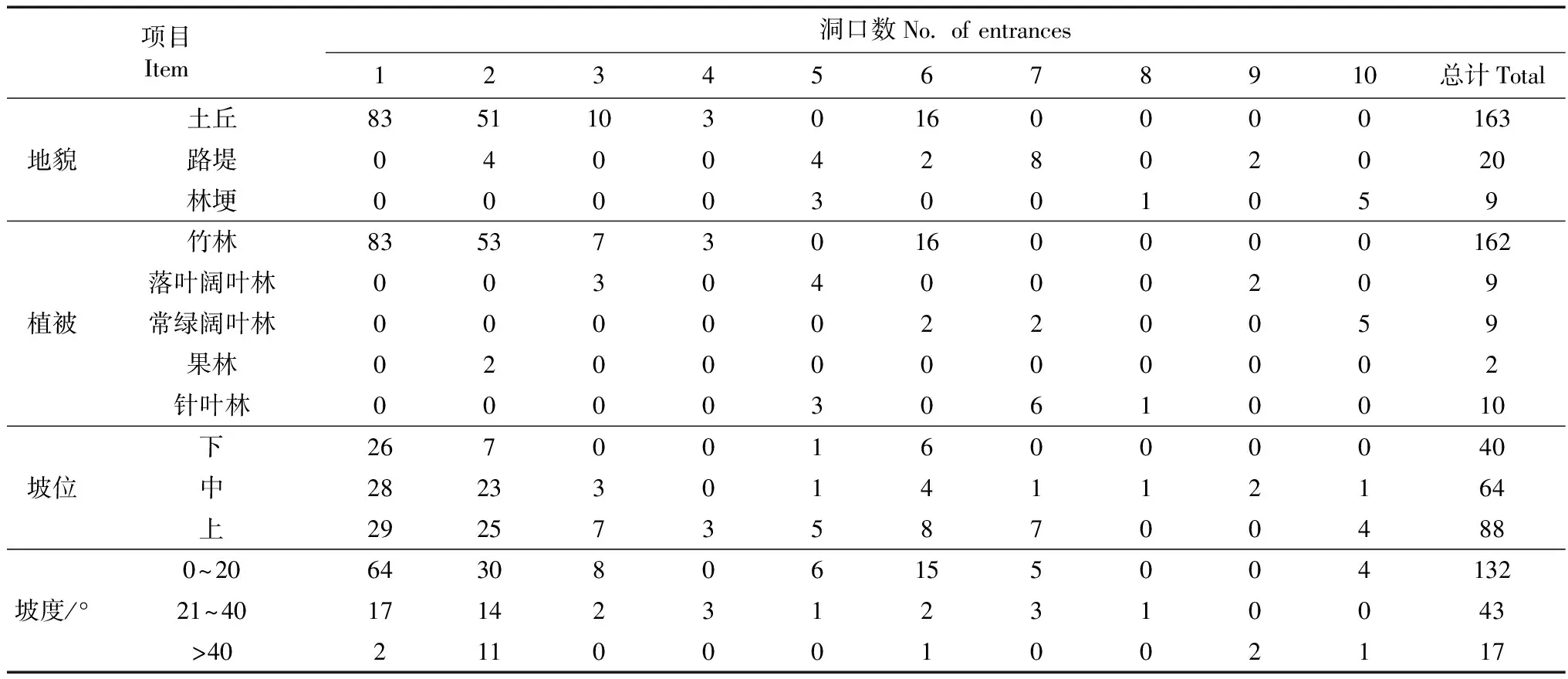
表2 狗獾洞巢的地形和植被
狗獾洞巢的洞口大多分布在竹林中(84.4%),果林最少(1.0%),洞口主要挖掘于土丘等地形稍高地區(84.8%),大部分洞口建立在中上坡位(79.2%),位于緩坡上的洞口數最多(68.8%)(表2)。
3.4 影響洞巢的生境因子
本研究調查了100個樣方,經單個樣本Kolmogorov-Smirnov Test檢驗,13個生境因子均不符合正態分布(P<0.05),且經數據轉換仍不符合正態分布,故采用Spearman相關分析檢驗洞口和食坑密度與
生境因子間的相關性,分析主要影響狗獾建巢和覓食活動的生境因子。
喬木密度、喬木基蓋度、距水源距離3個因子與洞口密度極顯著負相關(P<0.01),竹密度、竹林基蓋度、地下管道密度、距公路距離4個因子與洞口密度極顯著正相關(P<0.01)。據統計,55%以上洞口分布于喬木密度0%、喬木基蓋度0%、竹林密度>3.0棵/m2、竹林基蓋度>0.5%、地下管道密度>0.01個/m2的生境中,且所有洞口距離公路600 m以上,但均靠近水源(<30 m)(表3)。
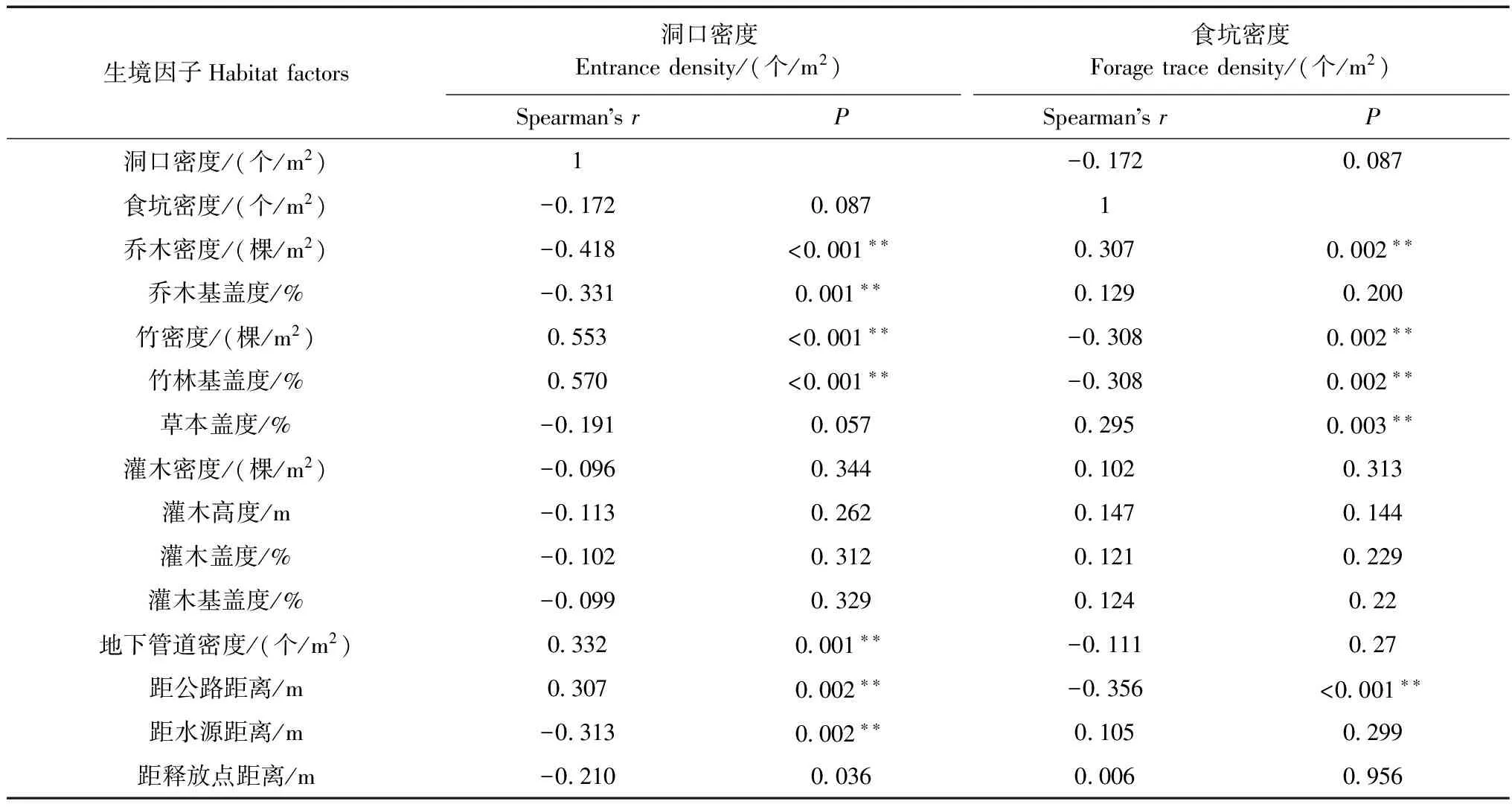
表3 狗獾洞口密度和食坑密度與生境因子的Spearman相關分析
注 Note:**P<0.01.
竹密度、竹林基蓋度、距公路距離3個因子與食坑密度極顯著負相關(P<0.01),喬木密度、草本蓋度與食坑密度極顯著正相關(P<0.01)。據統計,65%以上食坑分布于喬木密度>0.05棵/m2、草本蓋度>30%、竹林密度<3.0棵/m2、竹林基蓋度<0.5%的生境中,且所有食坑距離公路200 m以上(表3)。
3.5 狗獾冬季的食源
5種植被生境的30個樣方調查結果顯示,冬季土壤中狗獾主要食源為昆蟲成蟲(暗黑鰓金龜Holotrichiaparallela)、昆蟲幼蟲(蠐螬Melolonthidaeg.)和蚯蚓(環毛蚓Pheretimaspp.),其密度、鮮重及干重見表4。
土壤中狗獾的食物主要是蚯蚓,其密度和質量均最高,昆蟲成蟲和幼蟲次之。在密度及質量上,果林中食源各指標顯著高于常綠闊葉林及竹林(P<0.05),而其他生境類型之間食源各指標差異無統計學意義。
各植被類型中的食坑數據和食源數據取平均值,采用Kolmogorov-Smirnov Test檢驗各數據正態性,除蚯蚓密度、總食源密度不符合正態分布,采用Spearman相關分析檢驗外,其余數據均符合正態分布,利用Pearson 相關分析檢驗食坑密度與食源間的相關性。
從表5可見,蚯蚓的干、鮮重及總食源的干、鮮重與狗獾食坑密度顯著正相關。果林(0.061個/m2)、落葉闊葉林(0.053個/m2)和針葉林(0.045個/m2)食坑密度顯著高于常綠闊葉林(0.024個/m2)和竹林(0.013個/m2)(P<0.05),常綠闊葉林食坑密度顯著高于竹林(P<0.05)。申亞生態林區種植有大片果林,因此可以認為該地區冬季能滿足狗獾的食物需要。
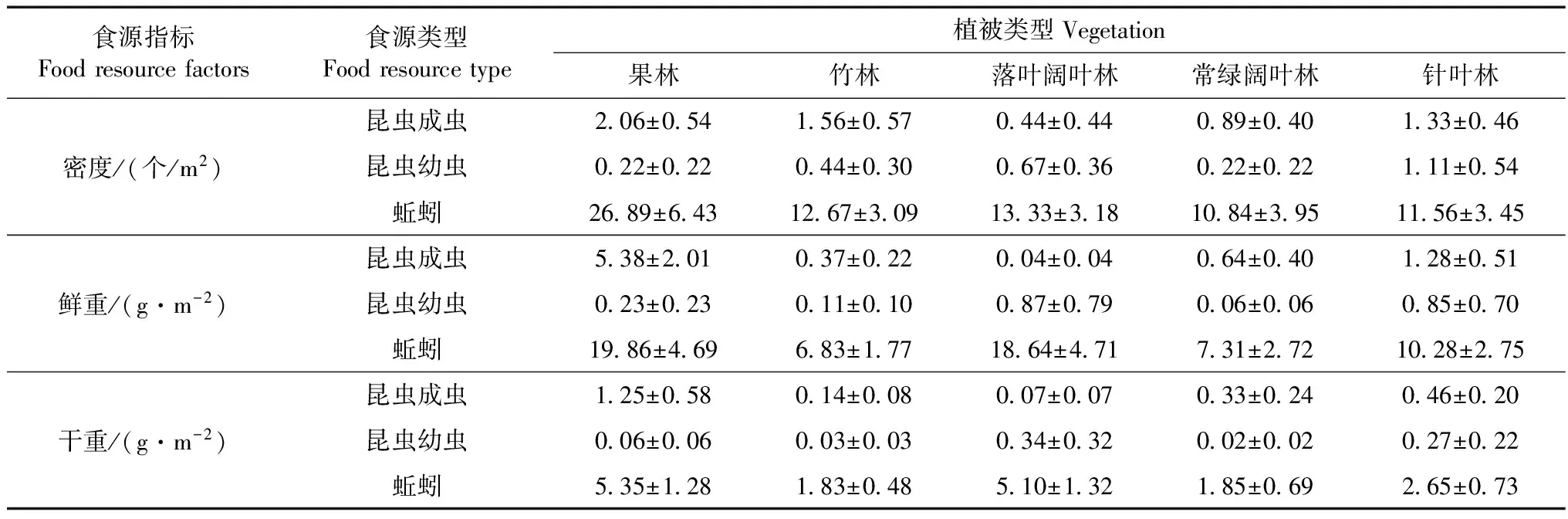
表4 狗獾冬季棲息地食源的密度、鮮重及干重

表5 狗獾食坑密度和食源的相關分析
注 Note:*P<0.05.
4 討論
狗獾在上海郊區被釋放后最初均利用地下空間作為隱蔽的居所,然后在此基礎上建巢,在荷蘭的狗獾重引入中也有相同的情況(Mulder,1996)。雖然狗獾善于挖洞(徐宏發等,1997;Roperetal.,2001;Wilsonetal.,2003),但建造一個復雜的洞穴需耗費大量的能量和時間(徐宏發等,1997;葉曉堤等,1999),利用原有地下空間加以改造是最合適的選擇。釋放地廢棄的排水管道建在田埂上,地勢較高,沒有地下水侵襲,成為剛釋放狗獾良好的藏身地。隨后狗獾利用原有管道不斷擴展,形成復雜的地下洞穴。可見狗獾重引入必須考慮當地有沒有可利用的地下空間,如沒有,應人工建造不受雨水和地下水影響的地下空間供狗獾使用。
釋放地周圍竹林面積僅占總面積的0.4%,但84.4%的狗獾洞口卻建在竹林中,表明狗獾偏好在竹林營巢,這和本地原有的野生狗獾習性類似(謝一民,2002;李偉等,2010)。釋放地的竹林一般栽植于地勢稍高的土丘,狗獾在土丘的斜坡上挖洞,有利于將洞內的新土推出(徐宏發等,1997;Macdonaldetal.,2004;李路云等,2015),高地勢可防止雨水和地下水侵入狗獾掘于中上坡位的洞口(李路云等,2015)。此外,地下發育良好的竹鞭和竹根有良好的固土作用(李偉等,2010),保證狗獾挖掘的洞穴不會崩塌;冬、春季竹林可為狗獾提供嫩筍作為食源;密度高的竹林有良好的隱蔽性。
釋放后狗獾全年主要的食物有昆蟲、蚯蚓、瓜果(梨、白果、瓜類等)、竹筍和小型脊椎動物(蛙類、鳥類等)。本研究表明,果林、落葉闊葉林和針葉林可為狗獾提供豐富的食物。由于當地的果林和落葉闊葉林定期翻耕土壤、清除雜草,該生境中土壤松軟,保水性較好,蚯蚓的數量多;另外,在夏季,果林中未成熟的果實會大量脫落,成熟期也有不少果實落地,這些都為狗獾提供了豐富的食物。針葉林中,地表多年的落葉使土質松軟潮濕,蚯蚓和昆蟲也十分豐富。因此,盡管狗獾很少在果林、落葉闊葉林、針葉林中建巢,但卻是它良好的取食地。
本研究地水系豐富,狗獾洞口均靠近水源地(<30 m),且發現水邊有大量狗獾足跡。從夜間紅外監測儀的監測來看,狗獾經常到水邊活動,除飲水、洗澡外,還捕食蛙類等脊椎動物。東北狗獾M.m.amurensis為了獲取充足的水源通常選擇在河流附近營巢(李路云等,2015),可見水源的可利用性對狗獾的生存非常重要。
上海郊區交通十分發達,在釋放區有一條繁忙的公路穿過生態林,人為干擾較大。調查發現,重引入狗獾挖掘的洞口均距離公路600 m以上,且都選擇竹林土丘、路堤、林埂等隱蔽性高的地方。狗獾洞巢一般有多個洞口,但大多數洞口很少利用,一旦狗獾發現洞口被干擾則會放棄轉而使用其他洞口或挖掘新洞(徐宏發等,1997;Roperetal.,2001)。相關研究表明人為干擾是動物棲息地選擇的負面因素(Forman & Alerander,1998;胡忠軍等,2005),因而在狗獾的重引入釋放過程中,除釋放地應隱蔽、遠離公路等人為活動極頻繁區域以外,還應采取適當的措施防止當地居民的踩踏、獵捕等干擾。
5 結論
重引入狗獾釋放后在大規模城市化的上海郊區仍能找到合適的棲息地,并能成功定居,其成功的必要條件是:一、可利用的地下空間;二、地勢較高的竹林生境;三、成熟的果林、落葉闊葉林、針葉林提供食物;四、充足的水源;五、避免人為干擾。
致謝:華東師范大學2013級薛敬冬、束瀟瀟同學參加了部分野外調查工作,上海市申亞測報點李國勝協助參與了調查,在此一并感謝。
崔勇勇, 謝志剛, 徐循, 等. 2013. 引入狗獾活動規律和取食行為的研究[J]. 安徽農業科學, 41(8): 3407-3409.
郝輝, 褚可龍, 裴恩樂, 等. 2009. 上海郊區狗獾活動規律的初步研究[J]. 四川動物, 29(1): 111-114.
胡忠軍, 于長青, 徐宏發, 等. 2005. 道路對陸棲野生動物的生態學影響[J]. 生態學雜志, 24(4): 433-437.
李峰, 羅振華, 李春林, 等. 2013. 古北界狗獾食性的生物地理模式:狗獾是蚯蚓專食者嗎?[J]. 科學通報, 58(16): 1565-1572.
李路云, 楊會濤, 滕麗微, 等. 2015. 張廣才嶺藏獾洞穴生境選擇[J]. 生態學報, 35(14): 4836-4842.
李偉, 謝志剛, 褚可龍, 等. 2010. 上海郊區狗獾洞穴特征及分布研究[J]. 安徽農業科學, 38(22): 11838-11840.
上海市奉賢縣縣志修編委員會. 1987. 奉賢縣志[M]. 上海: 上海人民出版社: 15-40.
王正寰, 王小明, 吳巍, 等. 2003. 四川西部石渠地區夏季藏狐巢穴選擇的生境分析[J]. 獸類學報, 23(1): 31-38.
謝一民. 2002. 上海陸生野生動植物資源(三)[M]. 上海: 上海市農林局: 209-210.
徐宏發, 陸厚基, 王小明. 1997. 鹽城保護區狗獾洞巢的初步研究[J]. 獸類學報, 17(2): 107-112.
徐循, 謝志剛, 崔勇勇, 等. 2012. 重引入狗獾秋冬季行為的初步研究[J]. 動物學雜志, 47(3): 49-52.
楊會濤, 劉振生, 徐坤, 等. 2010. 狗獾秋季對生境的選擇——以黑龍江省方正林業局為例[J]. 生態學報, 30(7): 1875-1881.
葉曉堤, 馬勇, 王潤海, 等. 1999. 沂泗水流域狗獾的洞道結構及其功能[J]. 獸類學報, 19(3): 231-232.
殷寶法, 劉宇慶, 劉國興, 等. 2011. 京杭運河堤壩區域狗獾的棲息地選擇[J]. 生態學報, 31(7): 2002-2007.
Forman TT, Alexander LE. 1998. Roads and their major ecological effects[J]. Annual Review of Ecology Evolution & Systematics, 29(4): 207-231.
Goszczyńsk J, Jedrzejewska B, Jedrzejewski W. 2000. Diet composition of badgers (Melesmeles) in a pristine forest and rural habitats of Poland compared to other European populations[J]. Journal of Zoology, 250(4): 495-505.
IUCN/SSC. 2013. Guidelines for reintroductions and other conservation translocations. Version 1.0[S]. Gland, Switzerland: IUCN Species Survival Commission, IX+57pp.
Kaneko Y, Maruyama N, Macdonald DW. 2006. Food habits and habitat selection of suburban badgers (Melesmeles) in Japan[J]. Journal of Zoology, 270(1): 78-89.
Macdonald DW, Newman C, Dean J,etal. 2004. The distribution of Eurasian badger,Melesmeles, setts in a high density area: field observation contradict the sett dispersion hypothesis[J]. Oikos, 106(2): 295-307.
Mulder JL. 1996. Reintroducing the badgerMelesmeles: stories of failure and success[J]. Lutra, 39: 1-32.
Roper TJ. 1992. BadgerMelesmelessetts-architecture, internal environment and function[J]. Mammal Review, 22(1): 43-53.
Roper TJ,Ostler JR, Selllnld TK,etal. 2001. Sett use in European badgersMelesmeles[J]. Behaviour, 138(2): 173-187.
Wilson GJ,Delahay RJ, Leeuw ANS,etal. 2003. Quantification of badger (Melesmeles) sett activity as a method of predicting badger numbers[J]. Journal of Zoology, 259(1): 49-56.
Setts Habitat Features of ReintroducedMelesmelesafter Release
WU Tong1, CHU Kelong2, JIANG Wenzhong2, GU Guolin2, YUAN Xiao3, XU Hongfa1*
(1. College of Life Sciences, East China Normal University, Shanghai 200241, China; 2. Forestry Bureau of Fengxian District, Shanghai 201400, China; 3. Shanghai Wildlife Conservation Bureau, Shanghai 200233, China)
In this study, the setts built by badgers (Melesmeles) were investigated in Shanghai suburbs from April 2014 to March 2016. The results showed that the reintroduced badgers were able to find appropriate habitats and adapt natural environment of Shanghai suburbs with large-scale urbanization. Based on the analysis of habitat characteristics, the reintroduced badgers preferred the habitats with underground space and high-terrain bamboo forests that were close to water sources and far away from highways. Moreover, the reintroduced badgers foraged food in the habitats with high-dense trees, fine herbage coverage and long distance from highways. The result of food resource survey indicated that badgers were mainly fed on earthworms and insects in winter. Their food resources were from fruit forests, deciduous broad-leaf forests, coniferous forests, bamboo forests and evergreen broad-leaf forests. Overall, these findings suggested that the establishment of wild colony of badgers in Shanghai suburbs requires 5 elements including underground space, high-terrain bamboo forests, sufficient water and minimum human disturbance, as well as food supplies from mature fruit forests, broad-leaved forests and coniferous forests.
Melesmeles; reintroduced; sett; habitat
2016-07-08 接受日期:2016-11-16
國家林業局上海市極小種群(狗獾)恢復項目; 上海市農業委員會項目[滬農科攻字(2014)第5-5號]
吳彤(1991—), 男, 碩士, 研究方向為動物生態學, E-mail:luguiwutong@foxmail.com
*通信作者Corresponding author, E-mail:hfxu@bio.ecnu.edu.cn
10.11984/j.issn.1000-7083.201600184
Q958.1; Q959.8
A
1000-7083(2017)01-0094-06
